
Thioredoxin Domain Containing 5 (TXNDC5): Friend or Foe?

 , ,
, ,  and
and 
Abstract
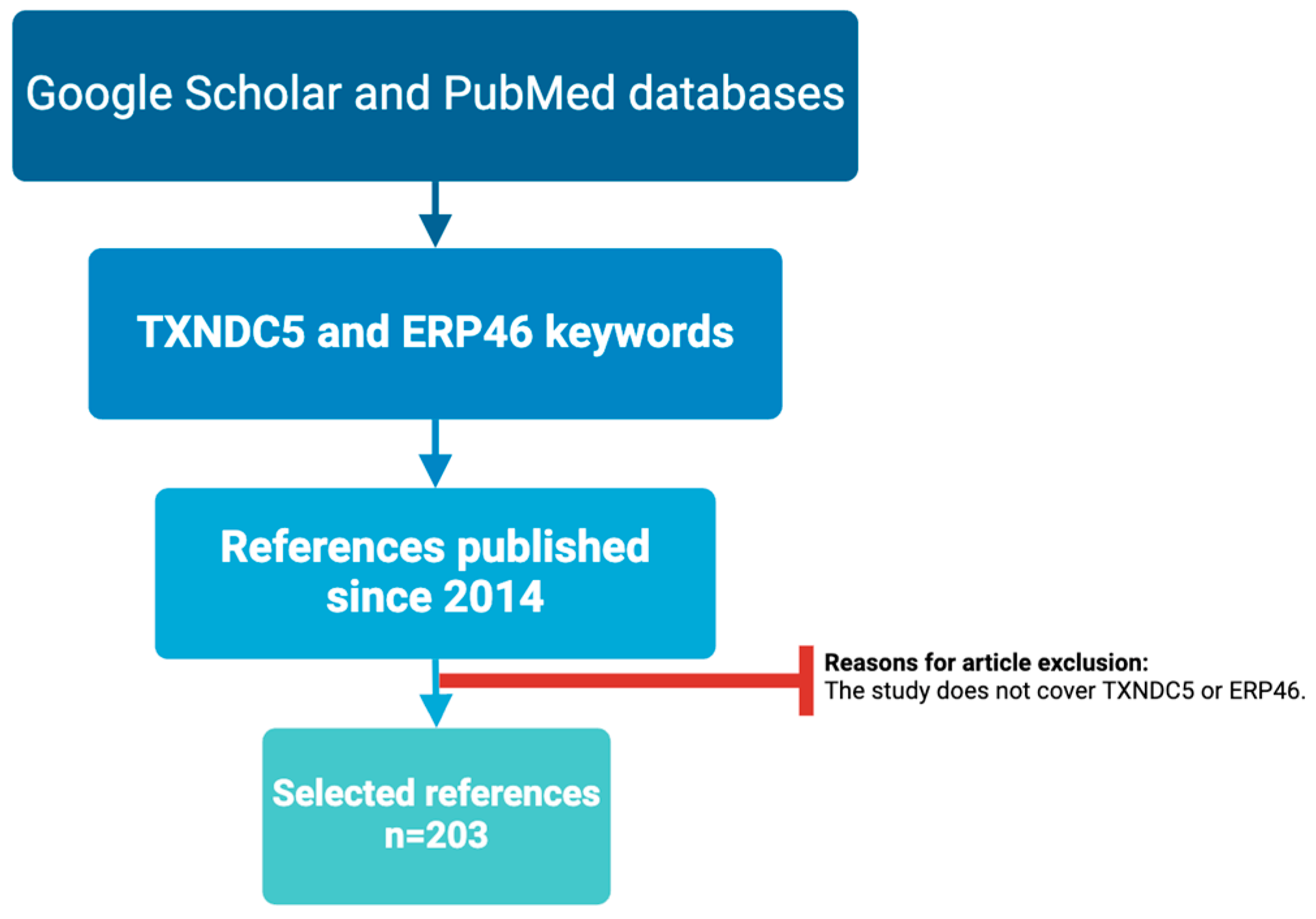
:1. Introduction
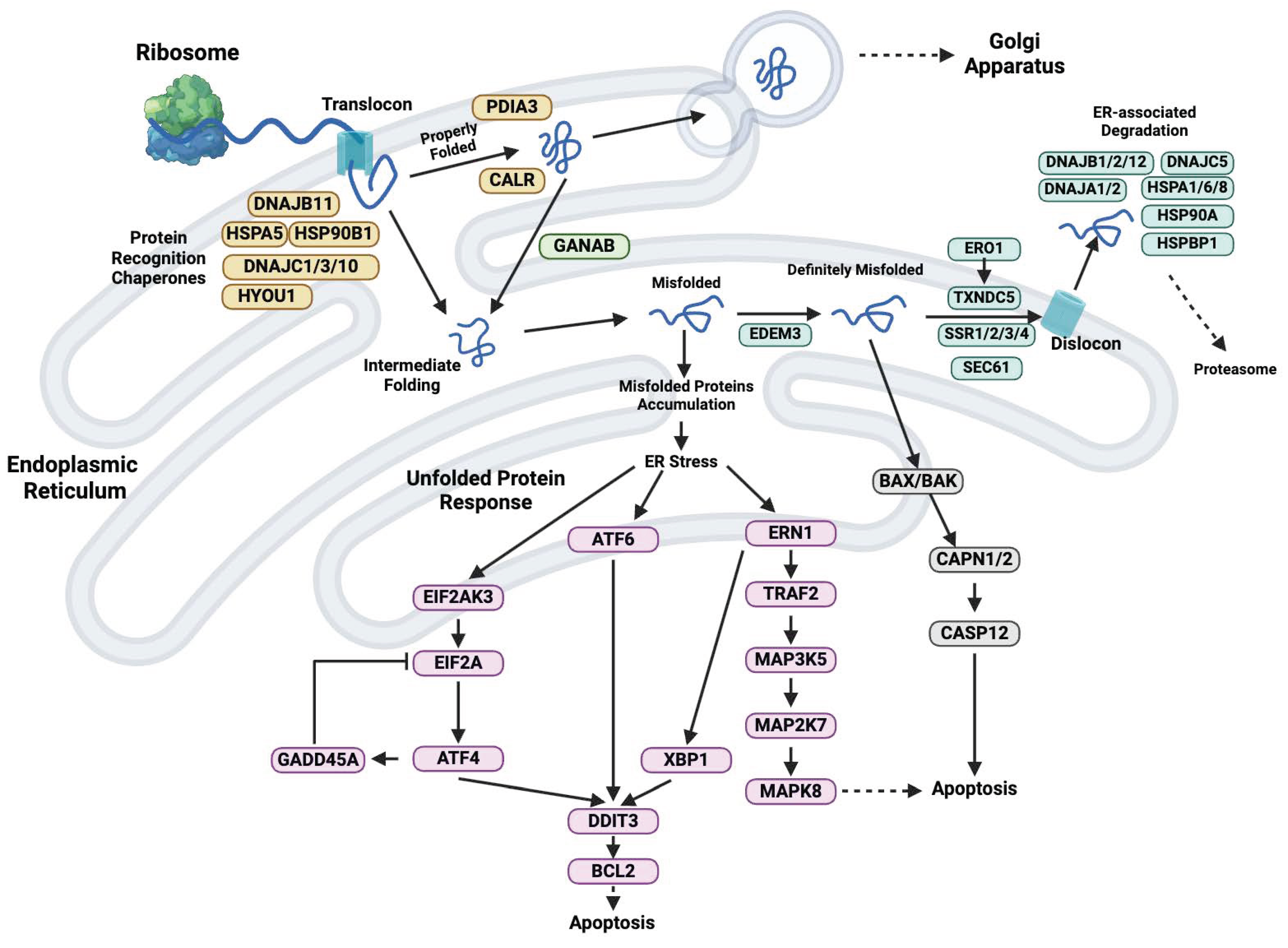
2. TXNDC5 Gene and Protein Structure
3. LncRNAs, Circular RNAs and MicroRNAs Targeting TXNDC5
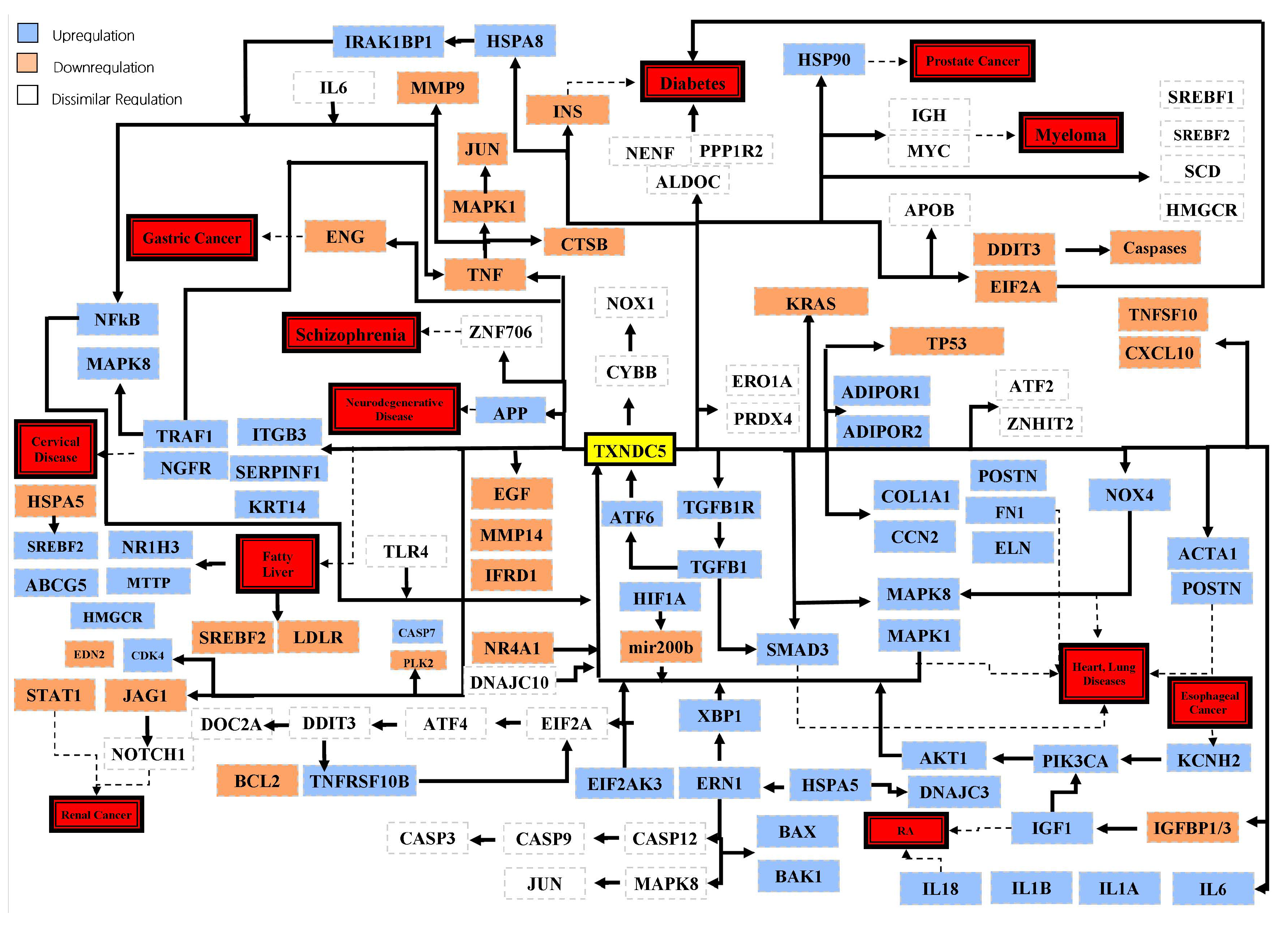
4. TXNDC5 and Cancers
4.1. TXNDC5 and Prostate Cancer
4.2. TXNDC5 and Breast Cancer
4.3. TXNDC5 and Endometrial and Ovarian Cancer
4.4. TXNDC5 and Skin Cancer
4.5. TXNDC5 and Brain Cancer
4.6. TXNDC5 and Colorectal Cancer
4.7. TXNDC5 and Cervical Cancer
4.8. TXNDC5 and Lung Cancer
4.9. TXNDC5 and Soft Tissue Cancer
4.10. TXNDC5 and Kidney Cancer
4.11. TXNDC5 and Liver Cancer
4.12. TXNDC5 and Blood Cancer
4.13. TXNDC5 and Other Types of Cancer

{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Gene Symbols * | Related Pathways | References |
|---|---|---|---|
| Prostate | HIF1A 1, NAAA 2, SH3BP4 3, ID1 4, RAB1B 5, HMGCR 6, HNRNPK 7, NDRG1 8, IDI1 9, VIM 10 | Proteasome-mediated degradation 1 Neurotransmitter release cycle 2 Cellular responses to stimuli 3,4 Cell cycle 5 Metabolism of steroids 6,9 Processing of capped intron-containing pre-mRNA 7 Gene expression 8 Cholesterol biosynthesis 9 Selective autophagy 10 | [42,49,50] |
| Breast | ALB 1, ANGPTL7 2, IL6 3, NGB 4, BLOC1S5 5, NR4A1 6, ITGB1 7, CDKN1A 8, SERPINB5 9, GADD45α 10 | Response to elevated platelet cytosolic Ca2+ 1 Apoptotic pathways 2,3,7,9 MIF-mediated glucocorticoid regulation 3 Intracellular oxygen transport 4 Biogenesis of lysosome-related organelles 5 Gene expression 6,10 Proteasome-mediated degradation 8 Selective autophagy 9 | [51,52] |
| Endometrial | NR4A1 1, IDH1 2, SESN2 3, PRKAA1 4 | Gene expression 1,3,4 Innate immune system 2 Cellular responses to stimuli 3 | [53,55] |
| Cutaneous squamous cell | APOA1 1, ALB 2, SERPINA1 3, HLA-B 4, HP 5, METTL3 6, STAT4 7, TNFRSF17 8, TNFAIP3 9, GSTM1 10, GSTM3 11 | Plasma lipoprotein assembly, remodeling, and clearance 1 Response to elevated platelet cytosolic Ca2+ 1,2,3 Innate immune system 4,5,7 Processing of capped intron-containing pre-mRNA 6 MIF-mediated glucocorticoid regulation 8 Metabolism of proteins 9 Glutathione conjugation 10,11 | [58,59,60] |
| Brain | P4HB 1, PDIA3 2, PDIA4 3, PDIA5 4, PDIA6 5, ERP27 6, ERP29 7, ERP44 8, TMX1 9, TMX3 10, TXNDC12 11, AGR3 12, DNAJC10 13, NR4A1 14, PRDX4 15 | Plasma lipoprotein assembly, remodeling, and clearance 1 Antigen processing–cross presentation 2 Calnexin/calreticulin cycle 2 Translational control 3,8 Unfolded protein response 4,5,6 Cellular responses to stimuli 4,5 Protein disulfide isomerase activity 7,9,11,13 Response to elevated platelet cytosolic Ca2+ 10,12 Gene expression 14 Innate immune system 15 | [41,61,62,66] |
| Colorectal | HIF1A 1, DDIT3 2, HSPA5 3, ATF4 4, BAX 5, CASP8 6, IDH1 7, NR4A1 8, PRDX4 9, EDEM3 10 | Proteasome-mediated degradation 1 Gene expression 2 Unfolded protein response 2,3 Response to elevated platelet cytosolic Ca2+ 3 Integrated stress response 4 MIF-mediated glucocorticoid regulation 5 Apoptotic pathways 6 Innate immune system 7,9 Gene expression 8 Calnexin/calreticulin cycle 10 Metabolism of proteins 10 | [32,67,68,69,70,71,72,73] |
| Cervical | SERPINF1 1, TRAF1 2, METTL3 3, EDEM3 4, PRDX4 5, GPX7 6, ERO1α 7, ITPR1 8, DNAJC10 9, LIF 10, NDUFA5 11, RLIM 12, SERF1A 13 | Apoptotic pathways 1,2 Processing of capped intron-containing pre-mRNA 3 Chromatin regulation 3 Calnexin/calreticulin cycle 4 Metabolism of proteins 4 Innate immune system 5,12 Cellular responses to stimuli 6,7 Development angiotensin activation 8 Protein disulfide isomerase activity 9 MIF-mediated glucocorticoid regulation 10 Respiratory electron transport 11 Protein aggregation 13 | [73,74,76,77,78,79,80,81,82,83,84] |
| Non-small cell lung | ERN1 1, XBP1 2, HSPA5 3, SRXN1 4, PRDX6 5, PDIA6 6, TGFβ1 7, TGFBR1 8, ATF6 9 | Cellular responses to stimuli 1,2,4,5,6,9 Unfolded protein response 2,3,6,9 Response to elevated platelet cytosolic Ca2+ 3 Innate immune system 5 Apoptotic pathways 7,8 | [86,88,89,90,91,93] |
| Rhabdomyosarcoma | NR4A1 1, IDH1 2, IL24 3, TP53 4, EIF2AK3 5, HSPA5 6, ATF4 7, DDIT3 8, E2F1 9, CCND2 10, BIRC5 11, CDK4 12, STAT5A 13, EGFR 14, SP1 15, EP300 16, POLR2A 17, ERO1 18, HCP5 19, TXNDC12 20, DNAJC10 21, AGO2 22 | Gene expression 1,8,11 Innate immune system 2,19 MIF-mediated glucocorticoid regulation 3 Proteasome-mediated degradation 4 Cellular responses to stimuli 5,18 Unfolded protein response 6,8 Response to elevated platelet cytosolic Ca2+ 6,20 Integrated stress response 7 Apoptotic pathways 9,14 Protein kinase binding 10 Cellular response to stress 12 Prolactin signaling 13,14,15 RNA polymerase and promoter opening 16,17 Protein disulfide isomerase activity 20,21 Transcriptional regulation 22 Cell junction organization 22 | [41,44,95,96,97,98] |
| Renal | NR4A1 1, IDH1 2, E2F1 3, CCND2 4, BIRC5 5, CDK4 6, STAT5A 7, SP1 8, AGO2 9, ATF6 10, TGFβ1 11, TGFBR1 12, EDEM3 13, ITPR1 14, SMAD3 15, IL12A 16, IL23α 17 | Gene expression 1,5 Innate immune system 2 Apoptotic pathways 3,11,12 Protein kinase binding 4 Cellular response to stress 6 Prolactin signaling 7,8 Transcriptional regulation 9 Cell junction organization 9 Cellular responses to stimuli 10 Unfolded protein response 10 Calnexin/calreticulin cycle 13 Metabolism of proteins 13 Development angiotensin activation 14 Proteasome-mediated degradation 15 Mediated glucocorticoid regulation 16,17 | [41,43,85,99,100,103,104,105,108,109,110,111,112,114] |
| Liver | SAA1 1, SAA2 2, APOA1 3, APOB 4, PRDX4 5, PRDX6 6, HSPA9 7, HSPA5 8, GSTM3 9, CRELD2 10, STAT3 11, TGFβ1 12, ATF6 13, ERN1 14, EIF2AK3 15, CALR 16, APMAP 17, SERPINA1 18, SREK1 19 | Cholesterol homeostasis 1,2 Plasma lipoprotein assembly, remodeling, and clearance 3,4 Response to elevated platelet cytosolic Ca2+ 3,8,10,18 Innate immune system 4,5 Cellular responses to stimuli 5,7,13,14,15 Unfolded protein response 8,10,13,16 Glutathione conjugation 9 Prolactin signaling 11 Apoptotic pathways 12 Adipocyte differentiation 17 Transport to the Golgi and subsequent modification 18 Processing of capped intron-containing pre-mRNA 19 | [38,105,118,119,120,121,122,123,124,125,126,127,129] |
| Myeloma | BCL2L11 1, CCND1 2, CCND2 3, LTBR 4, PDIA3P1 5, HSP90B1 6, MZB1 7, ACTB 8, P4HB 9, DERL3 10, HERPUD1 11, PDIA4 12, PDIA6 13, RRBP1 14, SSR3 15, SSR4 16, UBE2J1 17, TNFSF13 18, MYC 19, NSD2 20, MAF 21, FCHSD2 22, NSMCE2 23, MAP3K14 24, GALM 25, BMP6 26, FOXO3 27, DDIT3 28, ATF3 29, SEC24D 30, DHX15 31 | Apoptotic pathways 1,4,7,26 Proteasome-mediated degradation 2,10 Protein kinase binding 3,27 Protein disulfide isomerase activity 5 Unfolded protein response 6,13,28 Cellular responses to stimuli 6,11,13,29 Regulation of actin dynamics for phagocytic cup formation 8 Plasma lipoprotein assembly, remodeling, and clearance 9 PERK regulates gene expression 11,29 Translational control 12 Signaling receptor activity 14 Metabolism of proteins 15,16,23,24 Regulation of degradation 17 MIF-mediated glucocorticoid regulation 18,26 Prolactin signaling 19 Homology directed repair 20 DNA double strand break response 20 Gene expression 21,28 Endocytosis 22 Galactose metabolism 25 Vesicle trafficking 30 Processing of capped intron-containing pre-mRNA 31 | [137,139,140,141,142,143,144,145,146,147,148,149,150,151] |
| Esophageal squamous cell | KCNH2 | Potassium channels | [155] |
| Endothelial cell | YAP1 | Gene expression | [158] |
| Pancreas | NR4A1 1, CTNNB1 2 | Gene expression 1,2 Phosphorylation 2 | [71,159,160,161,162] |
| Head and neck | DDIT3 1, CASP3 2 | Gene expression 1 Unfolded protein response 1 Apoptotic pathways 2 | [163] |
5. TXNDC5 and Fibrosis-Related Pathologies
6. TXNDC5 and Rheumatoid Arthritis
7. TXNDC5 and Diabetes Mellitus
8. TXNDC5 and Heart Diseases
| Diseases | Gene Symbols * | Related Pathways | References |
|---|---|---|---|
| Rheumatoid arthritis | DNAJB11 1, CALR 2, ERP29 3, GANAB 4, HSP90B1 5, HSPA1A 6, HSPA5 7, HYOU1 8, LMAN1 9, PDIA4 10, CXCL8 11, IL6 12, TNF-α 13, HSPA8 14, PCSK6 15, HIF1A 16, CXCL10 17, TNFSF10 18, IGFBP1 19 | Unfolded protein response 1,2,5,7,8 Cellular responses to stimuli 1,5,8,19 Processing of secretory proteins 3 Translation of structural proteins 4 Proteasome-mediated degradation 6,16 Response to elevated platelet cytosolic Ca2+ 7 Signaling by Rho GTPases 9 Translational control 10 Apoptotic pathways 11,12 MIF-mediated glucocorticoid regulation 11,12,13,17,18 Processing of capped intron-containing pre-mRNA 14 Selective autophagy 14 Plasma lipoprotein assembly, remodeling, and clearance 15 Gene expression 19 | [40,166,167,168,173,174,175,176] |
| Diabetes mellitus | GLP-1 1, ATF6 2, XBP1 3, ERN1 4, IGF1 5 | Integration of energy metabolism 1 Unfolded protein response 2,3 Cellular responses to stimuli 2,3,4 Apoptotic pathways 5 | [168,177,180] |
| Heart diseases | ATF6 1, TNF-α 2, NOX4 3, SMAD3 4, TGFβ1 5, HSF1 6, HSP90AB1 7, NOS3 8, KLF2 9, ITGA2B 10, PDIA6 11, HSPA5 12, CALR 13 | Unfolded protein response 1,11,12,13 Cellular responses to stimuli 1,3,11 MIF-mediated glucocorticoid regulation2 Proteasome-mediated degradation 4 Apoptotic pathways 5,10 Cellular response to heat stress 6 Selective autophagy 6 Inflammasomes 7 Metabolism of nitric oxide 8 Cell differentiation 9 Response to elevated platelet cytosolic Ca2+ 12 | [43,180,181,182,183,184,185,188,189,191] |
9. TXNDC5 and Other Disorders
10. Conclusions

Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hung, C.-T.; Tsai, Y.-W.; Wu, Y.-S.; Yeh, C.-F.; Yang, K.-C. The novel role of ER protein TXNDC5 in the pathogenesis of organ fibrosis: Mechanistic insights and therapeutic implications. J. Biomed. Sci. 2022, 29, 63. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Chang, X. The role and mechanism of TXNDC5 in diseases. Eur. J. Med. Res. 2022, 27, 145. [Google Scholar] [CrossRef] [PubMed]
- Okumura, M.; Kadokura, H.; Inaba, K. Structures and functions of protein disulfide isomerase family members involved in proteostasis in the endoplasmic reticulum. Free Radic. Biol. Med. 2015, 83, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Janssen-Heininger, Y.; Reynaert, N.L.; van der Vliet, A.; Anathy, V. Endoplasmic reticulum stress and glutathione therapeutics in chronic lung diseases. Redox Biol. 2020, 33, 101516. [Google Scholar] [CrossRef] [PubMed]
- Kuribara, T.; Totani, K. Structural insights into N-linked glycan-mediated protein folding from chemical and biological perspectives. Curr. Opin. Struct. Biol. 2021, 68, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, J.; Liu, L.; Li, J. The crucial role of demannosylating asparagine-linked glycans in ERADicating misfolded glycoproteins in the endoplasmic reticulum. Front. Plant Sci. 2021, 11, 625033. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, O.A.; Bartosch, B.; Zakirova, N.F.; Kochetkov, S.N.; Ivanov, A.V. Polyamine metabolism and oxidative protein folding in the ER as ROS-producing systems neglected in virology. Int. J. Mol. Sci. 2018, 19, 1219. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, C.; Machida, K.; Noi, K.; Murakawa, T.; Okumura, M.; Ogura, T.; Imataka, H.; Inaba, K. Distinct roles and actions of protein disulfide isomerase family enzymes in catalysis of nascent-chain disulfide bond formation. iScience 2021, 24, 102296. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: From mechanisms to medicines. Nature 2020, 587, 555–566. [Google Scholar] [CrossRef]
- Libby, P. The changing landscape of atherosclerosis. Nature 2021, 592, 524–533. [Google Scholar] [CrossRef]
- Chen, D.-L.; Xiang, J.-N.; Yang, L.-Y. Role of ERp46 in β-cell lipoapoptosis through endoplasmic reticulum stress pathway as well as the protective effect of exendin-4. Biochem. Biophys. Res. Commun. 2012, 426, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Torres, A.; Barceló-Batllori, S.; Martínez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Arnal, C.; Guillén, N.; Acín, S.; Osada, J. Proteomics and gene expression analyses of squalene-supplemented mice identify microsomal thioredoxin domain-containing protein 5 changes associated with hepatic steatosis. J. Proteom. 2012, 77, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Costas, A.O. Characterization of the Element of Response to Squalene in the Murine Gene Cyp2b10. Master’s Thesis, University of Zaragoza, Zaragoza, Spain, 2021. [Google Scholar]
- Wang, L.; Zheng, Y.; Xu, H.; Yan, X.; Chang, X. Investigate pathogenic mechanism of TXNDC5 in rheumatoid arthritis. PLoS ONE 2013, 8, e53301. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Xu, B.; Wang, L.; Wang, Y.; Wang, Y.; Yan, S. Investigating a pathogenic role for TXNDC5 in tumors. Int. J. Oncol. 2013, 43, 1871–1884. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Paskeh, M.D.A.; Entezari, M.; Bidooki, S.H.; Ghaleh, V.J.; Rezaei, S.; Hejazi, E.S.; Kakavand, A.; Behroozaghdam, M.; Movafagh, A.; et al. siRNA and targeted delivery systems in breast cancer therapy. Clin. Transl. Oncol. 2023, 25, 1167–1188. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Liu, C.M.; Liu, Y.L.; Shen-Jang Fann, C.; Hsiao, P.C.; Wu, J.Y.; Hung, S.I.; Chen, C.H.; Wu, H.M.; Jou, Y.S.; et al. Clustering by neurocognition for fine mapping of the schizophrenia susceptibility loci on chromosome 6p. Genes Brain Behav. 2009, 8, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.H.; Shin, M.K.; Uhm, Y.K.; Kim, H.J.; Chung, J.H.; Lee, M.H. Association of TXNDC5 gene polymorphisms and susceptibility to nonsegmental vitiligo in the Korean population. Br. J. Dermatol. 2010, 162, 759–764. [Google Scholar] [CrossRef]
- Victor, P.; Sarada, D.; Ramkumar, K.M. Crosstalk between endoplasmic reticulum stress and oxidative stress: Focus on protein disulfide isomerase and endoplasmic reticulum oxidase 1. Eur. J. Pharmacol. 2021, 892, 173749. [Google Scholar] [CrossRef]
- Isomoto, A.; Shoguchi, E.; Hisata, K.; Inoue, J.; Sun, Y.; Inaba, K.; Satoh, N.; Ogawa, T.; Shibata, H. Active expression of genes for protein modification enzymes in habu venom glands. Toxins 2022, 14, 300. [Google Scholar] [CrossRef]
- Horna-Terrón, E.; Pradilla-Dieste, A.; Sánchez-de-Diego, C.; Osada, J. TXNDC5, a newly discovered disulfide isomerase with a key role in cell physiology and pathology. Int. J. Mol. Sci. 2014, 15, 23501–23518. [Google Scholar] [CrossRef]
- Sandamalika, W.G.; Samaraweera, A.V.; Yang, H.; Lee, J. A newly discovered teleost disulfide isomerase, thioredoxin domain containing 5 (TXNDC5), from big-belly seahorse (Hippocampus abdominalis): Insights into its molecular and functional properties and immune regulatory functions. Dev. Comp. Immunol. 2021, 114, 103827. [Google Scholar] [CrossRef]
- Tassano, E.; Uccella, S.; Severino, M.; Giacomini, T.; Nardi, F.; Gimelli, G.; Tavella, E.; Ronchetto, P.; Malacarne, M.; Coviello, D. Expanding the phenotype associated with interstitial 6p25. 1p24. 3 microdeletion: A new case and review of the literature. J. Genet. 2021, 100, 9. [Google Scholar] [CrossRef]
- He, Y.; Yuan, C.; Chen, L.; Lei, M.; Zellmer, L.; Huang, H.; Liao, D.J. Transcriptional-readthrough RNAs reflect the phenomenon of “a gene contains gene (s)” or “gene (s) within a gene” in the human genome, and thus are not chimeric RNAs. Genes 2018, 9, 40. [Google Scholar] [CrossRef]
- Li, P.; Li, Y.; Ma, L. Potential role of chimeric genes in pathway-related gene co-expression modules. World J. Surg. Oncol. 2021, 19, 149. [Google Scholar] [CrossRef]
- Mallén-Ponce, M.J.; Huertas, M.J.; Florencio, F.J. Exploring the diversity of the thioredoxin systems in cyanobacteria. Antioxidants 2022, 11, 654. [Google Scholar] [CrossRef]
- Marcuello, C.; Frempong, G.A.; Balsera, M.; Medina, M.; Lostao, A. Atomic force microscopy to elicit conformational transitions of ferredoxin-dependent flavin thioredoxin reductases. Antioxidants 2021, 10, 1437. [Google Scholar] [CrossRef]
- Arai, K.; Ueno, H.; Asano, Y.; Chakrabarty, G.; Shimodaira, S.; Mugesh, G.; Iwaoka, M. Protein Folding in the Presence of Water-Soluble Cyclic Diselenides with Novel Oxidoreductase and Isomerase Activities. ChemBioChem 2018, 19, 207–211. [Google Scholar] [CrossRef]
- Cheng, F.; Ji, Q.; Wang, L.; Wang, C.c.; Liu, G.H.; Wang, L. Reducing oxidative protein folding alleviates senescence by minimizing ER-to-nucleus H2O2 release. EMBO Rep. 2023, 24, e56439. [Google Scholar] [CrossRef]
- Bak, D.W.; Gao, J.; Wang, C.; Weerapana, E. A quantitative chemoproteomic platform to monitor selenocysteine reactivity within a complex proteome. Cell Chem. Biol. 2018, 25, 1157–1167.e4. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Yang, K.; Wang, H.; Shergalis, A.; Kyani, A.; Bankhead III, A.; Tamura, S.; Yang, S.; Wang, X. Inhibition of protein disulfide isomerase in glioblastoma causes marked downregulation of DNA repair and DNA damage response genes. Theranostics 2019, 9, 2282. [Google Scholar] [CrossRef]
- George, G.; Ninagawa, S.; Yagi, H.; Saito, T.; Ishikawa, T.; Sakuma, T.; Yamamoto, T.; Imami, K.; Ishihama, Y.; Kato, K. EDEM2 stably disulfide-bonded to TXNDC11 catalyzes the first mannose trimming step in mammalian glycoprotein ERAD. eLife 2020, 9, e53455. [Google Scholar] [CrossRef]
- Matsusaki, M.; Okada, R.; Tanikawa, Y.; Kanemura, S.; Ito, D.; Lin, Y.; Watabe, M.; Yamaguchi, H.; Saio, T.; Lee, Y.-H.; et al. Functional interplay between p5 and pdi/erp72 to drive protein folding. Biology 2021, 10, 1112. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, Z.; Wang, W.; Jing, L.; Yu, Q.; Li, Z.; Chen, X.; Zhang, J.; Zhang, P.; Feng, F.; et al. LncRNA-ENST00000556926 regulates the proliferation, apoptosis and mRNA transcriptome of malignant-transformed BEAS-2B cells induced by coal tar pitch. Toxicol. Res. 2021, 10, 1144–1152. [Google Scholar] [CrossRef]
- Lu, H.; Gao, L.; Lv, J. Circ_0078710 promotes the development of liver cancer by upregulating TXNDC5 via miR-431-5p. Ann. Hepatol. 2022, 27, 100551. [Google Scholar] [CrossRef]
- Zang, H.; Li, Y.; Zhang, X.; Huang, G. Circ_0000517 contributes to hepatocellular carcinoma progression by upregulating TXNDC5 via sponging miR-1296-5p. Cancer Manag. Res. 2020, 12, 3457–3468. [Google Scholar] [CrossRef]
- Yu, J.; Yang, M.; Zhou, B.; Luo, J.; Zhang, Z.; Zhang, W.; Yan, Z. CircRNA-104718 acts as competing endogenous RNA and promotes hepatocellular carcinoma progression through microRNA-218-5p/TXNDC5 signaling pathway. Clin. Sci. 2019, 133, 1487–1503. [Google Scholar] [CrossRef]
- Chang, C.; Rajasekaran, M.; Qiao, Y.; Dong, H.; Wang, Y.; Xia, H.; Deivasigamani, A.; Wu, M.; Sekar, K.; Gao, H.; et al. The aberrant upregulation of exon 10-inclusive SREK1 through SRSF10 acts as an oncogenic driver in human hepatocellular carcinoma. Nat. Commun. 2022, 13, 1363. [Google Scholar] [CrossRef]
- Tavasolian, F.; Abdollahi, E.; Rezaei, R.; Momtazi-borojeni, A.A.; Henrotin, Y.; Sahebkar, A. Altered expression of microRNAs in rheumatoid arthritis. J. Cell. Biochem. 2018, 119, 478–487. [Google Scholar] [CrossRef]
- Wang, L.; Song, G.; Zheng, Y.; Wang, D.; Dong, H.; Pan, J.; Chang, X. miR-573 is a negative regulator in the pathogenesis of rheumatoid arthritis. Cell. Mol. Immunol. 2016, 13, 839–849. [Google Scholar] [CrossRef]
- Tenga, A.; Beard, J.A.; Takwi, A.; Wang, Y.-M.; Chen, T. Regulation of nuclear receptor Nur77 by miR-124. PLoS ONE 2016, 11, e0148433. [Google Scholar] [CrossRef]
- Wang, L.; Song, G.; Chang, X.; Tan, W.; Pan, J.; Zhu, X.; Liu, Z.; Qi, M.; Yu, J.; Han, B. The role of TXNDC5 in castration-resistant prostate cancer—Involvement of androgen receptor signaling pathway. Oncogene 2015, 34, 4735–4745. [Google Scholar] [CrossRef]
- Fang, Y. Precision Nanomedicine Targeting Novel Endothelial Mechano-sensing Mechanisms To Treat Vascular Diseases. Arterioscler. Thromb. Vasc. Biol. 2022, 42 (Suppl. S1), A139. [Google Scholar] [CrossRef]
- Zhao, J.; Schnitzler, G.R.; Iyer, L.K.; Aronovitz, M.J.; Baur, W.E.; Karas, R.H. MicroRNA-offset RNA alters gene expression and cell proliferation. PLoS ONE 2016, 11, e0156772. [Google Scholar] [CrossRef]
- Pang, X.; Wang, L.; Ma, H.; Zhang, Y.; Pan, J.; Chen, Y.; Du, B.; Wei, Q. Enhanced photoelectrochemical aptasensing platform for TXNDC5 gene based on exciton energy transfer between NCQDs and TiO2 nanorods. Sci. Rep. 2016, 6, 19202. [Google Scholar] [CrossRef]
- Yuan, K.; Xie, K.; Lan, T.; Xu, L.; Chen, X.; Li, X.; Liao, M.; Li, J.; Huang, J.; Zeng, Y.; et al. TXNDC12 promotes EMT and metastasis of hepatocellular carcinoma cells via activation of β-catenin. Cell Death Differ. 2020, 27, 1355–1368. [Google Scholar] [CrossRef]
- Duivenvoorden, W.; Federov, S.; Hopmans, S.; Pinthus, J. MP37-05 ERP46 mediates prostate cancer tumorigenesis. J. Urol. 2015, 193, e439. [Google Scholar] [CrossRef]
- Duivenvoorden, W.; Hopmans, S.N.; Austin, R.C.; Pinthus, J.H. Endoplasmic reticulum protein ERp46 in prostate adenocarcinoma. Oncol. Lett. 2017, 13, 3624–3630. [Google Scholar] [CrossRef]
- Pinthus, J.H.; Hopmans, S.N.; Federov, S.; Duivenvoorden, W.C. ERp46 (thioredoxin domain-containing protein 5, TXND5) promotes prostate cancer growth in vitro and in vivo. Cancer Res. 2015, 75 (Suppl. S15), 4960. [Google Scholar] [CrossRef]
- Jung, E.J.; Chung, K.H.; Kim, C.W. Identification of simvastatin-regulated targets associated with JNK activation in DU145 human prostate cancer cell death signaling. BMB Rep. 2017, 50, 466. [Google Scholar] [CrossRef]
- Wang, N.; Gu, Y.; Li, L.; Chi, J.; Liu, X.; Xiong, Y.; Jiang, S.; Zhang, W.; Zhong, C. Identification of novel prognostic risk signature of breast cancer based on ferroptosis-related genes. Sci. Rep. 2022, 12, 13766. [Google Scholar] [CrossRef]
- Hedrick, E.; Li, X.; Cheng, Y.; Lacey, A.; Mohankumar, K.; Zarei, M.; Safe, S. Potent inhibition of breast cancer by bis-indole-derived nuclear receptor 4A1 (NR4A1) antagonists. Breast Cancer Res. Treat. 2019, 177, 29–40. [Google Scholar] [CrossRef]
- Mohankumar, K.; Li, X.; Sridharan, S.; Karki, K.; Safe, S. Nuclear receptor 4A1 (NR4A1) antagonists induce ROS-dependent inhibition of mTOR signaling in endometrial cancer. Gynecol. Oncol. 2019, 154, 218–227. [Google Scholar] [CrossRef]
- Fan, G.; Wrzeszczynski, K.O.; Fu, C.; Su, G.; Pappin, D.J.; Lucito, R.; Tonks, N.K. A quantitative proteomics-based signature of platinum sensitivity in ovarian cancer cell lines. Biochem. J. 2015, 465, 433–442. [Google Scholar] [CrossRef]
- Zhang, L.; Mohankumar, K.; Martin, G.; Mariyam, F.; Park, Y.; Han, S.J.; Safe, S. Flavonoids Quercetin and Kaempferol Are NR4A1 Antagonists and Suppress Endometriosis in Female Mice. Endocrinology 2023, 164, bqad133. [Google Scholar] [CrossRef]
- Chiappetta, G.; Gamberi, T.; Faienza, F.; Limaj, X.; Rizza, S.; Messori, L.; Filomeni, G.; Modesti, A.; Vinh, J. Redox proteome analysis of auranofin exposed ovarian cancer cells (A2780). Redox Biol. 2022, 52, 102294. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, J.H.; Suk, J.M.; Lee, Y.I.; Kim, J.; Lee, J.H.; Lee, K.H. Identification of skin aging biomarkers correlated with the biomechanical properties. Ski. Res. Technol. 2021, 27, 940–947. [Google Scholar] [CrossRef]
- Azimi, A.; Kaufman, K.L.; Ali, M.; Arthur, J.; Kossard, S.; Fernandez-Penas, P. Differential proteomic analysis of actinic keratosis, Bowen’s disease and cutaneous squamous cell carcinoma by label-free LC–MS/MS. J. Dermatol. Sci. 2018, 91, 69–78. [Google Scholar] [CrossRef]
- Yue, Z.; Cao, M.; Hong, A.; Zhang, Q.; Zhang, G.; Jin, Z.; Zhao, L.; Wang, Q.; Fang, F.; Wang, Y.; et al. m6A Methyltransferase METTL3 Promotes the Progression of Primary Acral Melanoma via Mediating TXNDC5 Methylation. Front. Oncol. 2022, 11, 770325. [Google Scholar] [CrossRef]
- Andrews, J.M.; Schmidt, J.A.; Carson, K.R.; Musiek, A.C.; Mehta-Shah, N.; Payton, J.E. Novel cell adhesion/migration pathways are predictive markers of HDAC inhibitor resistance in cutaneous T cell lymphoma. EBioMedicine 2019, 46, 170–183. [Google Scholar] [CrossRef]
- Hu, Q.; Huang, K.; Tao, C.; Zhu, X. Protein disulphide isomerase can predict the clinical prognostic value and contribute to malignant progression in gliomas. J. Cell. Mol. Med. 2020, 24, 5888–5900. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, Y.; Cao, H.; Zou, H.; Wan, X.; Zeng, W.; Liu, Y.; Hu, J.; Zhang, N.; Xia, Z.; et al. Protein disulfide isomerases are promising targets for predicting the survival and tumor progression in glioma patients. Aging 2020, 12, 2347. [Google Scholar] [CrossRef]
- Kocatürk, B. Identification of thioredoxin domain containing family members’ expression pattern and prognostic value in diffuse gliomas via in silico analysis. Cancer Med. 2023, 12, 3830–3844. [Google Scholar] [CrossRef]
- Liu, X.; Yang, C.; Chen, P.; Zhang, L.; Cao, Y. The uses of transcriptomics and lipidomics indicated that direct contact with graphene oxide altered lipid homeostasis through ER stress in 3D human brain organoids. Sci. Total Environ. 2022, 849, 157815. [Google Scholar] [CrossRef] [PubMed]
- Yeste, N.; Pérez-Valle, J.; Vázquez-Gómez, M.; García-Contreras, C.; González-Bulnes, A.; Bassols, A. Polyphenols and IUGR pregnancies: Effects of the antioxidant hydroxytyrosol on the hippocampus proteome in a porcine model. Antioxidants 2022, 11, 1135. [Google Scholar] [CrossRef]
- Homma, T.; Fujiwara, H.; Osaki, T.; Fujii, S.; Fujii, J. Consequences of a peroxiredoxin 4 (Prdx4) deficiency on learning and memory in mice. Biochem. Biophys. Res. Commun. 2022, 621, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.; Zhu, H.; He, X.; Yu, N.; Zhang, X.; Xu, H.; Pei, H. Role of TXNDC5 in tumorigenesis of colorectal cancer cells: In vivo and in vitro evidence. Int. J. Mol. Med. 2018, 42, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Chen, C.; Yang, K. PO-299 Aberrant TXNDC5 expression in colon stromal fibroblasts promotes colorectal cancer carcinogenesis. ESMO Open 2018, 3, A344–A345. [Google Scholar] [CrossRef]
- Vieujean, S.; Hu, S.; Bequet, E.; Salee, C.; Massot, C.; Bletard, N.; Pierre, N.; Quesada Calvo, F.; Baiwir, D.; Mazzucchelli, G.; et al. Potential role of epithelial endoplasmic reticulum stress and anterior gradient protein 2 homologue in crohn’s disease fibrosis. J. Crohn’s Colitis 2021, 15, 1737–1750. [Google Scholar] [CrossRef]
- Zhang, L.; Martin, G.; Mohankumar, K.; Wright, G.A.; Mariyam, F.; Safe, S. Piperlongumine is a ligand for the orphan nuclear receptor 4A1 (NR4A1). Front. Pharmacol. 2023, 14, 1223153. [Google Scholar] [CrossRef]
- Hedrick, E.; Lee, S.o.; Safe, S. The nuclear orphan receptor NR4A1 regulates β1-integrin expression in pancreatic and colon cancer cells and can be targeted by NR4A1 antagonists. Mol. Carcinog. 2017, 56, 2066–2075. [Google Scholar] [CrossRef]
- Takagi, T.; Homma, T.; Fujii, J.; Shirasawa, N.; Yoriki, H.; Hotta, Y.; Higashimura, Y.; Mizushima, K.; Hirai, Y.; Katada, K.; et al. Elevated ER stress exacerbates dextran sulfate sodium-induced colitis in PRDX4-knockout mice. Free Radic. Biol. Med. 2019, 134, 153–164. [Google Scholar] [CrossRef] [PubMed]
- George, G.; Ninagawa, S.; Yagi, H.; Furukawa, J.-i.; Hashii, N.; Ishii-Watabe, A.; Deng, Y.; Matsushita, K.; Ishikawa, T.; Mamahit, Y.P. Purified EDEM3 or EDEM1 alone produces determinant oligosaccharide structures from M8B in mammalian glycoprotein ERAD. eLife 2021, 10, e70357. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.-Y.; Huo, F.-C.; Du, W.-Q.; Sun, X.-L.; Jiang, X.; Zhang, L.-S.; Pei, D.-S. METTL3 potentiates progression of cervical cancer by suppressing ER stress via regulating m6A modification of TXNDC5 mRNA. Oncogene 2022, 41, 4420–4432. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Zhang, L.; Shangguan, F.; Shi, G.; Wu, X.; Cui, Y.; Wang, X.; Liu, Y.; Lu, B.; et al. Targeting the functional interplay between endoplasmic reticulum oxidoreductin-1α and protein disulfide isomerase suppresses the progression of cervical cancer. eBioMedicine 2019, 41, 408–419. [Google Scholar] [CrossRef]
- Xu, B.; Li, J.; Liu, X.; Li, C.; Chang, X. TXNDC5 is a cervical tumor susceptibility gene that stimulates cell migration, vasculogenic mimicry and angiogenesis by down-regulating SERPINF1 and TRAF1 expression. Oncotarget 2017, 8, 91009. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.M.; Voitsitskyi, T.; Lee, J.-S. Organelle specific fluorescent phenomics and transcriptomic profiling to evaluate cellular response to tris (1, 3 dichloro 2 propyl) phosphate. Sci. Rep. 2022, 12, 4660. [Google Scholar] [CrossRef] [PubMed]
- Kanemura, S.; Okumura, M.; Yutani, K.; Ramming, T.; Hikima, T.; Appenzeller-Herzog, C.; Akiyama, S.; Inaba, K. Human ER oxidoreductin-1α (Ero1α) undergoes dual regulation through complementary redox interactions with protein-disulfide isomerase. J. Biol. Chem. 2016, 291, 23952–23964. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Li, D.-F.; Liang, J.; Sitia, R.; Wang, C.-c.; Wang, X. Crystal structure of the ERp44-peroxiredoxin 4 complex reveals the molecular mechanisms of thiol-mediated protein retention. Structure 2016, 24, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Kanemura, S.; Sofia, E.F.; Hirai, N.; Okumura, M.; Kadokura, H.; Inaba, K. Characterization of the endoplasmic reticulum–resident peroxidases GPx7 and GPx8 shows the higher oxidative activity of GPx7 and its linkage to oxidative protein folding. J. Biol. Chem. 2020, 295, 12772–12785. [Google Scholar] [CrossRef]
- Ushioda, R.; Nagata, K. Redox-mediated regulatory mechanisms of endoplasmic reticulum homeostasis. Cold Spring Harb. Perspect. Biol. 2019, 11, a033910. [Google Scholar] [CrossRef]
- Anelli, T.; Sannino, S.; Sitia, R. Proteostasis and “redoxtasis” in the secretory pathway: Tales of tails from ERp44 and immunoglobulins. Free Radic. Biol. Med. 2015, 83, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Moilanen, A.; Korhonen, K.; Saaranen, M.J.; Ruddock, L.W. Molecular analysis of human Ero1 reveals novel regulatory mechanisms for oxidative protein folding. Life Sci. Alliance 2018, 1, e201800090. [Google Scholar] [CrossRef] [PubMed]
- Matsusaki, M.; Okuda, A.; Matsuo, K.; Gekko, K.; Masuda, T.; Naruo, Y.; Hirose, A.; Kono, K.; Tsuchi, Y.; Urade, R. Regulation of plant ER oxidoreductin 1 (ERO1) activity for efficient oxidative protein folding. J. Biol. Chem. 2019, 294, 18820–18835. [Google Scholar] [CrossRef]
- Fujii, S.; Ushioda, R.; Nagata, K. Redox states in the endoplasmic reticulum directly regulate the activity of calcium channel, inositol 1, 4, 5-trisphosphate receptors. Proc. Natl. Acad. Sci. USA 2023, 120, e2216857120. [Google Scholar] [CrossRef] [PubMed]
- Chawsheen, H.A.; Jiang, H.; Ying, Q.; Ding, N.; Thapa, P.; Wei, Q. The redox regulator sulfiredoxin forms a complex with thioredoxin domain–containing 5 protein in response to ER stress in lung cancer cells. J. Biol. Chem. 2019, 294, 8991–9006. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, X.; Shan, Y.; Li, J.; Cui, W.; Wang, J.; Jiang, J.; Xie, Q.; Zhang, C.; Duan, C. Recognition of immune-related tumor antigens and immune subtypes for mRNA vaccine development in lung adenocarcinoma. Comput. Struct. Biotechnol. J. 2022, 20, 5001–5013. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Yeh, C.-F.; Lee, Y.-T.; Shih, Y.-C.; Chen, Y.-T.; Hung, C.-T.; You, M.-Y.; Wu, P.-C.; Shentu, T.-P.; Huang, R.-T.; et al. Fibroblast-enriched endoplasmic reticulum protein TXNDC5 promotes pulmonary fibrosis by augmenting TGFβ signaling through TGFBR1 stabilization. Nat. Commun. 2020, 11, 4254. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, C.; Liu, J.; He, Y.; Wei, Y.; Chen, J. Inhibition of ER stress by targeting the IRE1α–TXNDC5 pathway alleviates crystalline silica-induced pulmonary fibrosis. Int. Immunopharmacol. 2021, 95, 107519. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Zhang, Z.; Lu, Z.; Xiong, J.; Fan, T.; Song, C.; He, R.; Zhang, L.; Pan, S.; Li, D.; et al. Single-cell RNA sequencing analysis revealed cellular and molecular immune profiles in lung squamous cell carcinoma. Transl. Oncol. 2023, 27, 101568. [Google Scholar] [CrossRef]
- Chawsheen, H.A. Role of Sulfiredoxin Interacting Proteins in Lung Cancer Development. Ph.D. Thesis, University of Kentucky, Lexington, KY, USA, 2016. [Google Scholar]
- Hou, Y.; Wang, L.-J.; Jin, Y.-H.; Guo, R.-Y.; Yang, L.; Li, E.-C.; Zhang, J.-L. Triphenyltin exposure induced abnormal morphological colouration in adult male guppies (Poecilia reticulata). Ecotoxicol. Environ. Saf. 2022, 242, 113912. [Google Scholar] [CrossRef]
- Lee, T.; Lee, Y.; Lu, F.; Tsao, P.; Fang, Y.; Yang, K. Endoplasmic Reticulum Protein Thioredoxin Domain Containing 5 (TXNDC5) as a Novel Therapeutic Target for Pulmonary Fibrosis. In C28 Pathways of Lung Morphogenesis; American Thoracic Society: New York, NY, USA, 2019; p. A4431. [Google Scholar]
- Batool, S.; Bin-T-Abid, D.; Batool, H.; Shahid, S.; Saleem, M.; Khan, A.U.; Hamid, A.; Mahmood, M.S.; Ashraf, N.M. Development of multi-epitope vaccine constructs for non-small cell lung cancer (NSCLC) against USA human leukocyte antigen background: An immunoinformatic approach toward future vaccine designing. Expert Opin. Biol. Ther. 2021, 21, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, E.; Mohankumar, K.; Lacey, A.; Safe, S. Inhibition of NR4A1 promotes ROS accumulation and IL24-dependent growth arrest in rhabdomyosarcoma. Mol. Cancer Res. 2019, 17, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Lacey, A.; Hedrick, E.; Li, X.; Patel, K.; Doddapaneni, R.; Singh, M.; Safe, S. Nuclear receptor 4A1 (NR4A1) as a drug target for treating rhabdomyosarcoma (RMS). Oncotarget 2016, 7, 31257. [Google Scholar] [CrossRef] [PubMed]
- Oka, O.B.; Yeoh, H.Y.; Bulleid, N.J. Thiol-disulfide exchange between the PDI family of oxidoreductases negates the requirement for an oxidase or reductase for each enzyme. Biochem. J. 2015, 469, 279–288. [Google Scholar] [CrossRef]
- Yim, S.H.; Everley, R.A.; Schildberg, F.A.; Lee, S.-G.; Orsi, A.; Barbati, Z.R.; Karatepe, K.; Fomenko, D.E.; Tsuji, P.A.; Luo, H.R.; et al. Role of Selenof as a gatekeeper of secreted disulfide-rich glycoproteins. Cell Rep. 2018, 23, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-T.; Jhao, P.-Y.; Hung, C.-T.; Wu, Y.-F.; Lin, S.-J.; Chiang, W.-C.; Lin, S.-L.; Yang, K.-C. Endoplasmic reticulum protein TXNDC5 promotes renal fibrosis by enforcing TGF-β signaling in kidney fibroblasts. J. Clin. Investig. 2021, 131, e143645. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, E.; Lee, S.-O.; Kim, G.; Abdelrahim, M.; Jin, U.-H.; Safe, S.; Abudayyeh, A. Nuclear receptor 4A1 (NR4A1) as a drug target for renal cell adenocarcinoma. PLoS ONE 2015, 10, e0128308. [Google Scholar] [CrossRef] [PubMed]
- Mo, R.; Peng, J.; Xiao, J.; Ma, J.; Li, W.; Wang, J.; Ruan, Y.; Ma, S.; Hong, Y.; Wang, C.; et al. High TXNDC5 expression predicts poor prognosis in renal cell carcinoma. Tumor Biol. 2016, 37, 9797–9806. [Google Scholar] [CrossRef] [PubMed]
- Wrzesiński, T.; Szelag, M.; Cieślikowski, W.A.; Ida, A.; Giles, R.; Zodro, E.; Szumska, J.; Poźniak, J.; Kwias, Z.; Bluyssen, H.A.; et al. Expression of pre-selected TMEMs with predicted ER localization as potential classifiers of ccRCC tumors. BMC Cancer 2015, 15, 518. [Google Scholar] [CrossRef]
- Luo, L.; Wang, S.; Hu, Y.; Wang, L.; Jiang, X.; Zhang, J.; Liu, X.; Guo, X.; Luo, Z.; Zhu, C.; et al. Precisely Regulating M2 Subtype Macrophages for Renal Fibrosis Resolution. ACS Nano 2023, 17, 22508–22526. [Google Scholar] [CrossRef]
- Oka, O.B.; van Lith, M.; Rudolf, J.; Tungkum, W.; Pringle, M.A.; Bulleid, N.J. ER p18 regulates activation of ATF 6α during unfolded protein response. EMBO J. 2019, 38, e100990. [Google Scholar] [CrossRef]
- Paxman, R.; Plate, L.; Blackwood, E.A.; Glembotski, C.; Powers, E.T.; Wiseman, R.L.; Kelly, J.W. Pharmacologic ATF6 activating compounds are metabolically activated to selectively modify endoplasmic reticulum proteins. eLife 2018, 7, e37168. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Cho, Y.; Lee, H.; Joo, D.; Huh, K.; Kim, M.; Kim, Y. Comparative proteomic analysis of rapamycin versus cyclosporine combination treatment in mouse podocytes. Transplant. Proc. 2016, 48, 1297–1301. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhu, D.Y.; Xie, X.M.; Ding, M.; Wang, Y.L.; Sun, L.L.; Zhang, N.; Shen, E.; Wang, X.X. EA15, MIR22, LINC00472 as diagnostic markers for diabetic kidney disease. J. Cell. Physiol. 2019, 234, 8797–8803. [Google Scholar] [CrossRef] [PubMed]
- Kikuma, T.; Ibuki, H.; Nakamoto, M.; Seko, A.; Ito, Y.; Takeda, Y. In vitro mannosidase activity of EDEM3 against asparagine-linked oligomannose-type glycans. Biochem. Biophys. Res. Commun. 2022, 612, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Ito, S.; Wada, I.; Hosokawa, N. ER-resident protein 46 (ERp46) triggers the mannose-trimming activity of ER degradation–enhancing α-mannosidase–like protein 3 (EDEM3). J. Biol. Chem. 2018, 293, 10663–10674. [Google Scholar] [CrossRef] [PubMed]
- Shenkman, M.; Ron, E.; Yehuda, R.; Benyair, R.; Khalaila, I.; Lederkremer, G.Z. Mannosidase activity of EDEM1 and EDEM2 depends on an unfolded state of their glycoprotein substrates. Commun. Biol. 2018, 1, 172. [Google Scholar] [CrossRef] [PubMed]
- Kemter, E.; Frohlich, T.; Arnold, G.; Wolf, E.; Wanke, R. Mitochondrial dysregulation secondary to endoplasmic reticulum stress in autosomal dominant tubulointerstitial kidney disease-UMOD (ADTKD-UMOD). Sci. Rep. 2017, 7, 42970. [Google Scholar] [CrossRef]
- Fujimori, T.; Suno, R.; Iemura, S.I.; Natsume, T.; Wada, I.; Hosokawa, N. Endoplasmic reticulum proteins SDF 2 and SDF 2L1 act as components of the BiP chaperone cycle to prevent protein aggregation. Genes Cells 2017, 22, 684–698. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, R.; Wang, F.; Li, G.; Wen, Y.; Shan, H. Comparative proteomic analysis of PK15 swine kidney cells infected with a pseudorabies pathogenic variant and the Bartha-K/61 vaccine strain. Microb. Pathog. 2022, 170, 105698. [Google Scholar] [CrossRef]
- Mideksa, Y.G.; Aschenbrenner, I.; Fux, A.; Kaylani, D.; Weiß, C.A.; Nguyen, T.-A.; Bach, N.C.; Lang, K.; Sieber, S.A.; Feige, M.J. A comprehensive set of ER protein disulfide isomerase family members supports the biogenesis of proinflammatory interleukin 12 family cytokines. J. Biol. Chem. 2022, 298, 102677. [Google Scholar] [CrossRef] [PubMed]
- Nassar, A.; El-Rauf, A.; Mohammed, S.; Mostafa, A. Evaluation of Serum Level of Thioredoxin and its Gene Polymorphism in Diagnosis of Hepatocellular Carcinoma of HCV-Infected Patients. Benha Med. J. 2020, 37, 111–130. [Google Scholar] [CrossRef]
- Hung, C.-T.; Su, T.-H.; Chen, Y.-T.; Wu, Y.-F.; Chen, Y.-T.; Lin, S.-J.; Lin, S.-L.; Yang, K.-C. Targeting ER protein TXNDC5 in hepatic stellate cell mitigates liver fibrosis by repressing non-canonical TGFβ signalling. Gut 2022, 71, 1876–1891. [Google Scholar] [CrossRef] [PubMed]
- Durcan, E.; Sağlam, E.; Dağ, İ.; Dinç, M.; Andaç, Ş. Clinical Utility of Hepassocin and TXNDC5 in Patients with Non-alcoholic Fatty Liver Disease and/or Type 2 Diabetes. Bagcilar Med. Bull./Bağcılar Tıp Bülteni 2023, 8, 355–362. [Google Scholar] [CrossRef]
- Sánchez-Marco, J.; Martínez-Beamonte, R.; Diego, A.D.; Herrero-Continente, T.; Barranquero, C.; Arnal, C.; Surra, J.; Navarro, M.A.; Osada, J. Thioredoxin Domain Containing 5 Suppression Elicits Serum Amyloid A-Containing High-Density Lipoproteins. Biomedicines 2022, 10, 709. [Google Scholar] [CrossRef] [PubMed]
- Na, W.; Wu, Y.-Y.; Gong, P.-F.; Wu, C.-Y.; Cheng, B.-H.; Wang, Y.-X.; Wang, N.; Du, Z.-Q.; Li, H. Embryonic transcriptome and proteome analyses on hepatic lipid metabolism in chickens divergently selected for abdominal fat content. BMC Genom. 2018, 19, 384. [Google Scholar] [CrossRef] [PubMed]
- Bidooki, S.H.; Sánchez-Marco, J.; Martínez-Beamonte, R.; Herrero-Continente, T.; Navarro, M.A.; Rodríguez-Yoldi, M.J.; Osada, J. Endoplasmic Reticulum Protein TXNDC5 Interacts with PRDX6 and HSPA9 to Regulate Glutathione Metabolism and Lipid Peroxidation in the Hepatic AML12 Cell Line. Int. J. Mol. Sci. 2023, 24, 17131. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Lyu, M.; Zhang, Z.; Miyagi, M.; Huang, W.; Taylor, D.J.; Edward, W.Y. High-resolution structural-omics of human liver enzymes. Cell Rep. 2023, 42, 112609. [Google Scholar] [CrossRef] [PubMed]
- Kern, P.; Balzer, N.R.; Bender, F.; Frolov, A.; Wunderling, K.; Sowa, J.-P.; Bonaguro, L.; Ulas, T.; Thiele, C.; Schultze, J.L. Creld2 function during unfolded protein response is essential for liver metabolism homeostasis. FASEB J. 2021, 35, e21939. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, J.; Wu, H.; Tian, H.; Song, D.; Wu, W.; Dong, F. Knockdown of TXNDC5 alleviates CCL4-induced hepatic fibrosis in mice by enhancing endoplasmic reticulum stress. Am. J. Med. Sci. 2023, 366, 449–457. [Google Scholar] [CrossRef]
- Bidooki, S.H.; Alejo, T.; Sánchez-Marco, J.; Martínez-Beamonte, R.; Abuobeid, R.; Burillo, J.C.; Lasheras, R.; Sebastian, V.; Rodríguez-Yoldi, M.J.; Arruebo, M.; et al. Squalene loaded nanoparticles effectively protect hepatic AML12 cell lines against oxidative and endoplasmic reticulum stress in a TXNDC5-dependent way. Antioxidants 2022, 11, 581. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Marco, J.; Bidooki, S.H.; Abuobeid, R.; Barranquero, C.; Herrero-Continente, T.; Arnal, C.; Martínez-Beamonte, R.; Lasheras, R.; Surra, J.C.; Navarro, M.A.; et al. Thioredoxin domain containing 5 is involved in the hepatic storage of squalene into lipid droplets in a sex-specific way. J. Nutr. Biochem. 2024, 124, 109503. [Google Scholar] [CrossRef] [PubMed]
- Abuobeid, R.; Herrera-Marcos, L.V.; Arnal, C.; Bidooki, S.H.; Sánchez-Marco, J.; Lasheras, R.; Surra, J.C.; Rodríguez-Yoldi, M.J.; Martínez-Beamonte, R.; Osada, J. Differentially Expressed Genes in Response to a Squalene-Supplemented Diet Are Accurate Discriminants of Porcine Non-Alcoholic Steatohepatitis. Int. J. Mol. Sci. 2023, 24, 12552. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Xu, J.; Zhao, J.; Yin, W.; Liu, D.; Chen, W.; Hou, S.X. Arf1-mediated lipid metabolism sustains cancer cells and its ablation induces anti-tumor immune responses in mice. Nat. Commun. 2020, 11, 220. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhou, B.; Dong, C.; Zhang, Y.; Yu, J.; Pu, Y.; Feng, W.; Sun, L.; Hu, H.; Chen, Y.; et al. CRISPR/Cas9-2D silicene gene-editing nanosystem for remote nir-ii-induced tumor microenvironment reprogramming and augmented photonic tumor ablation. Adv. Funct. Mater. 2021, 31, 2107093. [Google Scholar] [CrossRef]
- Ao, N.; Yang, J.; Wang, X.; Du, J. Glucagon-like peptide-1 preserves non-alcoholic fatty liver disease through inhibition of the endoplasmic reticulum stress-associated pathway. Hepatol. Res. 2016, 46, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Karatas, E.; Raymond, A.-A.; Leon, C.; Dupuy, J.-W.; Di-Tommaso, S.; Senant, N.; Collardeau-Frachon, S.; Ruiz, M.; Lachaux, A.; Saltel, F.; et al. Hepatocyte proteomes reveal the role of protein disulfide isomerase 4 in alpha 1-antitrypsin deficiency. JHEP Rep. 2021, 3, 100297. [Google Scholar] [CrossRef] [PubMed]
- AO, N.; Yang, J.; Zheng, L.; MA, J.; DU, J. The influence of liraglutide on ERp46 in non-alcoholic fatty liver disease rats. Chin. J. Endocrinol. Metab. 2018, 34, 144–148. [Google Scholar]
- Zhang, X.; Wang, G.; Guo, S. Effect of hydrogen on endoplasmic reticulum stress during liver cold ischemia-reperfusion in rats. Chin. J. Anesthesiol. 2017, 12, 450–453. [Google Scholar]
- Broering, R.; Trippler, M.; Werner, M.; Real, C.; Megger, D.; Bracht, T.; Schweinsberg, V.; Sitek, B.; Eisenacher, M.; Meyer, H. Hepatic expression of proteasome subunit alpha type-6 is upregulated during viral hepatitis and putatively regulates the expression of ISG 15 ubiquitin-like modifier, a proviral host gene in hepatitis C virus infection. J. Viral Hepat. 2016, 23, 375–386. [Google Scholar] [CrossRef]
- Nasser, S.; Christofferson, A.; Legendre, C.; Aldrich, J.; Benard, B.; Network, T.M.C.; Yesil, J.; Auclair, D.; Liang, W.; Lonial, S. Comprehensive identification of fusion transcripts in multiple myeloma: An Mmrf commpass analysis. Blood 2017, 130, 61. [Google Scholar]
- Chakrabortty, A.; Kumar, H.; Pfitzer, J.; Sharma, N.; Waliagha, R.; Mukherjee, U.; Kumar, S.K.; Baughn, L.B.; Van Ness, B.; Mitra, A.K. Single-Cell Multi-Ome Analysis Reveals Novel Molecular Mechanisms Underlying Subclonal Response to Survivin Inhibition in Relapsed/Refractory Multiple Myeloma. Blood 2023, 142, 6616. [Google Scholar] [CrossRef]
- Dytfeld, D.; Luczak, M.; Wrobel, T.; Usnarska-Zubkiewicz, L.; Brzezniakiewicz, K.; Jamroziak, K.; Giannopoulos, K.; Przybylowicz-Chalecka, A.; Ratajczak, B.; Czerwinska-Rybak, J.; et al. Comparative proteomic profiling of refractory/relapsed multiple myeloma reveals biomarkers involved in resistance to bortezomib-based therapy. Oncotarget 2016, 7, 56726. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Jayasinghe, R.G.; Wang, T.; O’Neal, J.; Liu, R.; Rettig, M.P.; Tsai, C.-F.; Foltz, S.M.; Kohnen, D.; Mani, S.; et al. Myeloma Cell Associated Therapeutic Protein Discovery Using Single Cell RNA-Seq Data. Blood 2020, 136, 4–5. [Google Scholar] [CrossRef]
- Barwick, B.G.; Neri, P.; Bahlis, N.J.; Nooka, A.K.; Dhodapkar, M.V.; Jaye, D.L.; Hofmeister, C.C.; Kaufman, J.L.; Gupta, V.A.; Auclair, D.; et al. Multiple myeloma immunoglobulin lambda translocations portend poor prognosis. Nat. Commun. 2019, 10, 1911. [Google Scholar] [CrossRef] [PubMed]
- Vo, J.N.; Wu, Y.-M.; Mishler, J.; Hall, S.; Mannan, R.; Wang, L.; Ning, Y.; Zhou, J.; Hopkins, A.C.; Estill, J.C.; et al. The genetic heterogeneity and drug resistance mechanisms of relapsed refractory multiple myeloma. Nat. Commun. 2022, 13, 3750. [Google Scholar] [CrossRef]
- Cleynen, A.; Szalat, R.; Kemal Samur, M.; Robiou du Pont, S.; Buisson, L.; Boyle, E.; Chretien, M.; Anderson, K.; Minvielle, S.; Moreau, P.; et al. Expressed fusion gene landscape and its impact in multiple myeloma. Nat. Commun. 2017, 8, 1893. [Google Scholar] [CrossRef] [PubMed]
- Beycan, A.; Demiralp, D.F.O.; Dalva, K.; Beksaç, M. Comparative Proteomic Analysis of Bone Marrow Plasma Cells By Using Mass Spectrometry Based Bottom up Proteomic Strategies. Blood 2015, 126, 5334. [Google Scholar] [CrossRef]
- Foltz, S.M.; Gao, Q.; Yoon, C.J.; Weerasinghe, A.; Sun, H.; Yao, L.; Fiala, M.A.; Kohnen, D.R.; King, J.; Ju, Y.S.; et al. Comprehensive Multi-Omics Analysis of Gene Fusions in a Large Multiple Myeloma Cohort. Blood 2018, 132, 1898. [Google Scholar] [CrossRef]
- Poos, A.M.; Giesen, N.; Catalano, C.; Paramasivam, N.; Huebschmann, D.; John, L.; Baumann, A.; Hochhaus, J.; Mueller-Tidow, C.; Sauer, S.; et al. Comprehensive comparison of early relapse and end-stage relapsed refractory multiple myeloma. Blood 2020, 136, 1. [Google Scholar] [CrossRef]
- Misund, K.; Keane, N.; Asmann, Y.W.; Van Wier, S.; Riggs, D.; Ahmann, G.; Chesi, M.; Körtum, K.M.; Fonseca, R.; Stewart, A.K.; et al. Complementary activation of Ccnd, MYC, RAS and NFkB by mutations in multiple myeloma. Blood 2016, 128, 355. [Google Scholar] [CrossRef]
- Ashby, C.; Bauer, M.A.; Wang, Y.; Wardell, C.P.; Tytarenko, R.G.; Patel, P.; Flynt, E.; Ortiz, M.; Dervan, A.; Gockley, J.; et al. Chromothripsis and chromoplexy are associated with DNA instability and adverse clinical outcome in multiple myeloma. Blood 2018, 132, 408. [Google Scholar] [CrossRef]
- Liu, E.; Becker, N.; Sudha, P.; Mikulasova, A.; Zaid, M.A.; Suvannasankha, A.; Lee, K.P.; Abonour, R.; Walker, B.A. Unraveling Diverse Mechanisms of Complex Structural Variant Interactions through Multiomic Data in Multiple Myeloma. Blood 2023, 142, 641. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, G.; Seupel, R.; Feineis, D.; Bruennert, D.; Chatterjee, M.; Schlosser, A.; Bringmann, G. Epoxides related to dioncoquinone B: Synthesis, activity against multiple myeloma cells, and search for the target protein. Tetrahedron 2018, 74, 5102–5112. [Google Scholar] [CrossRef]
- Blaney, P.; Walker, B.A.; Mikulasova, A.; Gagler, D.; Ziccheddu, B.; Diamond, B.; Wang, Y.; Hou, L.; Liu, S.; Poos, A.M.; et al. Somatic Hypermutation in Enhancer Regions Shapes Non-Coding Myeloma Genome, Generating DNA Breaks and Driving Etiology through Mutation and Structural Variation. Blood 2023, 142, 1401. [Google Scholar] [CrossRef]
- Mikulasova, A.; Ashby, C.; Tytarenko, R.G.; Qu, P.; Rosenthal, A.; Dent, J.A.; Ryan, K.R.; Bauer, M.A.; Wardell, C.P.; Hoering, A.; et al. Microhomology-mediated end joining drives complex rearrangements and overexpression of MYC and PVT1 in multiple myeloma. Haematologica 2020, 105, 1055. [Google Scholar] [CrossRef] [PubMed]
- Ashby, C.; Boyle, E.M.; Bauer, M.A.; Mikulasova, A.; Wardell, C.P.; Williams, L.; Siegel, A.; Blaney, P.; Braunstein, M.; Kaminetsky, D.; et al. Structural variants shape the genomic landscape and clinical outcome of multiple myeloma. Blood Cancer J. 2022, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.A.; Chavan, S.S.; He, J.; Tytarenko, R.; Zhong, S.; Deshpande, S.; Patel, P.; Stein, C.K.; Stephens, O.W.; Weinhold, N.; et al. A Survey of Fusion Genes in Myeloma Identifies Kinase Domain Activation Which Could Be Targeted with Available Treatments; American Society of Hematology: Washington, DC, USA, 2016. [Google Scholar]
- Robinson, R.M.; Reyes, L.; Duncan, R.; Dolloff, N. Specifically targeting PDIA1 with indene inhibitors leads to bortezomib-potentiation in multiple myeloma. Cancer Res. 2019, 79 (Suppl. S13), 3863. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, L.; Li, N.; Sha, L.; Zhang, K. An immunohistochemical study of thioredoxin domain-containing 5 expression in gastric adenocarcinoma. Oncol. Lett. 2015, 9, 1154–1158. [Google Scholar] [CrossRef]
- Wu, Y.; Ye, H.; Peng, B.; Jiang, H.; Tang, Q.; Liu, Y.; Xi, J.; Chen, S. MiR-643 functions as a potential tumor suppressor in gastric cancer by inhibiting cell proliferation and invasion via targeting TXNDC9. Ann. Clin. Lab. Sci. 2021, 51, 494–502. [Google Scholar]
- Wang, H.; Yang, X.; Guo, Y.; Shui, L.; Li, S.; Bai, Y.; Liu, Y.; Zeng, M.; Xia, J. HERG1 promotes esophageal squamous cell carcinoma growth and metastasis through TXNDC5 by activating the PI3K/AKT pathway. J. Exp. Clin. Cancer Res. 2019, 38, 324. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zheng, Y.; Tang, J.; Wang, D.; Ma, Z.; Li, S.; Chang, X. Stimulation of DC-CIK with PADI4 protein can significantly elevate the therapeutic efficiency in esophageal cancer. J. Immunol. Res. 2019, 2019, 8587570. [Google Scholar] [CrossRef] [PubMed]
- Worah, K.; Mathan, T.S.; Manh, T.P.V.; Keerthikumar, S.; Schreibelt, G.; Tel, J.; Duiveman-de Boer, T.; Sköld, A.E.; van Spriel, A.B.; de Vries, I.J.M.; et al. Proteomics of human dendritic cell subsets reveals subset-specific surface markers and differential inflammasome function. Cell Rep. 2016, 16, 2953–2966. [Google Scholar] [CrossRef] [PubMed]
- Pi, Y.; Liang, Z.; Jiang, Q.; Chen, D.; Su, Z.; Ouyang, Y.; Zhang, Z.; Liu, J.; Wen, S.; Yang, L.; et al. The role of PIWI-interacting RNA in naringin pro-angiogenesis by targeting HUVECs. Chem.-Biol. Interact. 2023, 371, 110344. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Safe, S.; Lee, S.-O. Inactivation of the orphan nuclear receptor NR4A1 contributes to apoptosis induction by fangchinoline in pancreatic cancer cells. Toxicol. Appl. Pharmacol. 2017, 332, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Olsen, P.A.; Lund, K.; Krauss, S. Expression profiling of wild type and β-catenin gene disrupted human BxPC-3 pancreatic adenocarcinoma cells. Genom. Data 2015, 4, 150–152. [Google Scholar] [CrossRef]
- Safe, S.; Shrestha, R.; Mohankumar, K.; Howard, M.; Hedrick, E.; Abdelrahim, M. Transcription factors specificity protein and nuclear receptor 4A1 in pancreatic cancer. World J. Gastroenterol. 2021, 27, 6387. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Kim, S.-H.; Kim, B.-M.; Safe, S.; Lee, S.-O. Broussochalcone A is a novel inhibitor of the orphan nuclear receptor NR4A1 and induces apoptosis in pancreatic cancer cells. Molecules 2021, 26, 2316. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Zhang, H.; Du, Y.; Tan, P. Cetuximab enhances cisplatin-induced endoplasmic reticulum stress-associated apoptosis in laryngeal squamous cell carcinoma cells by inhibiting expression of TXNDC5. Mol. Med. Rep. 2018, 17, 4767–4776. [Google Scholar] [CrossRef]
- Wynn, T. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef]
- Shih, Y.-C.; Chen, C.-L.; Zhang, Y.; Mellor, R.L.; Kanter, E.M.; Fang, Y.; Wang, H.-C.; Hung, C.-T.; Nong, J.-Y.; Chen, H.-J.; et al. Endoplasmic reticulum protein TXNDC5 augments myocardial fibrosis by facilitating extracellular matrix protein folding and redox-sensitive cardiac fibroblast activation. Circ. Res. 2018, 122, 1052–1068. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, B.; Wu, C.; Yan, X.; Zhang, L.; Chang, X. TXNDC5 contributes to rheumatoid arthritis by down-regulating IGFBP1 expression. Clin. Exp. Immunol. 2018, 192, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wang, J.; Zhang, X.; Tian, R.; Qiao, L.; Ge, L.; Pan, J.; Wang, L. TXNDC5 protects synovial fibroblasts of rheumatoid arthritis from the detrimental effects of endoplasmic reticulum stress. Intractable Rare Dis. Res. 2020, 9, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Y.; Liu, Q.; Tian, Z.; Zhang, Y. Mechanistic and therapeutic links between rheumatoid arthritis and diabetes mellitus. Clin. Exp. Med. 2023, 23, 287–299. [Google Scholar] [CrossRef] [PubMed]
- de Seny, D.; Bianchi, E.; Baiwir, D.; Cobraiville, G.; Collin, C.; Deliège, M.; Kaiser, M.-J.; Mazzucchelli, G.; Hauzeur, J.-P.; Delvenne, P.; et al. Proteins involved in the endoplasmic reticulum stress are modulated in synovitis of osteoarthritis, chronic pyrophosphate arthropathy and rheumatoid arthritis, and correlate with the histological inflammatory score. Sci. Rep. 2020, 10, 14159. [Google Scholar] [CrossRef]
- Nakao, H.; Seko, A.; Ito, Y.; Sakono, M. PDI family protein ERp29 recognizes P-domain of molecular chaperone calnexin. Biochem. Biophys. Res. Commun. 2017, 487, 763–767. [Google Scholar] [CrossRef]
- Lan, H.; Tong, Z.; Jiao, Y.; Han, H.; Ma, Y.; Li, Y.; Jia, X.; Hu, B.; Zhang, W.; Zhong, M.; et al. Deep Vein Thrombosis Is Facilitated by Endothelial-Derived Extracellular Vesicles via the PDI–GRP94–GPIIb/IIIa Pathway in Mice. J. Clin. Med. 2023, 12, 4265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, R.; Ge, L.; Wang, L. Exosome-derived TXNDC5 is Required for the Inflammatory Progression of Rheumatoid Arthritis Fibroblast-like Synoviocytes. Explor. Res. Hypothesis Med. 2021, 6, 147–155. [Google Scholar] [CrossRef]
- Yuan, H.-Y.; Zhang, X.-L.; Zhang, X.-H.; Meng, L.; Wei, J.-F. Analysis of patents on anti-rheumatoid arthritis therapies issued in China. Expert Opin. Ther. Pat. 2015, 25, 909–930. [Google Scholar] [CrossRef]
- Wang, L.; Dong, H.; Song, G.; Zhang, R.; Pan, J.; Han, J. TXNDC5 synergizes with HSC70 to exacerbate the inflammatory phenotype of synovial fibroblasts in rheumatoid arthritis through NF-κB signaling. Cell. Mol. Immunol. 2018, 15, 685–696. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Jiang, H.; Chang, X.; Pan, J. Inhibition of PCSK6 may play a protective role in the development of rheumatoid arthritis. J. Rheumatol. 2015, 42, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Li, J.; Wu, C.; Liu, C.; Yan, X.; Chang, X. CXCL10 and TRAIL are upregulated by TXNDC5 in rheumatoid arthritis fibroblast-like synoviocytes. J. Rheumatol. 2018, 45, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, E.; Lymperopoulou, A.; Charonis, A. Reduced expression of ERp46 under diabetic conditions in β-cells and the effect of liraglutide. Metabolism 2016, 65, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Li, A.H.; Morrison, A.C.; Kovar, C.; Cupples, L.A.; Brody, J.A.; Polfus, L.M.; Yu, B.; Metcalf, G.; Muzny, D.; Veeraraghavan, N.; et al. Analysis of loss-of-function variants and 20 risk factor phenotypes in 8554 individuals identifies loci influencing chronic disease. Nat. Genet. 2015, 47, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Turkyilmaz, A.; Lee, Y.; Lee, M. Fermented extract of mealworm (Tenebrio molitor larvae) as a dietary protein source modulates hepatic proteomic profiles in C57BLKS/J-db/db mice. J. Insects Food Feed 2023, 9, 1199–1210. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Saito, M.; Kadokura, H.; Miyazaki, J.-i.; Tashiro, F.; Imagawa, Y.; Iwawaki, T.; Kohno, K. IRE1–XBP1 pathway regulates oxidative proinsulin folding in pancreatic β cells. J. Cell Biol. 2018, 217, 1287–1301. [Google Scholar] [CrossRef]
- Yeh, C.-F.; Cheng, S.-H.; Lin, Y.-S.; Shentu, T.-P.; Huang, R.-T.; Zhu, J.; Chen, Y.-T.; Kumar, S.; Lin, M.-S.; Kao, H.-L.; et al. Targeting mechanosensitive endothelial TXNDC5 to stabilize eNOS and reduce atherosclerosis in vivo. Sci. Adv. 2022, 8, eabl8096. [Google Scholar] [CrossRef]
- Cheng, S.; Yeh, C.; Fang, Y.; Yang, K. P593 Endoplasmic reticulum protein thioredoxin domain containing 5 (TXNDC5) is a novel mediator of endothelial dysfunction and atherosclerosis. Eur. Heart J. 2018, 39 (Suppl. S1), 71–72. [Google Scholar] [CrossRef]
- TXNDC5 is a novel therapeutic target of atrial fibrosis and fibrillation. Eur. Heart J. 2019, 40, ehz748.0884. [CrossRef]
- Kim, Y.-M.; Youn, S.-W.; Sudhahar, V.; Das, A.; Chandhri, R.; Grajal, H.C.; Kweon, J.; Leanhart, S.; He, L.; Toth, P.T.; et al. Redox regulation of mitochondrial fission protein Drp1 by protein disulfide isomerase limits endothelial senescence. Cell Rep. 2018, 23, 3565–3578. [Google Scholar] [CrossRef]
- García-Vilas, J.A.; Martínez-Poveda, B.; Quesada, A.R.; Medina, M.Á. (+)-Aeroplysinin-1 modulates the redox balance of endothelial cells. Mar. Drugs 2018, 16, 316. [Google Scholar] [CrossRef]
- Marynowska, M.; Herosimczyk, A.; Lepczyński, A.; Barszcz, M.; Konopka, A.; Dunisławska, A.; Ożgo, M. Gene and Protein Accumulation Changes Evoked in Porcine Aorta in Response to Feeding with Two Various Fructan Sources. Animals 2022, 12, 3147. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, Y.; Rauova, L.; Koma, G.; Wang, L.; Poncz, M.; Li, H.; Liu, T.; Fong, K.P.; Bennett, J.S.; et al. A novel role for endoplasmic reticulum protein 46 (ERp46) in platelet function and arterial thrombosis in mice. Blood J. Am. Soc. Hematol. 2022, 139, 2050–2065. [Google Scholar] [CrossRef]
- Holbrook, L.M.; Sandhar, G.K.; Sasikumar, P.; Schenk, M.P.; Stainer, A.R.; Sahli, K.A.; Flora, G.D.; Bicknell, A.B.; Gibbins, J.M. A humanized monoclonal antibody that inhibits platelet-surface ERp72 reveals a role for ERp72 in thrombosis. J. Thromb. Haemost. 2018, 16, 367–377. [Google Scholar] [CrossRef]
- Yang, M.; Kennedy, Q.; Patel, A.; Chen, D.-Y.; Scartelli, C.; Osataphan, S.; Bekendam, R.; Xie, H.; Lin, L.; Schmaier, A.A.; et al. Galloylated Polyphenols Represent a New Class of Antithrombotic Antagonists of Thiol Isomerases. Blood 2023, 142, 684. [Google Scholar] [CrossRef]
- Lay, A.J.; Dupuy, A.; Hagimola, L.; Tieng, J.; Larance, M.; Zhang, Y.; Yang, J.; Kong, Y.; Chiu, J.; Gray, E.; et al. Endoplasmic reticulum protein 5 attenuates platelet endoplasmic reticulum stress and secretion in a mouse model. Blood Adv. 2023, 7, 1650–1665. [Google Scholar] [CrossRef]
- Wang, T.-E.; Lai, Y.-H.; Yang, K.-C.; Lin, S.-J.; Chen, C.-L.; Tsai, P.-S. Counteracting cisplatin-induced testicular damages by natural polyphenol constituent honokiol. Antioxidants 2020, 9, 723. [Google Scholar] [CrossRef]
- Pienkowski, V.M.; Kucharczyk, M.; Młynek, M.; Szczałuba, K.; Rydzanicz, M.; Poszewiecka, B.; Skórka, A.; Sykulski, M.; Biernacka, A.; Koppolu, A.A. Mapping of breakpoints in balanced chromosomal translocations by shallow whole-genome sequencing points to EFNA5, BAHD1 and PPP2R5E as novel candidates for genes causing human Mendelian disorders. J. Med. Genet. 2019, 56, 104–112. [Google Scholar] [CrossRef]
- Min, Z.; Ye, Z.; Gang, L.; Boyu, D.; Xueyan, X. IFI27 as a potential indicator for severe Enterovirus 71-infected hand foot and mouth disease. Virus Res. 2020, 289, 198149. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.l.; Li, B.; Cheng, Y.j.; Li, M.; Yang, Z.d. Upregulation of cGMP-dependent Protein Kinase (PRKG1) in the Development of Adolescent Idiopathic Scoliosis. Orthop. Surg. 2020, 12, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Özcan, S.; Alessio, N.; Acar, M.B.; Mert, E.; Omerli, F.; Peluso, G.; Galderisi, U. Unbiased analysis of senescence associated secretory phenotype (SASP) to identify common components following different genotoxic stresses. Aging 2016, 8, 1316. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Peyper, J.M.; Garnett, S.; Tadokera, R.; Wilkinson, R.; Meintjes, G.; Blackburn, J.M. TB-IRIS: Proteomic analysis of in vitro PBMC responses to Mycobacterium tuberculosis and response modulation by dexamethasone. Exp. Mol. Pathol. 2017, 102, 237–246. [Google Scholar] [CrossRef]
- Couasnay, G.; Bon, N.; Devignes, C.S.; Sourice, S.; Bianchi, A.; Véziers, J.; Weiss, P.; Elefteriou, F.; Provot, S.; Guicheux, J. PiT1/Slc20a1 is required for endoplasmic reticulum homeostasis, chondrocyte survival, and skeletal development. J. Bone Miner. Res. 2019, 34, 387–398. [Google Scholar] [CrossRef] [PubMed]
- DiChiara, A.S.; Taylor, R.J.; Wong, M.Y.; Doan, N.-D.; Rosario, A.M.D.; Shoulders, M.D. Mapping and exploring the collagen-I proteostasis network. ACS Chem. Biol. 2016, 11, 1408–1421. [Google Scholar] [CrossRef]
- Kojima, R.; Paslawski, W.; Lyu, G.; Arenas, E.; Zhang, X.; Svenningsson, P. Secretome Analyses Identify FKBP4 as a GBA1-Associated Protein in CSF and iPS Cells from Parkinson’s Disease Patients with GBA1 Mutations. Int. J. Mol. Sci. 2024, 25, 683. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, L.; Yao, C.; Wang, T.; Wu, J.; Shang, Y.; Li, B.; Xia, H.; Huang, S.; Wang, F.; et al. Early plasma proteomic biomarkers and prediction model of acute respiratory distress syndrome after cardiopulmonary bypass: A prospective nested cohort study. Int. J. Surg. 2023, 109, 2561–2573. [Google Scholar] [CrossRef] [PubMed]
- Lambert, N.; Kengne-Ouafo, J.A.; Rissy, W.M.; Diane, U.; Murithi, K.; Kimani, P.; Awe, O.I.; Dillman, A. Transcriptional Profiles Analysis of COVID-19 and Malaria Patients Reveals Potential Biomarkers in Children. BioRxiv 2022. [Google Scholar] [CrossRef]
- Sekelova, Z.; Polansky, O.; Stepanova, H.; Fedr, R.; Faldynova, M.; Rychlik, I.; Vlasatikova, L. Different roles of CD4, CD8 and γδ T-lymphocytes in naive and vaccinated chickens during Salmonella enteritidis infection. Proteomics 2017, 17, 1700073. [Google Scholar] [CrossRef]
- Warren, W.C.; Rice, E.S.; Meyer, A.; Hearn, C.J.; Steep, A.; Hunt, H.D.; Monson, M.S.; Lamont, S.J.; Cheng, H.H. The immune cell landscape and response of Marek’s disease resistant and susceptible chickens infected with Marek’s disease virus. Sci. Rep. 2023, 13, 5355. [Google Scholar] [CrossRef]
- Liyanage, D.; Omeka, W.; Sandamalika, W.G.; Udayantha, H.; Jeong, T.; Lee, S.; Lee, J. PDI family thioredoxin from disk abalone (Haliotis discus discus): Responses to stimulants (PAMPs, bacteria, and viral) and functional characterization. Fish Shellfish Immunol. 2022, 120, 261–270. [Google Scholar] [CrossRef]
- Huang, D.; Liu, A.Y.; Leung, K.-S.; Tang, N.L. Direct measurement of b lymphocyte gene expression biomarkers in peripheral blood transcriptomics enables early prediction of vaccine seroconversion. Genes 2021, 12, 971. [Google Scholar] [CrossRef]
- Zeng, Y.; Ma, W.; Ren, X.; Wang, Y. Inhibition of TXNDC5 attenuates lipopolysaccharide-induced septic shock by altering inflammatory responses. Lab. Investig. 2022, 102, 422–431. [Google Scholar] [CrossRef]
- Kaya, B.; Turhan, U.; Sezer, S.; Bestel, A.; Okumuş, Z.G.; Dağ, İ.; Erel, Ö. Maternal serum TXNDC5 levels and thiol/disulfide homeostasis in preeclamptic pregnancies. J. Matern.-Fetal Neonatal Med. 2020, 33, 671–676. [Google Scholar] [CrossRef]
- Bradley, S.J.; Suarez-Fueyo, A.; Moss, D.R.; Kyttaris, V.C.; Tsokos, G.C. T cell transcriptomes describe patient subtypes in systemic lupus erythematosus. PLoS ONE 2015, 10, e0141171. [Google Scholar] [CrossRef]
- Bae, H.; Lunetta, K.L.; Murabito, J.M.; Andersen, S.L.; Schupf, N.; Perls, T.; Sebastiani, P. Genetic associations with age of menopause in familial longevity. Menopause 2019, 26, 1204. [Google Scholar] [CrossRef] [PubMed]
- Hulleman, J.D.; Genereux, J.C.; Nguyen, A. Mapping wild-type and R345W fibulin-3 intracellular interactomes. Exp. Eye Res. 2016, 153, 165–169. [Google Scholar] [CrossRef]
- Barry, J.D.; Fagny, M.; Paulson, J.N.; Aerts, H.J.; Platig, J.; Quackenbush, J. Histopathological image QTL discovery of immune infiltration variants. Iscience 2018, 5, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Sabeena, M.; Jamil, K.; Swamy, A. Identification of histone deacetylase10 (HDAC10) protein interaction network and its implications on cancer. Int. J. Adv. Sci. Res. Manag. 2018, 3, 157–162. [Google Scholar]
- Chawsheen, H.A.; Ying, Q.; Jiang, H.; Wei, Q. A critical role of the thioredoxin domain containing protein 5 (TXNDC5) in redox homeostasis and cancer development. Genes Dis. 2018, 5, 312–322. [Google Scholar] [CrossRef]
- DeZwaan-McCabe, D.; Sheldon, R.D.; Gorecki, M.C.; Guo, D.-F.; Gansemer, E.R.; Kaufman, R.J.; Rahmouni, K.; Gillum, M.P.; Taylor, E.B.; Teesch, L.M.; et al. ER stress inhibits liver fatty acid oxidation while unmitigated stress leads to anorexia-induced lipolysis and both liver and kidney steatosis. Cell Rep. 2017, 19, 1794–1806. [Google Scholar] [CrossRef]
- Adamson, B.; Norman, T.M.; Jost, M.; Cho, M.Y.; Nuñez, J.K.; Chen, Y.; Villalta, J.E.; Gilbert, L.A.; Horlbeck, M.A.; Hein, M.Y.; et al. A multiplexed single-cell CRISPR screening platform enables systematic dissection of the unfolded protein response. Cell 2016, 167, 1867–1882.e21. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-O.; Jin, U.-H.; Kang, J.H.; Kim, S.B.; Guthrie, A.S.; Sreevalsan, S.; Lee, J.-S.; Safe, S. The orphan nuclear receptor NR4A1 (Nur77) regulates oxidative and endoplasmic reticulum stress in pancreatic cancer cells. Mol. Cancer Res. 2014, 12, 527–538. [Google Scholar] [CrossRef] [PubMed]


| Name | Symbol | Related Pathway | Localization |
|---|---|---|---|
| Thioredoxin domain containing 5 | TXNDC5 | Unfolded protein response | Endoplasmic reticulum |
| Neuritin 1 | NRN1 | Metabolism of proteins, Post-translational modification | Extracellular |
| Coagulation factor XIII a chain | F13A1 | Response to elevated platelet cytosolic Ca2+ | Extracellular |
| Ras responsive element binding protein 1 | RREB1 | Ras/Raf-mediated cell differentiation | Nucleus |
| Signal sequence receptor subunit 1 | SSR1 | Unfolded protein response | Endoplasmic reticulum |
| Rio kinase 1 | RIOK1 | rRNA processing in the nucleus and cytosol Processing of capped intron-containing pre-mRNA | Cytosol, nucleus |
| Desmoplakin | DSP | Signaling by Rho GTPases | Nucleus, plasma membrane |
| Bone morphogenetic protein 6 | BMP6 | Apoptotic pathways in synovial fibroblasts MIF-mediated glucocorticoid regulation | Extracellular |
| Biogenesis of lysosomal organelles complex 1 subunit 5 | BLOC1S5 | Biogenesis of lysosome-related organelles | Cytosol |
| Eukaryotic translation elongation factor 1 epsilon 1 | EEF1E1 | Peptide chain elongation tRNA aminoacylation | Cytosol, nucleus |
| Solute carrier family 35 member b3 | SLC35B3 | Biotransformation Glycosaminoglycan metabolism | Golgi apparatus |
| Cancer antigen 1 | CAGE1 | Retinoblastoma progression | Cytosol, nucleus |
| Lymphocyte antigen 86 | LY86 | MyD88 dependent cascade initiated on endosome | Extracellular |
| Hepatocellular carcinoma upregulated long non-coding RNA | HULC | Cancer progression | Ribosome |
| Classification | Species | Disease | References |
|---|---|---|---|
| Long non-coding RNAs | LncRNA-ENST00000556926 | Lung cancer | [34] |
| Circular RNAs | CircRNA_0078710 | Hepatocellular carcinoma | [2,35] |
| CircRNA-104718 | Hepatocellular carcinoma | [37] | |
| CircRNA_0000517 | Hepatocellular carcinoma | [2,36] | |
| MicroRNAs | miRNA-431-5p | Hepatocellular carcinoma | [2,35] |
| miRNA-1296-5p | Hepatocellular carcinoma | [2,36] | |
| miRNA-218-5p | Hepatocellular carcinoma | [37] | |
| miRNA-30c-5p | Hepatocellular carcinoma | [38] | |
| miRNA-30e-5p | Hepatocellular carcinoma | [38] | |
| miRNA-573 | Rheumatoid arthritis | [2,39,40] | |
| miRNA-200b | Prostate cancer | [2,42] | |
| miRNA-92a | Atherosclerosis | [43] | |
| miRNA-124 | Medulloblastoma | [41] | |
| MicroRNA offset RNAs | moRNA-21 | Breast cancer | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bidooki, S.H.; Navarro, M.A.; Fernandes, S.C.M.; Osada, J. Thioredoxin Domain Containing 5 (TXNDC5): Friend or Foe? Curr. Issues Mol. Biol. 2024, 46, 3134-3163. https://doi.org/10.3390/cimb46040197
Bidooki SH, Navarro MA, Fernandes SCM, Osada J. Thioredoxin Domain Containing 5 (TXNDC5): Friend or Foe? Current Issues in Molecular Biology. 2024; 46(4):3134-3163. https://doi.org/10.3390/cimb46040197
Chicago/Turabian StyleBidooki, Seyed Hesamoddin, María A. Navarro, Susana C. M. Fernandes, and Jesus Osada. 2024. "Thioredoxin Domain Containing 5 (TXNDC5): Friend or Foe?" Current Issues in Molecular Biology 46, no. 4: 3134-3163. https://doi.org/10.3390/cimb46040197
APA StyleBidooki, S. H., Navarro, M. A., Fernandes, S. C. M., & Osada, J. (2024). Thioredoxin Domain Containing 5 (TXNDC5): Friend or Foe? Current Issues in Molecular Biology, 46(4), 3134-3163. https://doi.org/10.3390/cimb46040197