Antiviral Activities and Putative Identification of Compounds in Microbial Extracts from the Hawaiian Coastal Waters

 and
and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses | Viral Family | Enveloped | Structure |
|---|---|---|---|
| Herpes simplex virus type one (HSV-1) | Herpesviridae | Yes | Linear double-stranded DNA genome |
| Vesicular stomatitis virus (VSV) | Rhabdoviridae | Yes | Single-stranded negative-sense RNA genome |
| Vaccinia virus | Poxvirus | Yes | Linear double-stranded DNA genome |
| Polio virus type one (poliovirus-1) | Picornaviridae | No | Single-stranded positive-sense RNA genome |
2. Results and Discussion
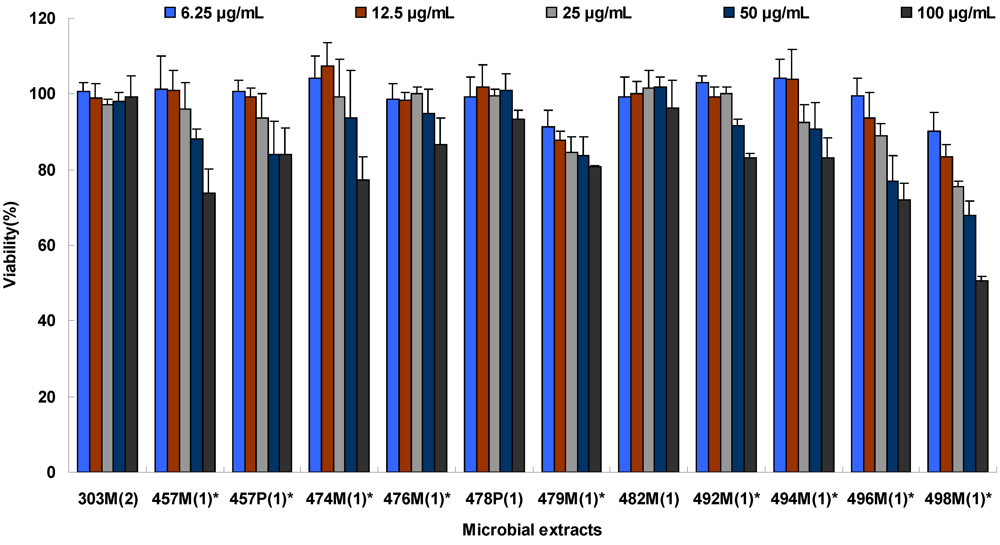
2.1. Extract Cytotoxicity

| Extracts b | Sources | IC10 (μg/mL) |
|---|---|---|
| 62M(1) | Oscillatoria sp.(Waikiki Aquarium) | 64.3 ± 5.7 |
| 62P(1) | Oscillatoria sp.(Waikiki Aquarium) | 49.2 ± 0.3 |
| 219P(3) | Pseudoalteromonas sp. from Mycale armata,Kaneohe Bay | ≤5.0 |
| 226P(3) | Pseudoalteromonas sp. from Mycale armata,Kaneohe Bay | 13.5 ± 1.1 |
| 258P(1) | Oscillatoria sp. from Fresh water | ≤5.0 |
| 258P(1) | Oscillatoria sp. from Fresh water | ≤5.0 |
| 298M(1) | bacterium from Kaneohe Bay, yet to be taxonomically classified | 42.5 ± 1.5 |
| 298P(1) | bacterium from Kaneohe Bay; yet to be taxonomically classified | 17.33 ± 0.17 |
| 303M(2) | bacterium from Kaneohe Bay; yet to be taxonomically classified | ≥100 |
| 331P(3) | Achromobacter fischeri from Mycale armata, Kaneohe Bay | ≤5.0 |
| 397M(1) | bacterium from Kaneohe Bay; yet to be taxonomically classified | ≥100 |
| 397P(1) | bacterium from Kaneohe Bay; yet to be taxonomically classified | 6.5 ± 0.4 |
| 435P(1) | bacterium from Hawaii Ocean Time Series (HOTS) site; yet to be taxonomically classified | ≤5.0 |
| 456P(1) | Marine bacterium from HOTS site; yet to be taxonomically classified | 9.1 ± 0.2 |
| 457M(1) | Diatom, Amphora sp., from Great Salt Lake | 44.6 ± 1.9 |
| 457P(1) | Diatom, Amphora sp., from Great Salt Lake | 51.7 ± 2.3 |
| 460M(1) | bacterium from HOTS site; yet to be taxonomically classified | 22.5 ± 3.2 |
| 464P(1) | bacterium from HOTS site; yet to be taxonomically classified | ≤5.0 |
| 474M(1) | bacterium; yet to be taxonomically classified | 63.1 ± 4.2 |
| 475M(1) | bacterium; yet to be taxonomically classified | ≥100 |
| 476M(1) | bacterium; yet to be taxonomically classified | 48.9 ± 3.0 |
| 477P(1) | bacterium; yet to be taxonomically classified | ≥100 |
| 478M(1) | bacterium; yet to be taxonomically classified | ≤5.0 |
| 478P(1) | bacterium; yet to be taxonomically classified | ≥100 |
| 479M(1) | bacterium; yet to be taxonomically classified | 7.4 ± 0.5 |
| 482M(1) | bacterium; yet to be taxonomically classified | ≥100 |
| 483P(1) | bacterium; yet to be taxonomically classified | ≥100 |
| 485M(1) | bacterium; yet to be taxonomically classified | 68.9 ± 5.8 |
| 485P(1) | bacterium; yet to be taxonomically classified | 54.8 ± 4.7 |
| 492M(1) | bacterium; yet to be taxonomically classified | 61.0 ± 3.2 |
| 492P(1) | bacterium; yet to be taxonomically classified | indissoluble |
| 494M(1) | bacterium; yet to be taxonomically classified | 44.0 ± 2.8 |
| 495M(1) | bacterium; yet to be taxonomically classified | ≥100 |
| 495P(1) | bacterium; yet to be taxonomically classified | 24.7 ± 1.8 |
| 496M(1) | bacterium; yet to be taxonomically classified | 16.2 ± 1.3 |
| 496P(1) | bacterium; yet to be taxonomically classified | ≤5.0 |
| 497P(1) | bacterium; yet to be taxonomically classified | ≤5.0 |
| 498M(1) | bacterium; yet to be taxonomically classified | 6.6 ± 0.6 |
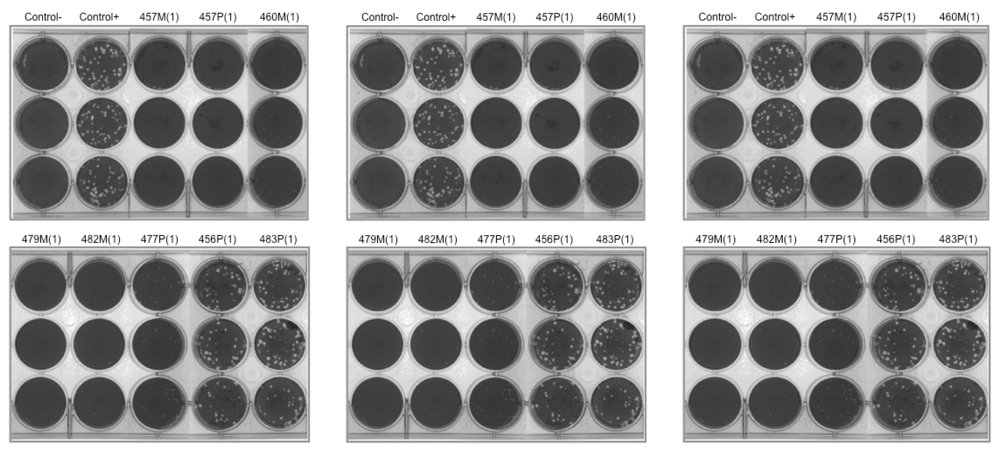
2.2. Viral Inhibition by Cell Pretreatment
2.3. Viral Inhibition by Virus Pretreatment
| Extracts | Concentrations (μg/mL) | HSV-1 | VSV | Vaccinia | Poliovirus-1 |
|---|---|---|---|---|---|
| 62M(1) | 50 | - | - | - | - |
| 62P(1) | 50 | - | +++ | - | - |
| 226P(3) | 12.5 | ++ | - | - | - |
| 298M(1) | 50 | - | +++ | - | - |
| 298P(1) | 12.5 | - | ++ | - | - |
| 303M(2) | 100 | + | +++ | + | - |
| 397M(1) | 100 | - | - | - | - |
| 397P(1) | 6.25 | - | - | - | - |
| 456P(1) | 12.5 | - | - | - | - |
| 457M(1) | 25.0 | +++ | +++ | +++ | - |
| 457P(1) | 25.0 | +++ | +++ | +++ | - |
| 460M(1) | 25.0 | ++ | ++ | - | - |
| 474M(1) | 50.0 | + | +++ | ++ | - |
| 475M(1) | 100 | ++ | ++ | + | - |
| 476M(1) | 50.0 | ++ | +++ | ++ | - |
| 477P(1) | 100 | + | + | ++ | - |
| 478P(1) | 100 | +++ | + | +++ | - |
| 479M(1) | 6.25 | + | +++ | +++ | - |
| 482M(1) | 100 | +++ | +++ | +++ | - |
| 483P(1) | 100 | - | - | - | - |
| 485M(1) | 50.0 | ++ | ++ | ++ | - |
| 485P(1) | 50.0 | ++ | - | ++ | - |
| 492M(1) | 50.0 | +++ | ++ | ++ | - |
| 494M(1) | 50.0 | +++ | ++ | ++ | - |
| 495M(1) | 100 | - | - | - | - |
| 495P(1) | 25.0 | - | - | - | - |
| 496M(1) | 12.5 | ++ | +++ | ++ | - |
| 498M(1) | 6.25 | + | +++ | ++ | - |

2.4. Viral Replication Inhibition Assay

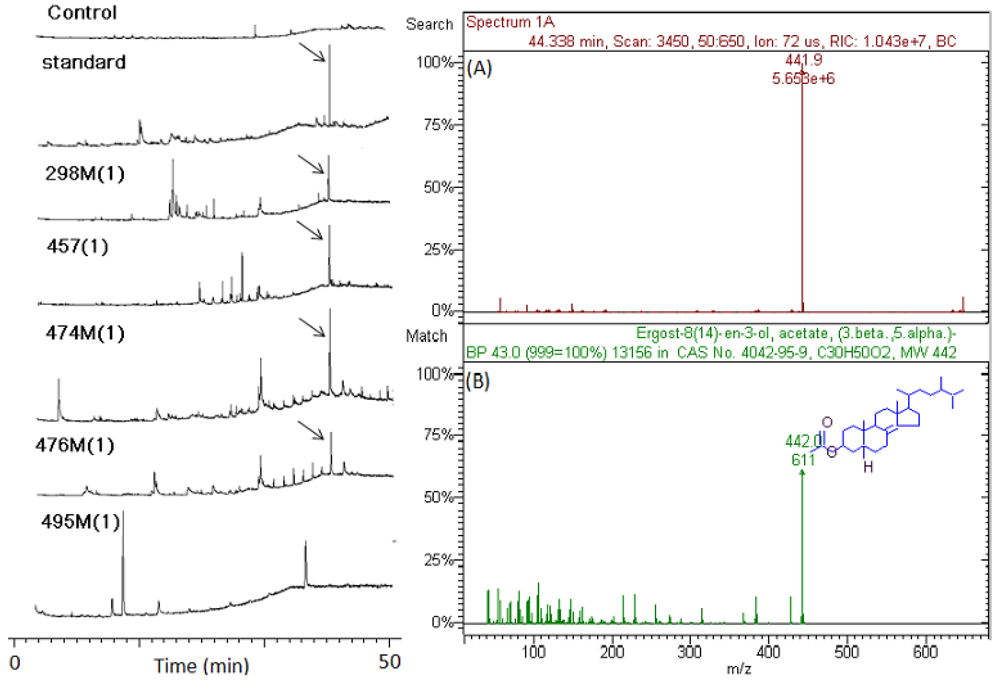
2.5. Isolation and Identification of Antiviral Components in Crude Extracts

2.6. Discussion
3. Experimental Section
3.1. Cell Culture and Media
3.2. Virus Stock Preparation
3.3. Marine Microbial Isolates
3.4. Marine Microbial Extracts
3.5. Cytotoxicity Assay by Cell Viability
3.6. Viral Plaque Reduction Assay
3.7. Viral Adsorption Assay–Cell Pretreatment
3.8. Viral Adsorption Assay–Virus Pretreatment
3.9. Viral Replication Inhibition Assay
3.10. Marine Microbial Extract Clean-Up and Fractionation
3.11. GC-ITMS Analysis
3.12. Data Analysis
4. Conclusions
Acknowledgements
References
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar]
- Ausubel, J.H.; Crist, D.T.; Waggoner, P.E. First Census of Marine Life 2010: Highlights of a Decade of Discovery; Census of Marine Life: Washington, DC, USA, 2010; p. 3. [Google Scholar]
- Donia, M.; Hamann, M.T. Marine natural products and their potential applications as anti-infective agents. Lancet Infect. Dis. 2003, 3, 338–348. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2002, 19, 1–48. [Google Scholar]
- Fautin, D.; Dalton, P.; Incze, L.S.; Leong, J.A.C.; Pautzke, C.; Rosenberg, A.; Sandifer, P.; Sedberry, G.; Tunnell, J.W.; Abbott, I.; et al. An overview of marine biodiversity in United States waters. PLoS One 2010, 5, e11914. [Google Scholar]
- Tissot, B.N.; Walsh, W.J.; Hixon, M.A. Hawaiian islands marine ecosystem case study: ecosystem- and community-based management in Hawaii. Coast. Manag. 2009, 37, 255–273. [Google Scholar]
- Hill, R.T.; Fenical, W. Pharmaceuticals from marine natural products: Surge or ebb? Curr. Opin. Biotechnol. 2010, 21, 777–779. [Google Scholar] [CrossRef]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar]
- SPSS Statistics Software, version 16.0, SPSS Inc.: Chicago, IL, USA, 2007.
- Sica, D.; Musumeci, D. Secosteroids of marine origin. Steroids 2004, 69, 743–756. [Google Scholar]
- Mayer, A.M.; Gustafson, K.R. Marine pharmacology in 2005-6: Antitumour and cytotoxic compounds. Eur. J. Cancer 2008, 44, 2357–2387. [Google Scholar]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2005–6: marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta 2009, 1790, 283–308. [Google Scholar] [CrossRef]
- Verkman, A.S. Drug discovery in academia. Am. J. Phys. Cell Phys. 2004, 286, C465–C474. [Google Scholar]
- Toranzo, A.E.; Barja, J.L.; Hetrick, F.M. Antiviral activity of antibiotic-producing marine bacteria. Can. J. Microbiol. 1982, 28, 231–238. [Google Scholar]
- Denis, F.; Brisou, J.F.; Dupuis, T. Inactivation of enteroviruses in seawater. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D 1975, 281, 471–474. [Google Scholar]
- Magnusson, S.; Gundersen, K.; Brandberg, A.; Lycke, E. Marine bacteria and their possible relation to the virus inactivation capacity of sea water. Acta Pathol. Microbiol. Scand. 1967, 71, 274–280. [Google Scholar]
- Gundersen, K.; Brandberg, A.; Magnusson, S.; Lycke, E. Characterization of a marine bacterium associated with virus inactivating capacity. Acta Pathol. Microbiol. Scand. 1967, 71, 281–286. [Google Scholar]
- Yasuhara, B.J.; Lu, Y.A. Marine compounds and their antiviral activities. Antiviral Res. 2010, 86, 231–240. [Google Scholar]
- Yasuhara-Bell, J.; Yang, Y.; Barlow, R.; Trapido-Rosenthal, H.; Lu, Y. In vitro evaluation of marine-microorganism extracts for anti-viral activity. Virol. J. 2010, 7, 182. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar] [CrossRef]
- Prestegard, S.K.; Oftedal, L.; Coyne, R.T.; Nygaard, G.; Skjaerven, K.H.; Knutsen, G.; Doskeland, S.O.; Herfindal, L. Marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity. Mar. Drugs 2009, 7, 605–623. [Google Scholar]
- Ianora, A.; Poulet, S.A.; Miralto, A. The effects of diatoms on copepod reproduction: A review. Phycologia 2003, 42, 351–363. [Google Scholar]
- Lebeau, T.; Robert, J.M. Diatom cultivation and biotechnologically relevant products. Part I: Cultivation at various scales. Appl. Microbiol. Biotechnol. 2003, 60, 612–623. [Google Scholar]
- Da Poian, A.T.; Gomes, A.M.O.; Coelho-Sampaio, T. Kinetics of intracellular viral disassembly and processing probed by Bodipy fluorescence dequenching. J. Virol. Methods 1998, 70, 45–58. [Google Scholar]
- Enwall, E.L.; van der Helm, D.; Hsu, I.N.; Pattabhiraman, T.; Scmitz, F.J.; Spraggins, R.L.; Weinheimer, A.J. Crystal structure and absolute configuration of two cyclopropane containing marine steroids. J. Chem. Soc., Chem. Commun. 1972, 4, 215–216. [Google Scholar]
- Lu, Y.N.; Aguirre, A.A.; Wang, Y.; Zeng, L.B.; Loh, P.C.; Yanagihara, R. Viral susceptibility of newly established cell lines from the Hawaiian monk seal Monachus schauinslandi. Dis. Aquat. Organ. 2003, 57, 183–191. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 16, 55–63. [Google Scholar]
- Lu, Y.N.; Loh, P.C. Some biological properties of a rhabdovirus isolated from penaeid shrimps. Arch. Virol. 1992, 127, 339–343. [Google Scholar]
- Sample Availability: Available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tong, J.; Trapido-Rosenthal, H.; Wang, J.; Wang, Y.; Li, Q.X.; Lu, Y. Antiviral Activities and Putative Identification of Compounds in Microbial Extracts from the Hawaiian Coastal Waters. Mar. Drugs 2012, 10, 521-538. https://doi.org/10.3390/md10030521
Tong J, Trapido-Rosenthal H, Wang J, Wang Y, Li QX, Lu Y. Antiviral Activities and Putative Identification of Compounds in Microbial Extracts from the Hawaiian Coastal Waters. Marine Drugs. 2012; 10(3):521-538. https://doi.org/10.3390/md10030521
Chicago/Turabian StyleTong, Jing, Hank Trapido-Rosenthal, Jun Wang, Youwei Wang, Qing X. Li, and Yuanan Lu. 2012. "Antiviral Activities and Putative Identification of Compounds in Microbial Extracts from the Hawaiian Coastal Waters" Marine Drugs 10, no. 3: 521-538. https://doi.org/10.3390/md10030521
APA StyleTong, J., Trapido-Rosenthal, H., Wang, J., Wang, Y., Li, Q. X., & Lu, Y. (2012). Antiviral Activities and Putative Identification of Compounds in Microbial Extracts from the Hawaiian Coastal Waters. Marine Drugs, 10(3), 521-538. https://doi.org/10.3390/md10030521