Marine Compounds Selectively Induce Apoptosis in Female Reproductive Cancer Cells but Not in Primary-Derived Human Reproductive Granulosa Cells


Abstract
:1. Introduction
2. Results and Discussion
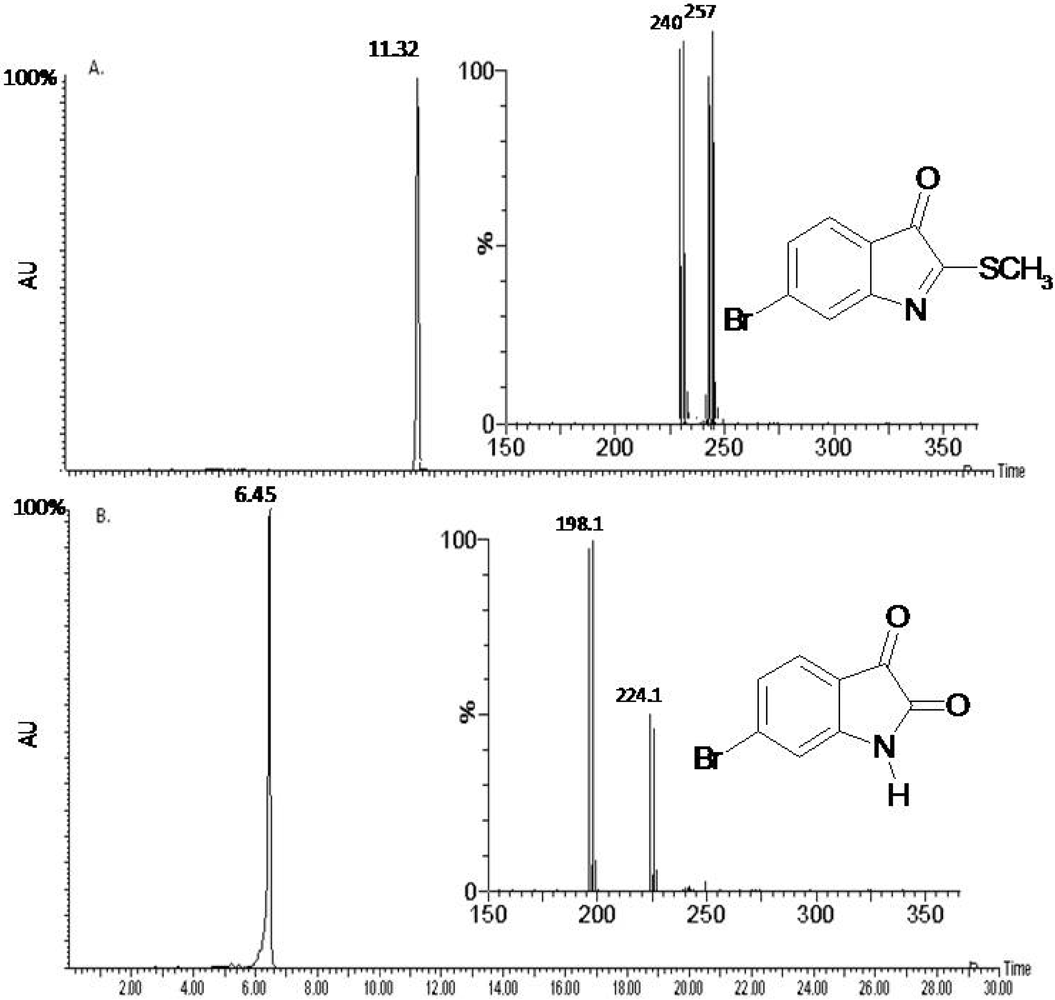
2.1. Compound Identification

2.2. Biological Activity of the D. orbita Compounds
2.2.1. Cell Viability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Time (h) | IC50 (µM) | |||
|---|---|---|---|---|---|
| Tyrindoleninone | 6-Bromoisatin | ||||
| CV | MTT | CV | MTT | ||
| HGC | 4 | >1953 | 3516 | >2232 | >4464 |
| KGN | 156 | 39 | 892 | 178 | |
| JAr | 234 | 39 | 223 | 223 | |
| OVCAR-3 | 313 | 156 | 446 | 402 | |
| HGC | 24 | >1953 | 1563 | >2232 | >4464 |
| KGN | 156 | 78 | 223 | 446 | |
| JAr | 195 | 117 | 178 | 446 | |
| OVCAR-3 | 352 | 234 | 268 | 446 | |
| HGC | 48 | n/t | 1563 | n/t | 1785 |
| KGN | n/t | 20 | n/t | 22 | |
| JAr | n/t | 78 | n/t | 446 | |
| OVCAR-3 | n/t | 31 | n/t | 89 | |
| HGC | 72 | n/t | 1563 | n/t | 2232 |
| KGN | n/t | <20 | n/t | <22 | |
| JAr | n/t | n/t | n/t | n/t | |
| OVCAR-3 | n/t | <20 | n/t | <22 | |
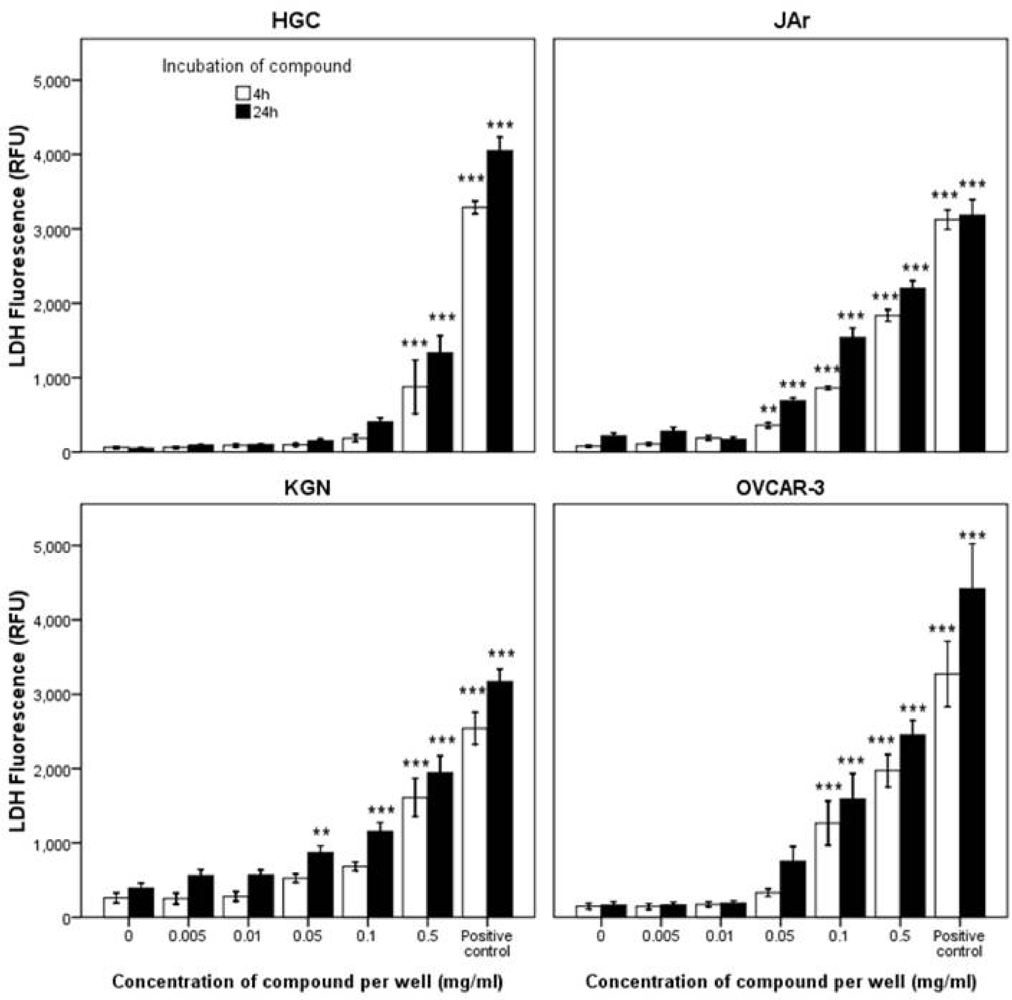
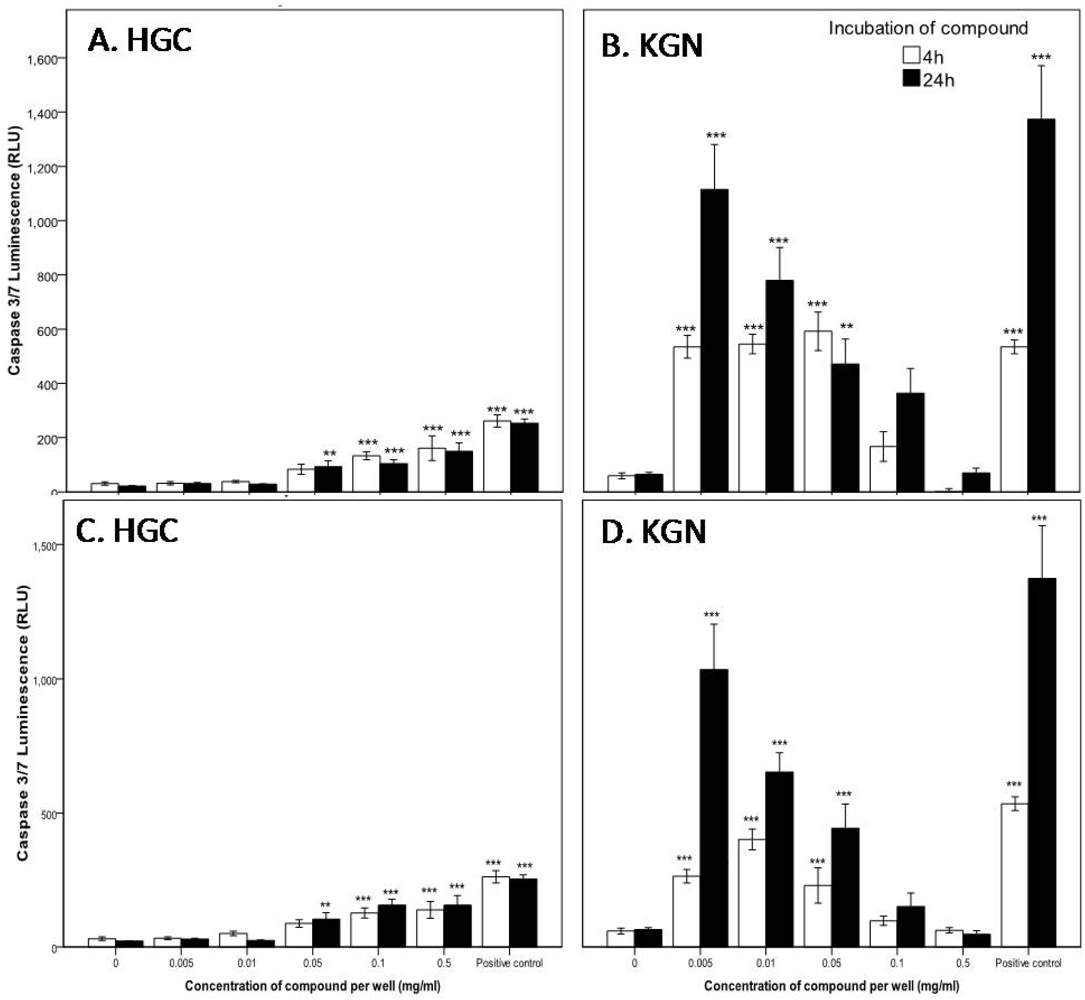
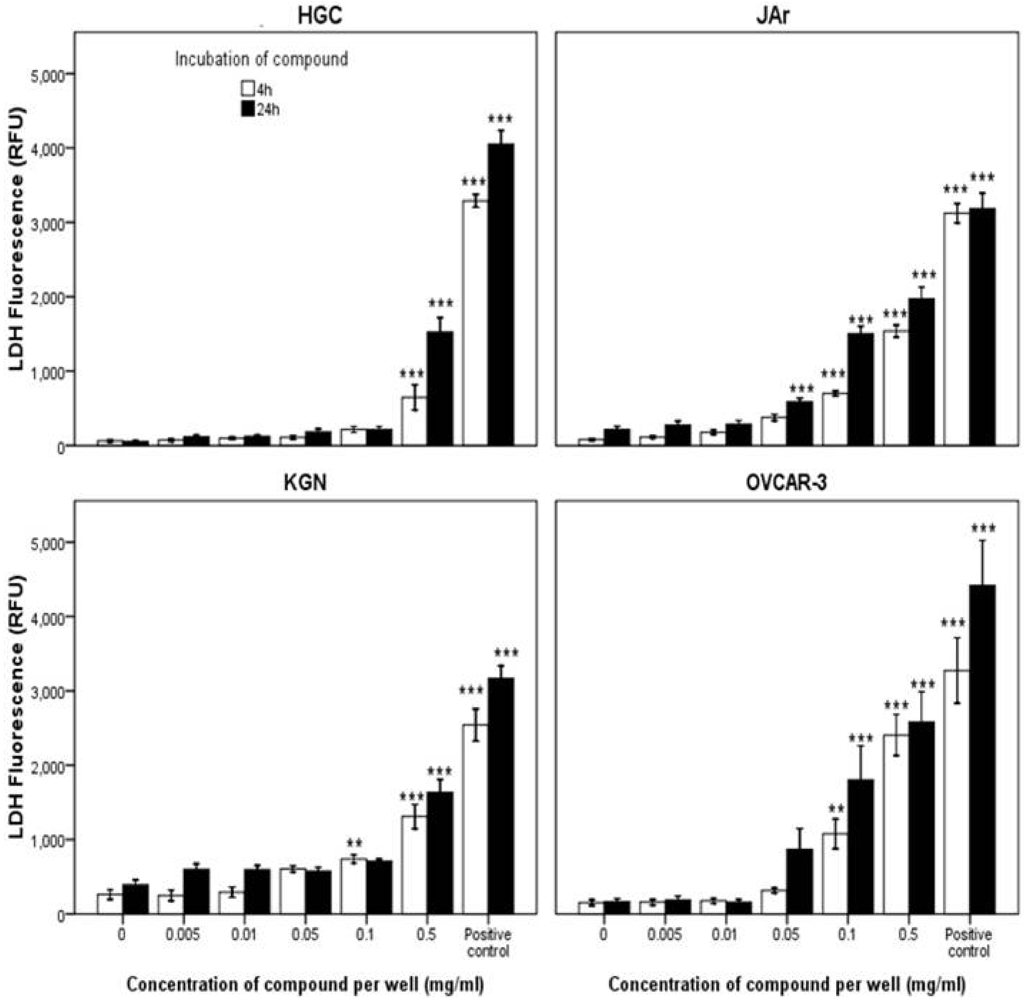
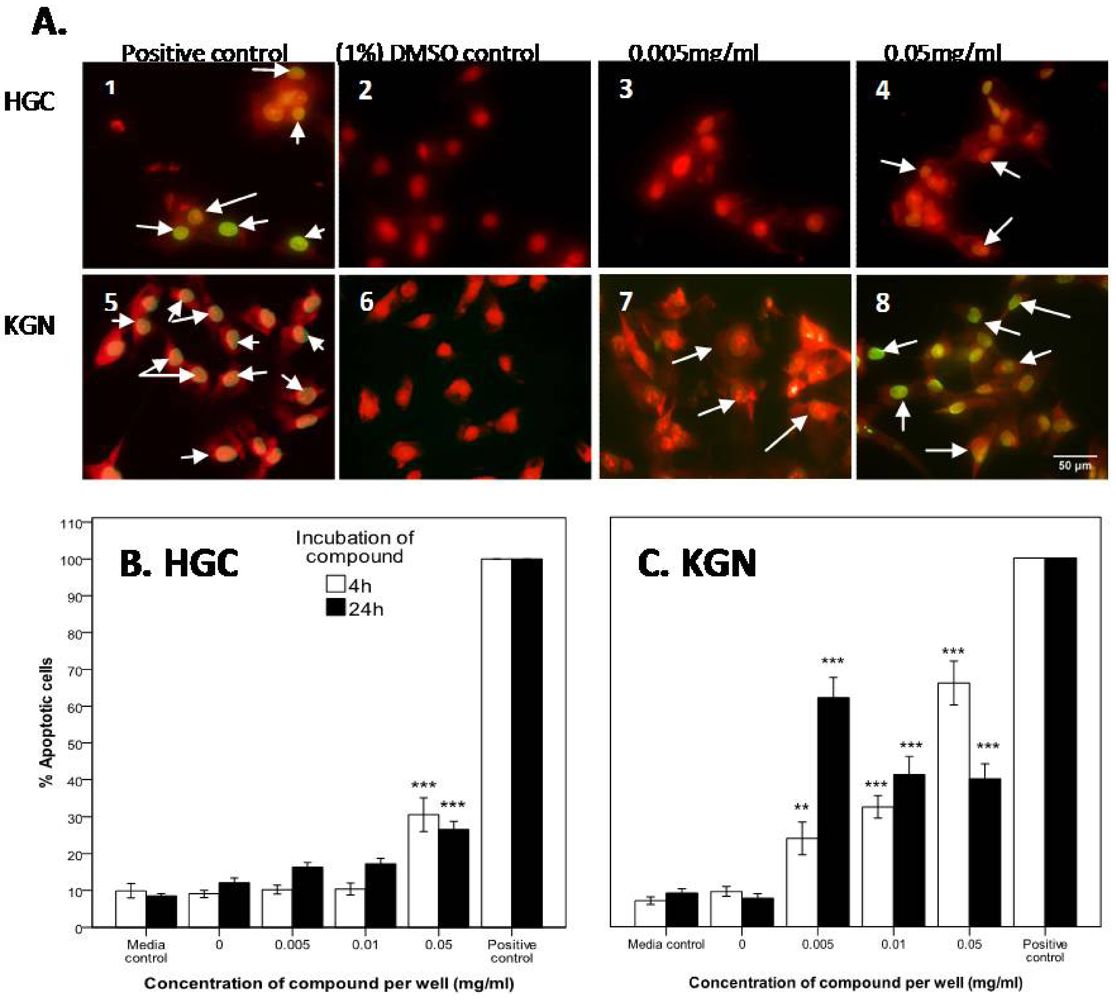
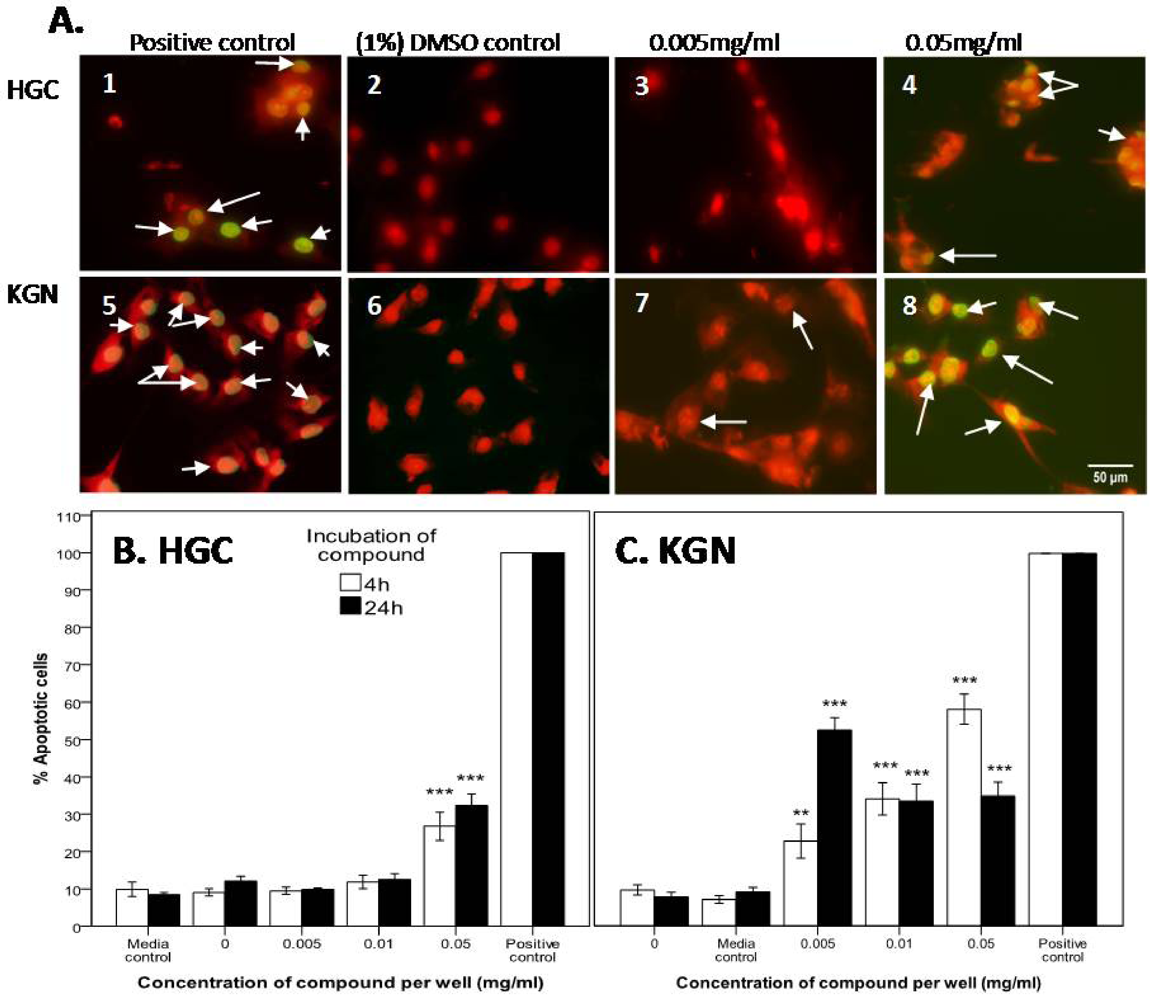
2.2.2. Mode of Action Investigation: Apoptosis and Necrosis Assays



2.2.3. Confirmation of Apoptosis by TUNEL Staining


3. Experimental Section
3.1. Extraction, Purification and Chemical Analysis
3.2. Cell Culture
3.2.1. Isolation of Primary-Derived Human Granulosa Cells
3.2.2. Cell Line Culture
3.3. Combined Caspase 3/7, Membrane Integrity and Cell Viability Assays
3.3.1. LDH Membrane Integrity and Caspase-Glo 3/7 Assay
3.3.2. Crystal Violet Cell Viability Assay
3.3.3. MTT Cell Viability Assay
3.4. Detection of Apoptotic KGN and Primary-Derived Granulosa Cells by TUNEL
3.5. Statistical Analysis
4. Conclusion
Acknowledgments
- Samples Availability: Available from the authors.
References
- Chen, Y.C.; Gudrun, P.; Wang, T.L.; Morin, P.J.; Risberg, B.; Kristensen, G.B.; Yu, A.; Davidson, B.; Shih, I.M. Apolipoprotein E is required for cell proliferation and survival in ovarian cancer. Cancer Res. 2005, 65, 331–337. [Google Scholar]
- Wray, N.; Markovic, M.; Manderson, L. Discourses of normality and difference: Responses to diagnosis and treatment of gynaecological cancer of Australian women. Soc. Sci. Med. 2007, 64, 2260–2271. [Google Scholar]
- Sapoznik, S.; Cohen, B.; Tzuman, Y.; Meir, G.; Ben-Dor, S.; Harmelin, A.; Neeman, M. Gonadotrophin-regulated lymphangiogenesis in ovarian cancer is mediated by LEDGF-induced Expression of VEGF-C. Cancer Res. 2009, 69, 9306–9314. [Google Scholar]
- Decruze, S.B.; Kirwan, J.M. Ovarian cancer. Curr. Obestetrics Gynaecol. 2006, 16, 161–167. [Google Scholar]
- World Health Organisation, Comprehensive Cervical Cancer Control a Guide to Essential Practice; World Health Organisation: New Delhi, India, 2006; pp. 1–282.
- Luo, H.; Daddysman, M.; Rankin, G.; Jiang, B.; Chen, Y. Kaempferol enhances cisplatin’s effect on ovarian cancer cells through promoting apoptosis caused by down regulation of cMyc. Cancer Cell Int. 2010, 10. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar]
- Chin, Y.; Balunas, M.J.; Chai, H.B.; Kinghorn, D. Drug discovery from natural sources. AAPS J. 2006, 8, E239–E253. [Google Scholar]
- Simmons, T.L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine natural products as anticancer drugs. Mol. Cancer Ther. 2005, 4, 333–342. [Google Scholar]
- Jimeno, J.; Faircloth, G.; Fernández Sousa-Faro, J.M.; Scheuer, P.; Rinehart, K. New marine derived anticancer therapeutics—A journey from the sea to clinical trials. Mar. Drugs 2004, 2, 14–29. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Hu, W.P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2009, 26, 170–244. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2010, 27, 165–237. [Google Scholar]
- Erwin, P.M.; Lopez-Legentil, S.; Schuhmann, P.W. The pharmaceutical value of marine biodiversity for anti-cancer drug discovery. Ecol. Econ. 2010, 70, 445–451. [Google Scholar]
- Mayer, A.M.; Gustafson, K.R. Marine pharmacology in 2005–2006: Antitumour and cytotoxic compounds. Eur. J. Cancer 2008, 44, 2357–2387. [Google Scholar]
- Gottesman, M.M. Mechanism of cancer drug resistance. Annu. Rev. Med. 2002, 56, 615–627. [Google Scholar]
- Sessa, C.; de Braud, F.; Perotti, A.; Bauer, J.; Curigliano, G.; Noberasco, C.; Zanaboni, F.; Gianni, L.; Marsoni, S.; Jimeno, J.; et al. Trabectedin for women with ovarian carcinoma after treatment with platinum and taxanes fails. J. Clin. Oncol. 2005, 23, 1867–1874. [Google Scholar] [PubMed]
- D’Incalci, M.; Galmarini, C.M. A review of trabectedin (ET-743): A unique mechanism of action. Mol. Cancer Ther. 2010, 9, 2157–2163. [Google Scholar]
- Carter, N.J.; Keam, S.J. Trabectedin: A review of its use in soft tissue sarcoma and ovarian cancer. Drugs 2010, 70, 335–376. [Google Scholar]
- Benkendorff, K. Molluscan biological and chemical diversity: Secondary metabolites and medicinal resources produced by marine molluscs. Biol. Rev. Camb. Philos. Soc. 2010, 85, 757–775. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Hu, W.P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2008, 25, 35–94. [Google Scholar]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tuinman, A.A.; Boettner, F.E.; Kizu, H.; Schmidt, J.M.; Baczynskyj, L.; Tomer, K.B.; Bontems, R.J. The isolation and structure of a remarkable marine animal antineoplastic constituent: Dolastatin 10. J. Am. Chem. Soc. 1987, 109, 6883–6885. [Google Scholar]
- Tran, H.T.; Newman, R.A.; Beck, D.E.; Huie, R.; Abruzzese, J.L.; Madden, T.A. A phase I, pharmocokinetic/pharmocodynamic study of dolastatin-10 in adult patients with advanced solid tumors. Proc. Am. Assoc. Cancer Res. 1997, 38, 306. [Google Scholar]
- Perez, E.; Hillman, D.; Fishkin, P.; Krook, J.; Tan, W.; Kuriakose, P.; Alberts, S.; Dakhil, S. Phase II trial of dolastatin-10 in patients with advanced breast cancer. Investig. New Drugs 2005, 23, 257–261. [Google Scholar]
- Hoffman, M.A.; Blessing, J.A.; Lentz, S.S. A phase II trial of dolastatin-10 in recurrent platinum-sensitive ovarian carcinoma: A gynecologic oncology group study. Gynecol. Oncol. 2003, 89, 95–98. [Google Scholar]
- Hamann, M.T.; Otto, C.S.; Scheuer, P.J. Kahalalides: Bioactive peptides from a marine mollusk elysia rufescens and its algal diet Bryopsis sp. J. Org. Chem. 1996, 61, 6594–6600. [Google Scholar]
- Janmaat, M.L.; Rodriguez, J.A.; Jimeno, J.; Kruyt, F.A.E.; Giaccone, G. Kahalalide F induces necrosis-like death that involves depletion of ErB3 and inhibition of Akt signaling. Mol. Pharmacol. 2005, 68, 502–510. [Google Scholar]
- Ling, Y.-H.; Aracil, M.; Zou, Y.; Yuan, Z.; Lu, B.; Jimeno, J.; Cuervo, A.M.; Perez-Soler, R. PM02734 (Elisidepsin) induces caspase-independent cell death associated with features of autophagy, inhibition of the Akt/mTOR signaling pathway, and activation of death-associated protein kinase. Clin.Cancer Res. 2011, 17, 5353–5366. [Google Scholar]
- Den Brok, M.W.; Nuijen, B.; Garcia, J.L.; Miranda, E.; Calvo, P.; Manada, C.; Beijnen, J.H. Compatibility and stability of the novel anticancer agent ES-285 × HCl formulated with 2-hydroxypropyl-beta-cyclodextrin in infusion devices. Pharmazie 2006, 61, 21–24. [Google Scholar]
- Benkendorff, K.; McIver, C.M.; Abbott, C.A. Bioactivity of the murex homeopathic remedy and of extracts from an australian muricid mollusc against human cancer cells. Evid. Based Complement. Altern. Med. 2011, 2011. [Google Scholar] [CrossRef]
- Vine, K.; Locke, J.; Ranson, M.; Benkendorff, K.; Pyne, S.; Bremner, J. In vitro cytotoxicity evaluation of some substituted isatin derivatives. Bioorg. Med. Chem. 2007, 15, 931–938. [Google Scholar]
- Westley, C.B.; McIver, C.M.; Abbott, C.A.; le Leu, R.K.; Benkendorff, K. Enhanced acute apoptotic response to azoxymethane-induced DNA damage in the rodent colonic epithelium by Tyrian purple purcursors: A potential colorectal cancer chemopreventative. Cancer Biol. Ther. 2010, 9, 34–42. [Google Scholar]
- Cazalet, S. Materia Medica (Reversed Kent’s Repertory). 2008. Available online: http://www.homeoint.org/hidb/kent/m/murx.htm (accessed on 7 April 2008).
- Dunham, C. Murex purpurea. Am. Homeopath. Rev. 1864, 4, 306–315. [Google Scholar]
- Hoessel, R.; Leclerc, S.; Endicot, J.A.; Nobel, M.E.M.; Lawrie, A.; Tunnah, P.; Leost, M.; Damiens, E.; Marie, D.; Marko, D.; et al. Indirubin, the active constituent of a Chinese antileukaemia medicine, inhibits cyclin-dependent kinases. Nat. Cell Biol. 1999, 1, 60–67. [Google Scholar]
- Kim, S.A.; Kim, Y.C.; Kim, S.W.; Lee, S.H.; Min, J.J.; Ahn, S.G.; Yoon, J.H. Antitumor activity of novel indirubin derivatives in rat tumor model. Clin. Cancer Res. 2007, 13, 253–259. [Google Scholar]
- Matesic, L.; Locke, J.M.; Bremner, J.B.; Pyne, S.G.; Skropeta, D.; Ranson, M.; Vine, K.L. N-phenethyl and N-naphthylmethyl isatins and analogues as in vitro cytotoxic agents. Bioorg. Med. Chem. 2008, 16, 3118–3124. [Google Scholar]
- Nishi, Y.; Yanase, T.; Mu, Y.; Oba, K.; Ichino, I.; Saito, M.; Nomura, M.; Mukasa, C.; Okabe, T.; Goto, K.; et al. Establishment and characterization of a steroidogenic human granulosa-like tumor cell line, KGN, that expresses functional follicle-stimulating hormone receptor. Endocrinology 2001, 142, 437–444. [Google Scholar]
- Hamilton, T.C.; Young, R.C.; McKoy, W.M.; Grotzinger, K.G.; Green, J.A.; Chu, E.W.; Whang-Peng, J.; Rogan, A.M.; Green, W.R.; Ozols, R.F. Characterization of a human ovarian carcinoma cell line (NIH:OVCAR-3) with androgen and estrogen receptors. Cancer Res. 1983, 43, 5379–5389. [Google Scholar]
- Pattillo, R.A.; Ruckert, A.; Hussa, R.; Bernstein, R. The jar cell line—Continuous human multihormone production and controls. Vitro Abstr. 1971, 6, 398–399. [Google Scholar]
- Benkendorff, K.; Bremner, J.B.; Davis, A.R. Tyrian purple precursors in the egg masses of the australian muricid, Dicathais orbita: A possible defence role. J. Chem. Ecol. 2000, 26, 1037–1050. [Google Scholar]
- Riss, T.L.; Moravec, R.A. Use of multiple assay endpoints to investigate the effects of incubation time, dose of toxin, and plating density in cell-based cytotoxicity assays. Assay Drug Dev. Technol. 2004, 2, 51–62. [Google Scholar]
- Jin, Z.; El-Deiry, W.S. Overview of cell death signaling pathways. Cancer Biol. Ther. 2005, 4, 139–163. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar]
- Vine, K.L.; Locke, J.M.; Ranson, M.; Pyne, S.G.; Bremner, J.B. An investigation into the cytotoxicity and mode of action of some novel N-Alkyl-substituted isatins. J. Med. Chem. 2007, 50, 5109–5117. [Google Scholar]
- Weng, J.R.; Tsai, C.H.; Kulp, S.K.; Wang, D.; Lin, C.H.; Yang, H.C.; Ma, Y.; Sargeant, A.; Chiu, C.F.; Tsai, M.H.; Chen, C.S. A potent indole-3-carbinol derived antitumor agent with pleiotropic effects on multiple signaling pathways in prostate cancer cells. Cancer Res. 2007, 67, 7815–7824. [Google Scholar]
- Cane, A.; Tournaire, M.C.; Barritault, D.; Crumeyrolle-Arias, M. The endogenous oxindoles 5-hydroxyoxindole and isatin are antiproliferative and proapoptotic. Biochem. Biophys. Res. Commun. 2000, 276, 379–384. [Google Scholar]
- Steinmetz, R.; Wagoner, H.A.; Zeng, P.; Hammone, J.R.; Hannon, T.S.; Meyers, J.L.; Pescovitz, O.H. Mechanisms regulating the constitutive activation of the extracellular signal-regulated kinase (ERK) signaling pathway in ovarian cancer and the effect of ribonucleic acid interference for ERK1/2 on cancer cell proliferation. Mol. Endocrinol. 2004, 18, 2570–2582. [Google Scholar]
- Woods, D.C.; Liu, H.; Nishi, Y.; Yanase, T.; Johnson, A.L. Inhibition of proteasome activity sensitizes human granulosa tumor cells to TRAIL-induced cell death. Cancer Lett. 2008, 260, 20–27. [Google Scholar]
- Manabe, N.; Goto, Y.; Matsuda-Minehata, F.; Inoue, N.; Maeda, A.; Sakamaki, K.; Miyano, T. Regulation mechanism of selective atresia in porcine follicles: Regulation of granulosa cell apoptosis during atresia. J. Reprod. Dev. 2004, 50, 493–514. [Google Scholar]
- Jääskeläinen, M.; Kyrönlahti, A.; Anttonen, M.; Nishi, Y.; Yanase, T.; Secchiero, P.; Zauli, G.; Tapanainen, J.S.; Heikinheimo, M.; Vaskivuo, T.E. TRAIL pathway components and their putative role in granulosa cell apoptosis in the human ovary. Differentiation 2009, 77, 369–376. [Google Scholar]
- Pozhilenkova, E.A.; Salmina, A.B.; Yamanova, M.V.; Ruksha, T.G.; Mikhutkina, S.V.; Trufanova, L.V. Disorders of folliculogenesis are associated with abnormal expression of peripheral benzodiazepine receptors in granulosa cells. Bull. Exp. Biol. Med. 2008, 145, 29–32. [Google Scholar]
- Young, F.M.; Menadue, M.F.; Lavranos, T.C. Effects of the insecticide amitraz, an α2-adrenergic receptor agonist, on human luteinized granulosa cells. Hum. Reprod. 2005, 20, 3018–3025. [Google Scholar]
- Freshney, R.I. Culture of Animal Cells: A Manual of Basic Technique; John Wiley & Sons: Hoboken, NJ, USA, 2005; pp. 346-348, 359-373. [Google Scholar]
- Siddiqui, E.J.; Shabbir, M.A.; Mikhailidis, D.P.; Mumtaz, F.H.; Thompson, C.S. The effect of serotonin and serotonin antagonists on bladder cancer cell proliferation. Br. J. Urol. Int. 2006, 97, 634–639. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and suvival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Pallant, J. SPSS Survival Manual a Step by Step Guide to Data Analysis Using SPSS, 2nd ed; Allen & Unwin: Sydney, Australia, 2002; pp. 229–237. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Edwards, V.; Benkendorff, K.; Young, F. Marine Compounds Selectively Induce Apoptosis in Female Reproductive Cancer Cells but Not in Primary-Derived Human Reproductive Granulosa Cells. Mar. Drugs 2012, 10, 64-83. https://doi.org/10.3390/md10010064
Edwards V, Benkendorff K, Young F. Marine Compounds Selectively Induce Apoptosis in Female Reproductive Cancer Cells but Not in Primary-Derived Human Reproductive Granulosa Cells. Marine Drugs. 2012; 10(1):64-83. https://doi.org/10.3390/md10010064
Chicago/Turabian StyleEdwards, Vicki, Kirsten Benkendorff, and Fiona Young. 2012. "Marine Compounds Selectively Induce Apoptosis in Female Reproductive Cancer Cells but Not in Primary-Derived Human Reproductive Granulosa Cells" Marine Drugs 10, no. 1: 64-83. https://doi.org/10.3390/md10010064
APA StyleEdwards, V., Benkendorff, K., & Young, F. (2012). Marine Compounds Selectively Induce Apoptosis in Female Reproductive Cancer Cells but Not in Primary-Derived Human Reproductive Granulosa Cells. Marine Drugs, 10(1), 64-83. https://doi.org/10.3390/md10010064