Inhibitory Effect of Dihydroaustrasulfone Alcohol on the Migration of Human Non-Small Cell Lung Carcinoma A549 Cells and the Antitumor Effect on a Lewis Lung Carcinoma-Bearing Tumor Model in C57BL/6J Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
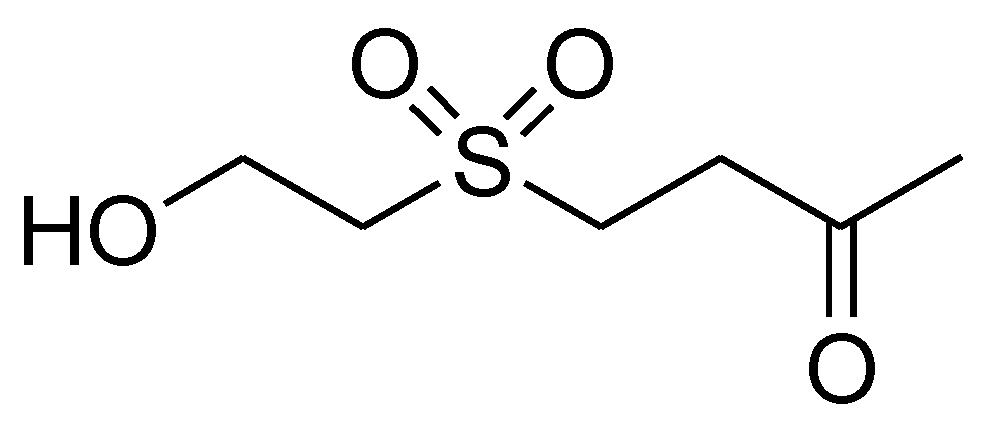
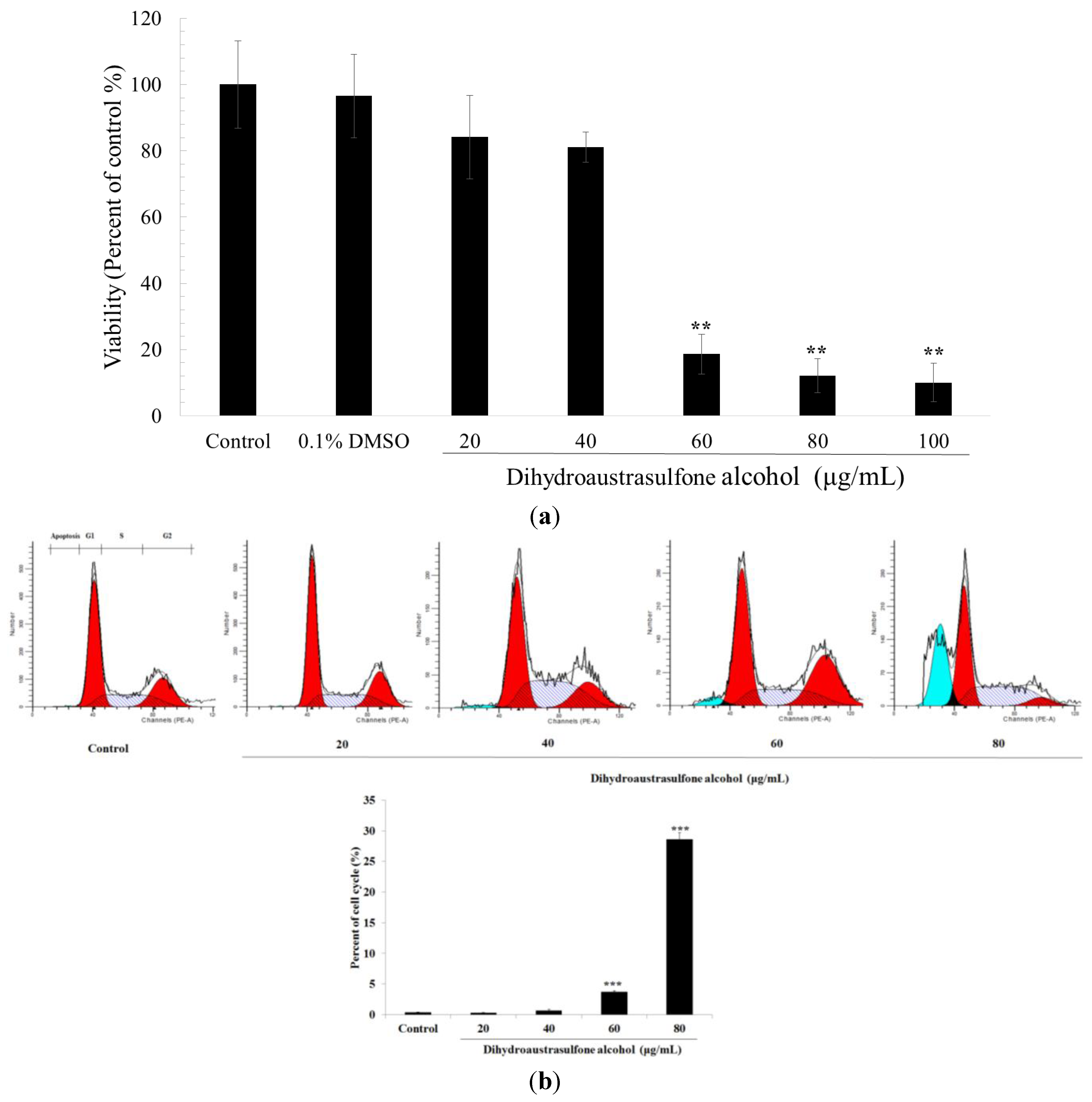
2.1. Cytotoxicity of Dihydroaustrasulfone Alcohol in A549 Cells

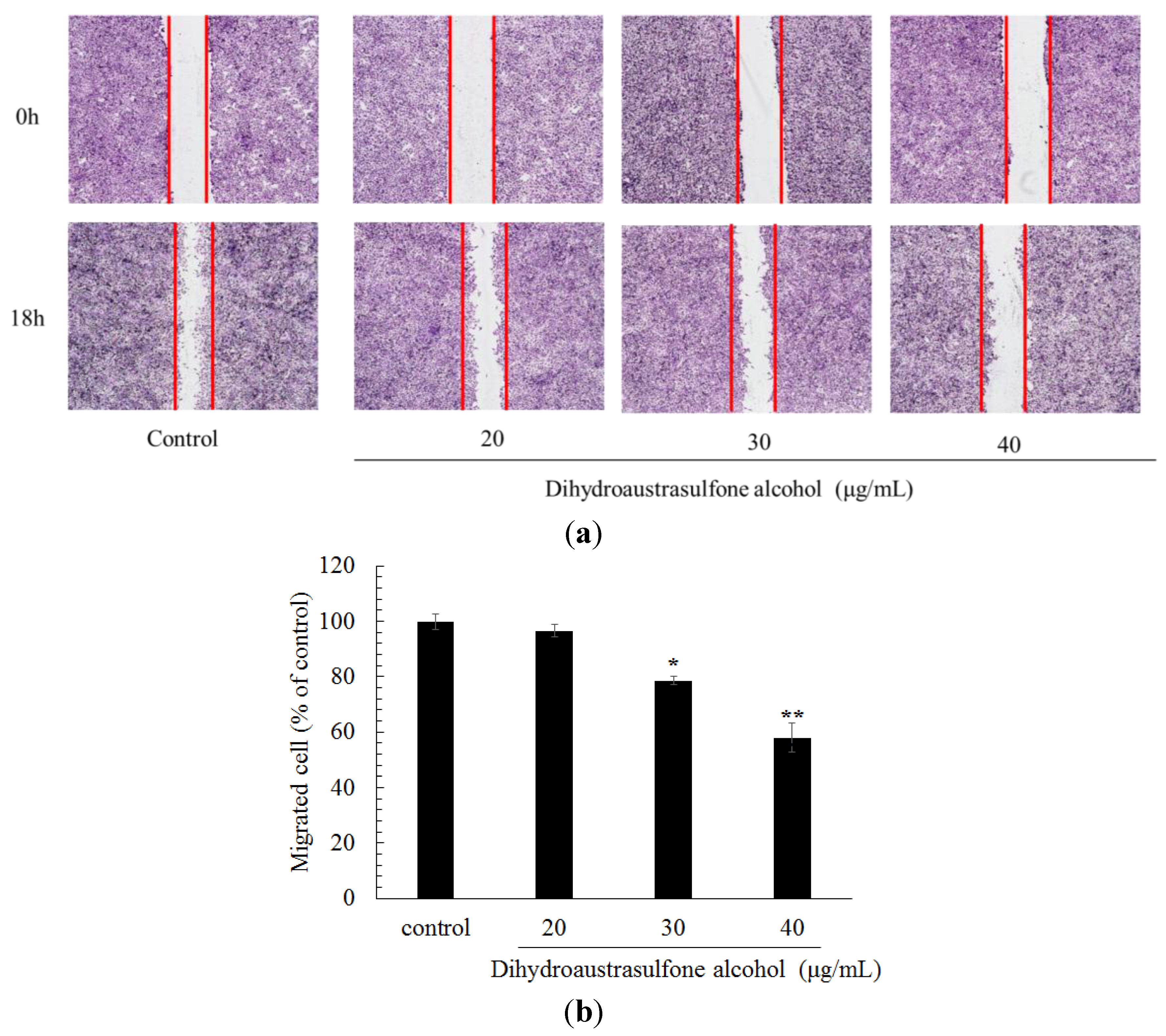
2.2. Effect of Dihydroaustrasulfone Alcohol on a Wound-Healing Assay in A549 Cells

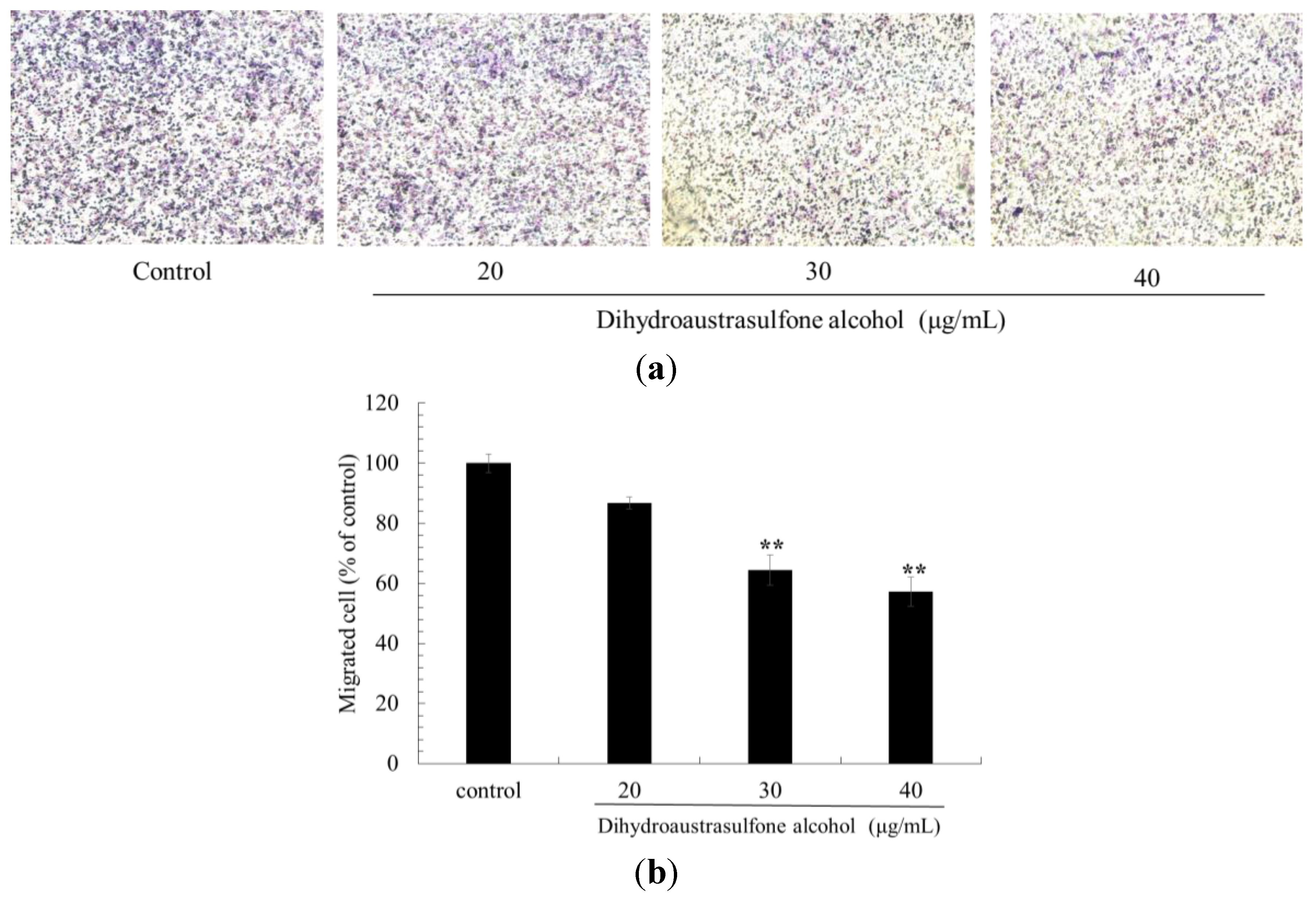
2.3. Effect of Dihydroaustrasulfone Alcohol in a Trans-well Assay with A549 Cells

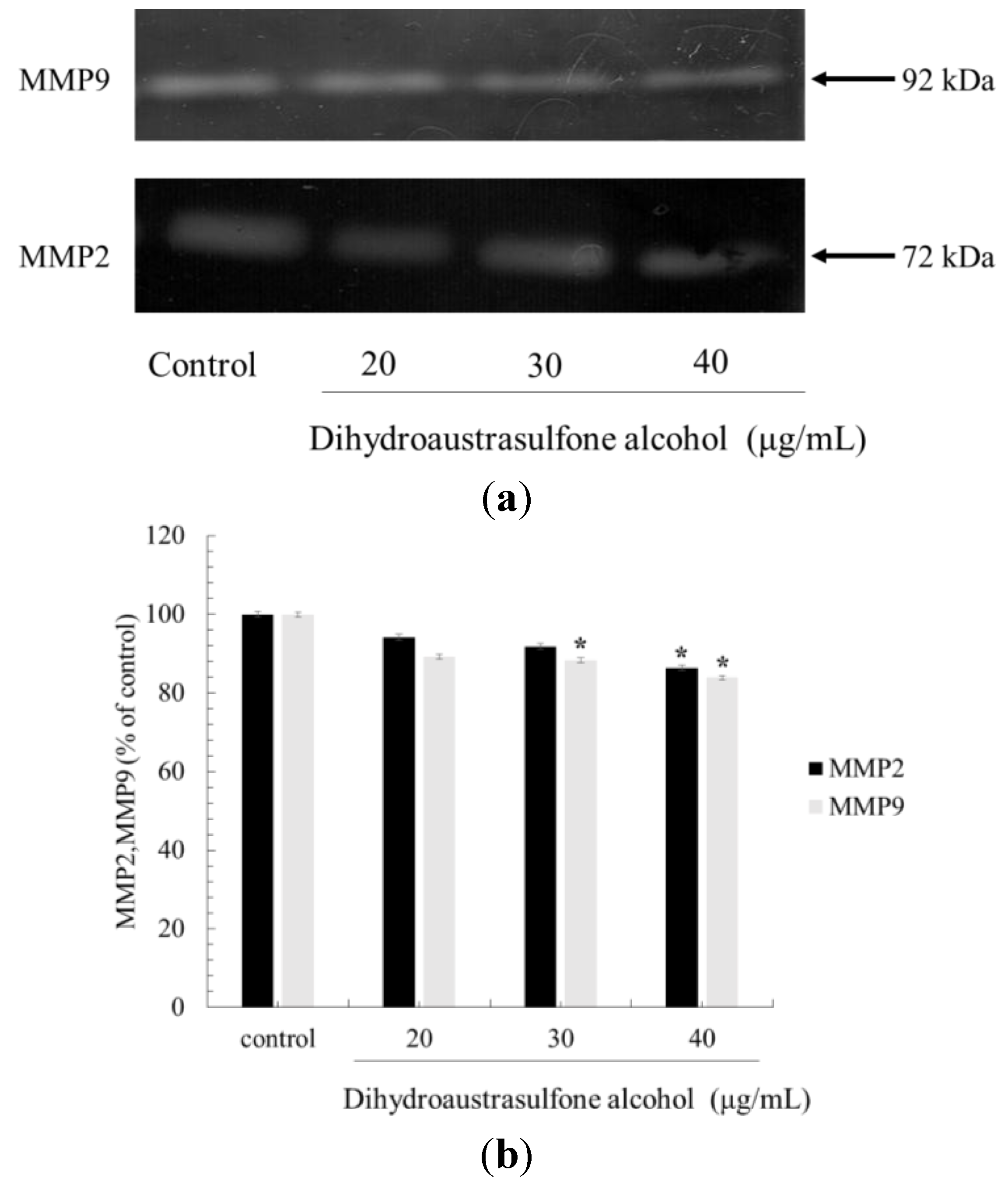
2.4. Effect of Dihydroaustrasulfone Aalcohol Inhibits the Release of MMP in A549 Cells

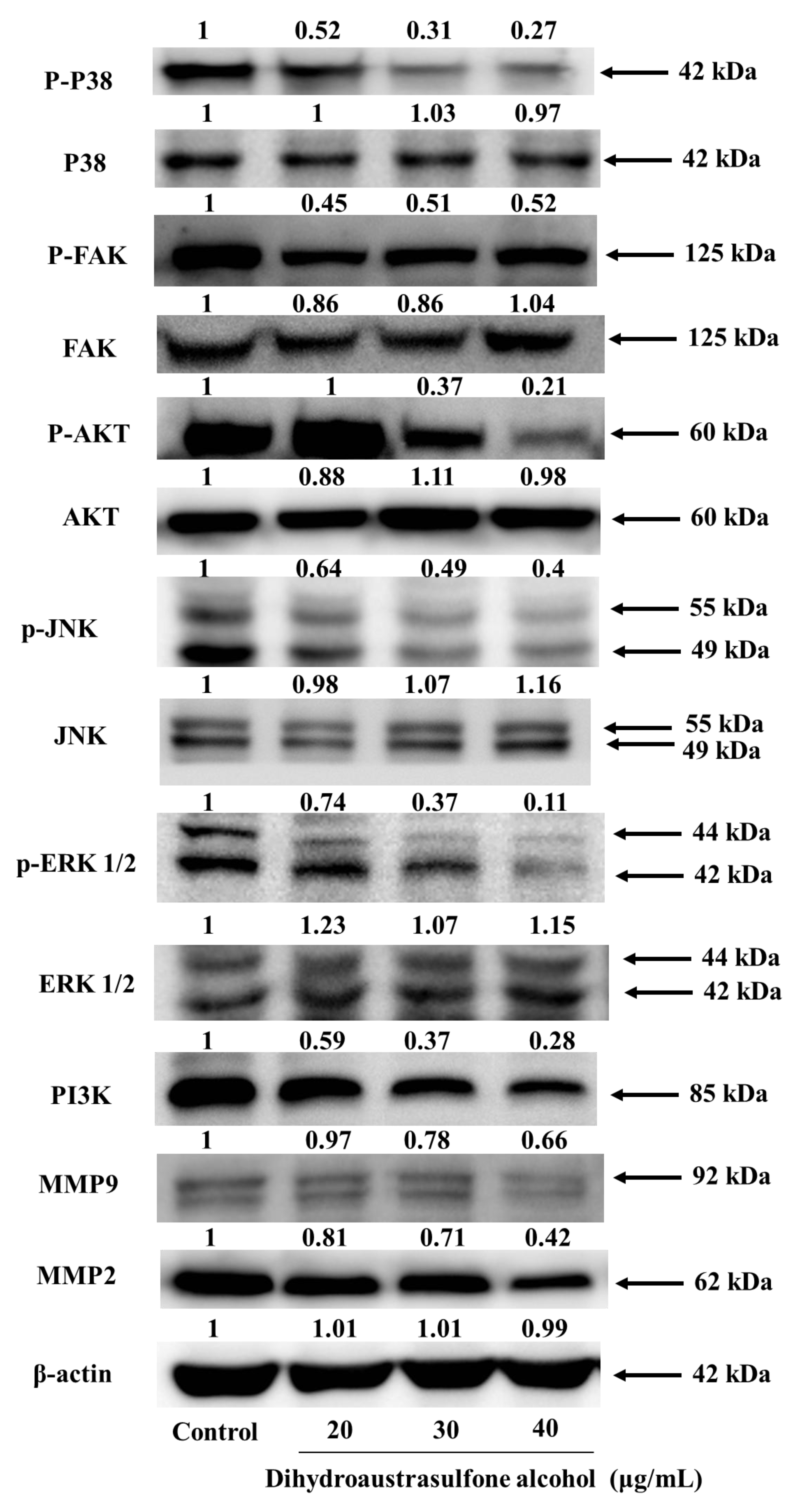
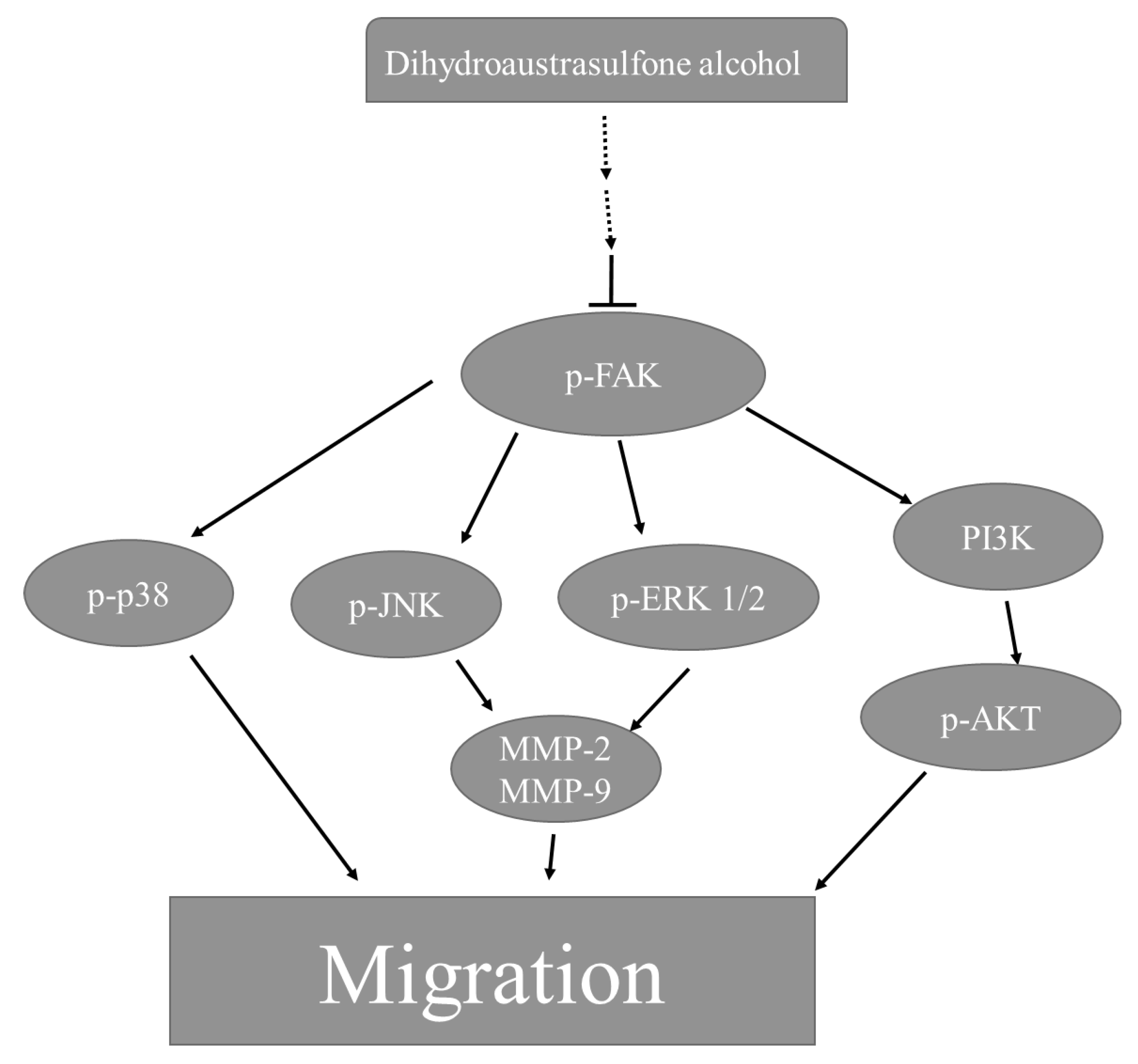
2.5. Dihydroaustrasulfone Alcohol Inhibits Migration-Related Proteins and PI3K/AKT Signaling in A549 Cells


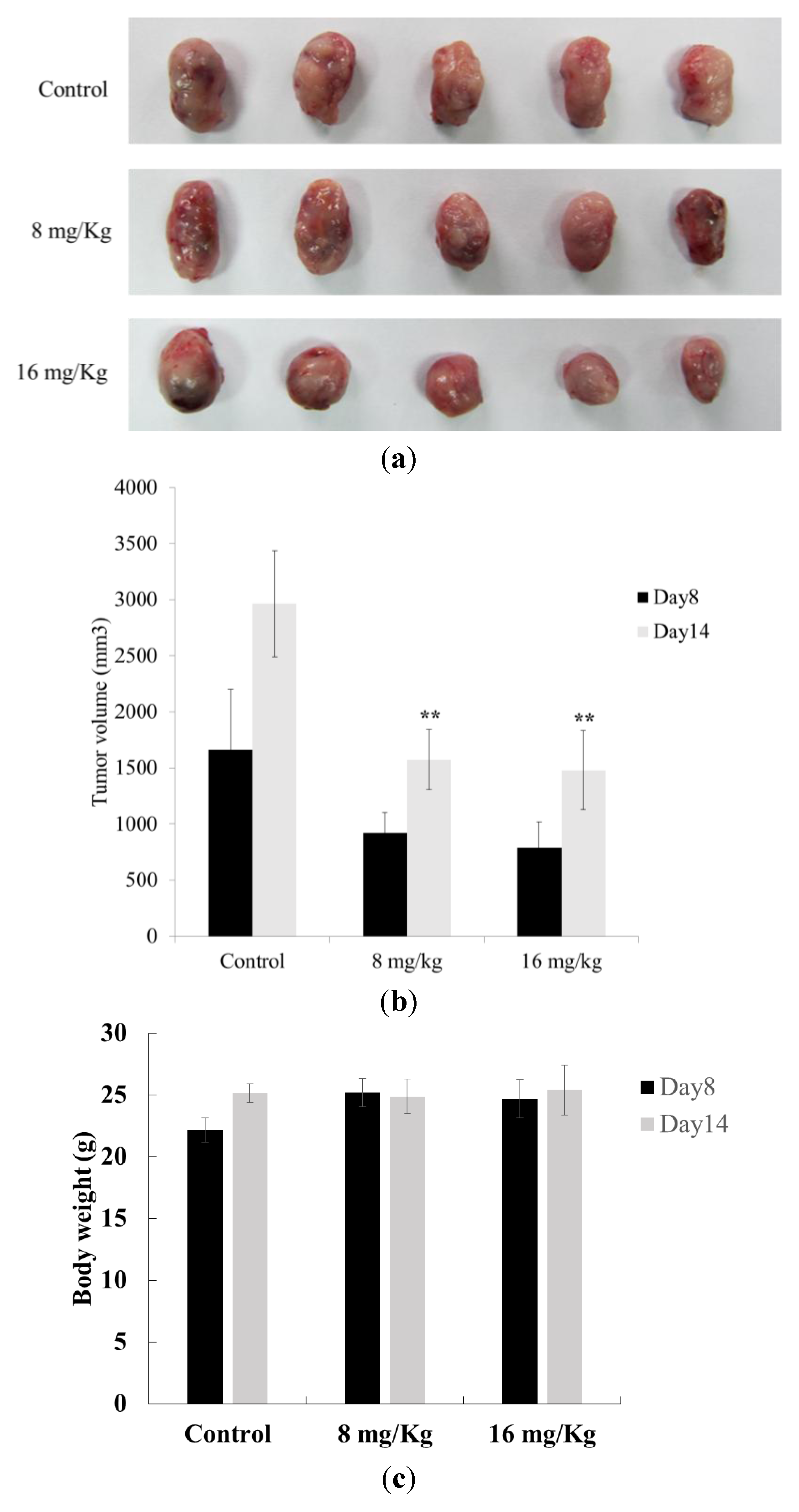
2.6. Antitumor Effect of Dihydroaustrasulfone Alcohol on a Lewis Lung Carcinoma (LLC)-Bearing Tumor Model in C57BL/6J Mice
3. Experimental Section
3.1. Materials

3.2. Cell Lines and Cultures
3.3. Cell Proliferation Assay
3.4. Flow Cytometry Analysis
3.5. Wound Healing Assay
3.6. Cell Migration Assay
3.7. Determination of MMP-2 and MMP-9 by Zymography
3.8. Western Blotting Analysis
3.9. Lewis Lung Cancer Cell Bearing Tumor Model of C57BL/6J Mice
3.10. Statistical Analysis
4. Conclusions

Acknowledgments
Conflicts of Interest
References
- Saba, N.F.; Khuri, F.R. Chemoprevention strategies for patients with lung cancer in the context of screening. Clin. Lung Cancer 2005, 7, 92–99. [Google Scholar] [CrossRef]
- Li, C.M.; Narayanan, R.; Lu, Y.; Hurh, E.; Coss, C.C.; Barrett, C.M.; Miller, D.D.; Dalton, J.T. 2-Arylthiazolidine-4-carboxylic acid amides (ATCAA) target dual pathways in cancer cells: 5′-AMP-activated protein kinase (AMPK)/mTOR and PI3K/Akt/mTOR pathways. Int. J. Oncol. 2010, 37, 1023–1030. [Google Scholar]
- Man, S.; Gao, W.; Zhang, Y.; Liu, Z.; Yan, L.; Huang, L.; Liu, C. Formosanin C-inhibited pulmonary metastasis through repression of matrix metalloproteinases on mouse lung adenocarcinoma. Cancer Biol. Ther. 2011, 11, 592–598. [Google Scholar] [CrossRef]
- Lirdprapamongkol, K.; Kramb, J.P.; Suthiphongchai, T.; Surarit, R.; Srisomsap, C.; Dannhardt, G.; Svasti, J. Vanillin suppresses metastatic potential of human cancer cells through PI3K inhibition and decreases angiogenesis in vivo. J. Agric. Food Chem. 2009, 57, 3055–3063. [Google Scholar] [CrossRef]
- Kodate, M.; Kasai, T.; Hashimoto, H.; Yasumoto, K.; Iwata, Y.; Manabe, H. Expression of matrix metalloproteinase (gelatinase) in T1 adenocarcinoma of the lung. Pathol. Int. 1997, 47, 461–469. [Google Scholar] [CrossRef]
- Weng, C.J.; Wu, C.F.; Huang, H.W.; Wu, C.H.; Ho, C.T.; Yen, G.C. Evaluation of anti-invasion effect of resveratrol and related methoxy analogues on human hepatocarcinoma cells. J. Agric. Food Chem. 2010, 58, 2886–2894. [Google Scholar] [CrossRef]
- Campbell, I.G.; Russell, S.E.; Choong, D.Y.; Montgomery, K.G.; Ciavarella, M.L.; Hooi, C.S.; Cristiano, B.E.; Pearson, R.B.; Phillips, W.A. Mutation of the PIK3CA gene in ovarian and breast cancer. Cancer Res. 2004, 64, 7678–7681. [Google Scholar] [CrossRef]
- Gupta, A.K.; Soto, D.E.; Feldman, M.D.; Goldsmith, J.D.; Mick, R.; Hahn, S.M.; Machtay, M.; Muschel, R.J.; McKenna, W.G. Signaling pathways in NSCLC as a predictor of outcome and response to therapy. Lung 2004, 182, 151–162. [Google Scholar]
- Shih, Y.W.; Chen, P.S.; Wu, C.H.; Jeng, Y.F.; Wang, C.J. Alpha-chaconine-reduced metastasis involves a PI3K/Akt signaling pathway with downregulation of NF-kappaB in human lung adenocarcinoma A549 cells. J. Agric. Food Chem. 2007, 55, 11035–11043. [Google Scholar]
- Jean, Y.H.; Chen, W.F.; Duh, C.Y.; Huang, S.Y.; Hsu, C.H.; Lin, C.S.; Sung, C.S.; Chen, I.M.; Wen, Z.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory and analgesic effects of the natural marine compound lemnalol from Formosan soft coral Lemnalia cervicorni. Eur. J. Pharmacol. 2008, 578, 323–331. [Google Scholar] [CrossRef]
- Jean, Y.H.; Chen, W.F.; Sung, C.S.; Duh, C.Y.; Huang, S.Y.; Lin, C.S.; Tai, M.H.; Tzeng, S.F.; Wen, Z.H. Capnellene, a natural marine compound derived from soft coral, attenuates chronic constriction injury-induced neuropathic pain in rats. Br. J. Pharmacol. 2009, 158, 713–725. [Google Scholar] [CrossRef]
- Tseng, Y.J.; Wen, Z.H.; Dai, C.F.; Chiang, M.Y.; Sheu, J.H. Nanolobatolide, a new C18 metabolite from the Formosan soft coral Sinularia nanolobata. Org. Lett. 2009, 11, 5030–5032. [Google Scholar] [CrossRef]
- Chao, C.H.; Wen, Z.H.; Wu, Y.C.; Yeh, H.C.; Sheu, J.H. Cytotoxic and anti-inflammatory cembranoids from the soft coral Lobophytum crassum. J. Nat. Prod. 2008, 71, 1819–1824. [Google Scholar] [CrossRef]
- Gallet, P.; Phulpin, B.; Merlin, J.L.; Leroux, A.; Bravetti, P.; Mecellem, H.; Tran, N.; Dolivet, G. Long-term alterations of cytokines and growth factors expression in irradiated tissues and relation with histological severity scoring. PLoS One 2011, 6, e29399. [Google Scholar]
- Greenfield, J.R.; McCormack, A.I. Medication to prevent breast cancer—too much to swallow? Med. J. Aust. 2012, 196, 646–649. [Google Scholar] [CrossRef]
- Rodemann, H.P.; Blaese, M.A. Responses of normal cells to ionizing radiation. Semin. Radiat. Oncol. 2007, 17, 81–88. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Wen, Z.H.; Chao, C.H.; Wu, M.H.; Sheu, J.H. A neuroprotective sulfone of marine origin and the in vivo anti-inflammatory activity of an analogue. Eur. J. Med. Chem. 2010, 45, 5998–6004. [Google Scholar] [CrossRef]
- Tsai, A.C.; Pai, H.C.; Wang, C.Y.; Liou, J.P.; Teng, C.M.; Wang, J.C.; Pan, S.L. In vitro and in vivo antitumor effects of MPT0B014, a novel derivative aroylquinoline, and in combination with erlotinib in human non-small-cell lung cancer cells. Br. J. Pharmacol. 2013, 171, 122–133. [Google Scholar]
- Wyrebska, A.; Gach, K.; Lewandowska, U.; Szewczyk, K.; Hrabec, E.; Modranka, J.; Jakubowski, R.; Janecki, T.; Szymanski, J.; Janecka, A. Anticancer Activity of New Synthetic alpha-Methylene-delta-Lactones on Two Breast Cancer Cell Lines. Basic Clin. Pharmacol. Toxicol. 2013, in press. [Google Scholar]
- Chung, T.W.; Choi, H.J.; Lee, J.Y.; Jeong, H.S.; Kim, C.H.; Joo, M.; Choi, J.Y.; Han, C.W.; Kim, S.Y.; Choi, J.S.; et al. Marine algal fucoxanthin inhibits the metastatic potential of cancer cells. Biochem. Biophys. Res. Commun. 2013, 439, 580–585. [Google Scholar] [CrossRef]
- Luo, Q.; Li, X.; Gao, Y.; Long, Y.; Chen, L.; Huang, Y.; Fang, L. MiRNA-497 regulates cell growth and invasion by targeting cyclin E1 in breast cancer. Cancer Cell Int. 2013, 13, 95:1–95:8. [Google Scholar]
- Liu, X.H.; Liu, Z.L.; Sun, M.; Liu, J.; Wang, Z.X.; De, W. The long non-coding RNA HOTAIR indicates a poor prognosis and promotes metastasis in non-small cell lung cancer. BMC Cancer 2013, 13, 464:1–464:10. [Google Scholar]
- Zhu, X.L.; Wang, Y.L.; Chen, J.P.; Duan, L.L.; Cong, P.F.; Qu, Y.C.; Li-Ling, J.; Zhang, M.X. Alternol inhibits migration and invasion of human hepatocellular carcinoma cells by targeting epithelial-to-mesenchymal transition. Tumour Biol. 2013, in press. [Google Scholar]
- Parsons, J.T. Focal adhesion kinase: The first ten years. J. Cell Sci. 2003, 116, 1409–1416. [Google Scholar] [CrossRef]
- Canel, M.; Secades, P.; Garzon-Arango, M.; Allonca, E.; Suarez, C.; Serrels, A.; Frame, M.; Brunton, V.; Chiara, M.D. Involvement of focal adhesion kinase in cellular invasion of head and neck squamous cell carcinomas via regulation of MMP-2 expression. Br. J. Cancer 2008, 98, 1274–1284. [Google Scholar] [CrossRef]
- Hwangbo, C.; Kim, J.; Lee, J.J.; Lee, J.H. Activation of the integrin effector kinase focal adhesion kinase in cancer cells is regulated by crosstalk between protein kinase Calpha and the PDZ adapter protein mda-9/Syntenin. Cancer Res. 2010, 70, 1645–1655. [Google Scholar] [CrossRef]
- Meng, X.N.; Jin, Y.; Yu, Y.; Bai, J.; Liu, G.Y.; Zhu, J.; Zhao, Y.Z.; Wang, Z.; Chen, F.; Lee, K.Y.; et al. Characterisation of fibronectin-mediated FAK signalling pathways in lung cancer cell migration and invasion. Br. J. Cancer 2009, 101, 327–334. [Google Scholar] [CrossRef]
- Lai, K.C.; Huang, A.C.; Hsu, S.C.; Kuo, C.L.; Yang, J.S.; Wu, S.H.; Chung, J.G. Benzyl isothiocyanate (BITC) inhibits migration and invasion of human colon cancer HT29 cells by inhibiting matrix metalloproteinase-2/-9 and urokinase plasminogen (uPA) through PKC and MAPK signaling pathway. J. Agric. Food Chem. 2010, 58, 2935–2942. [Google Scholar]
- Liao, H.F.; Chen, Y.Y.; Liu, J.J.; Hsu, M.L.; Shieh, H.J.; Liao, H.J.; Shieh, C.J.; Shiao, M.S.; Chen, Y.J. Inhibitory effect of caffeic acid phenethyl ester on angiogenesis, tumor invasion, and metastasis. J. Agric. Food Chem. 2003, 51, 7907–7012. [Google Scholar] [CrossRef]
- Kuo, P.T.; Lin, T.P.; Liu, L.C.; Huang, C.H.; Lin, J.K.; Kao, J.Y.; Way, T.D. Penta-O-galloyl-beta-d-glucose suppresses prostate cancer bone metastasis by transcriptionally repressing EGF-induced MMP-9 expression. J. Agric. Food Chem. 2009, 57, 3331–3339. [Google Scholar]
- Zhang, Y.; Thant, A.A.; Hiraiwa, Y.; Naito, Y.; Sein, T.T.; Sohara, Y.; Matsuda, S.; Hamaguchi, M. A role for focal adhesion kinase in hyluronan-dependent MMP-2 secretion in a human small-cell lung carcinoma cell line, QG90. Biochem. Biophys. Res. Commun. 2002, 290, 1123–1127. [Google Scholar] [CrossRef]
- Chang, Y.M.; Shih, Y.T.; Chen, Y.S.; Liu, C.L.; Fang, W.K.; Tsai, C.H.; Tsai, F.J.; Kuo, W.W.; Lai, T.Y.; Huang, C.Y. Schwann Cell Migration Induced by Earthworm Extract via Activation of PAs and MMP2/9 Mediated through ERK1/2 and p38. Evid Based Complement Alternat Med. 2011, 2011, 395–458. [Google Scholar]
- Kumar, B.; Koul, S.; Petersen, J.; Khandrika, L.; Hwa, J.S.; Meacham, R.B.; Wilson, S.; Koul, H.K. p38 mitogen-activated protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer by modulation of MMP-2 and MMP-9 activity. Cancer Res. 2010, 70, 832–841. [Google Scholar] [CrossRef]
- Loesch, M.; Zhi, H.Y.; Hou, S.W.; Qi, X.M.; Li, R.S.; Basir, Z.; Iftner, T.; Cuenda, A.; Chen, G. p38gamma MAPK cooperates with c-Jun in trans-activating matrix metalloproteinase 9. J. Biol. Chem. 2010, 285, 15149–15158. [Google Scholar]
- Chan, K.C.; Ho, H.H.; Huang, C.N.; Lin, M.C.; Chen, H.M.; Wang, C.J. Mulberry leaf extract inhibits vascular smooth muscle cell migration involving a block of small GTPase and Akt/NF-kappaB signals. J. Agric. Food Chem. 2009, 57, 9147–9153. [Google Scholar] [CrossRef]
- Zeng, Z.Z.; Jia, Y.; Hahn, N.J.; Markwart, S.M.; Rockwood, K.F.; Livant, D.L. Role of focal adhesion kinase and phosphatidylinositol 3′-kinase in integrin fibronectin receptor-mediated, matrix metalloproteinase-1-dependent invasion by metastatic prostate cancer cells. Cancer Res. 2006, 66, 8091–8099. [Google Scholar] [CrossRef]
- Choi, Y.A.; Lim, H.K.; Kim, J.R.; Lee, C.H.; Kim, Y.J.; Kang, S.S.; Baek, S.H. Group IB secretory phospholipase A2 promotes matrix metalloproteinase-2-mediated cell migration via the phosphatidylinositol 3-kinase and Akt pathway. J. Biol. Chem. 2004, 279, 36579–36585. [Google Scholar]
- Zhou, D.H.; Wang, X.; Yang, M.; Shi, X.; Huang, W.; Feng, Q. Combination of Low Concentration of (−)-Epigallocatechin Gallate (EGCG) and Curcumin Strongly Suppresses the Growth of Non-Small Cell Lung Cancer in Vitro and in Vivo through Causing Cell Cycle Arrest. Int. J. Mol. Sci. 2013, 14, 12023–12036. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, H.J.; Jeong, S.J.; Lee, H.J.; Lee, E.O.; Kim, H.S.; Zhang, Y.; Ryu, S.Y.; Lee, M.H.; Lu, J.; Kim, S.H. Galbanic acid isolated from Ferula assafoetida exerts in vivo anti-tumor activity in association with anti-angiogenesis and anti-proliferation. Pharm. Res. 2011, 28, 597–609. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chou, P.Y.; Chien, Y.C.; Wu, C.H.; Wu, T.S.; Sheu, M.J. Ethanol extracts of fruiting bodies of Antrodia cinnamomea exhibit anti-migration action in human adenocarcinoma CL1-0 cells through the MAPK and PI3K/AKT signaling pathways. Phytomedicine 2012, 19, 768–778. [Google Scholar] [CrossRef]
- Wang, L.Q.; Shi, H.S.; Wang, Y.S. Liposomal curcumin inhibits tumor growth and angiogenesis in Lewis lung cancer. Sichuan Da Xue Xue Bao Yi Xue Ban 2013, 44, 46–48. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, S.-C.; Chien, Y.-C.; Pan, C.-H.; Sheu, J.-H.; Chen, C.-Y.; Wu, C.-H. Inhibitory Effect of Dihydroaustrasulfone Alcohol on the Migration of Human Non-Small Cell Lung Carcinoma A549 Cells and the Antitumor Effect on a Lewis Lung Carcinoma-Bearing Tumor Model in C57BL/6J Mice. Mar. Drugs 2014, 12, 196-213. https://doi.org/10.3390/md12010196
Chen S-C, Chien Y-C, Pan C-H, Sheu J-H, Chen C-Y, Wu C-H. Inhibitory Effect of Dihydroaustrasulfone Alcohol on the Migration of Human Non-Small Cell Lung Carcinoma A549 Cells and the Antitumor Effect on a Lewis Lung Carcinoma-Bearing Tumor Model in C57BL/6J Mice. Marine Drugs. 2014; 12(1):196-213. https://doi.org/10.3390/md12010196
Chicago/Turabian StyleChen, Shuo-Chueh, Yi-Chung Chien, Chun-Hsu Pan, Jyh-Horng Sheu, Chih-Yi Chen, and Chieh-Hsi Wu. 2014. "Inhibitory Effect of Dihydroaustrasulfone Alcohol on the Migration of Human Non-Small Cell Lung Carcinoma A549 Cells and the Antitumor Effect on a Lewis Lung Carcinoma-Bearing Tumor Model in C57BL/6J Mice" Marine Drugs 12, no. 1: 196-213. https://doi.org/10.3390/md12010196
APA StyleChen, S. -C., Chien, Y. -C., Pan, C. -H., Sheu, J. -H., Chen, C. -Y., & Wu, C. -H. (2014). Inhibitory Effect of Dihydroaustrasulfone Alcohol on the Migration of Human Non-Small Cell Lung Carcinoma A549 Cells and the Antitumor Effect on a Lewis Lung Carcinoma-Bearing Tumor Model in C57BL/6J Mice. Marine Drugs, 12(1), 196-213. https://doi.org/10.3390/md12010196