First Report of a Peroxiredoxin Homologue in Jellyfish: Molecular Cloning, Expression and Functional Characterization of CcPrx4 from Cyanea capillata

Abstract
:1. Introduction
2. Materials and Methods
2.2. EST Analysis and Identification of CcPrx4
2.3. Sequence Analysis of the Full-Length CcPrx4 cDNA
2.4. Quantification Analysis of CcPrx4 Expression by Quantitative Real-Time PCR
2.5. Construction of the Recombinant Plasmid CcPrx4/pET-24a
2.6. Expression and Purification of Recombinant CcPrx4 Protein in E. coli
2.7. Western Blotting
2.8. In Vitro Peroxidase Activity Assay
2.9. Metal-Catalyzed Oxidation (MCO) Assay
3. Results
3.1. Identification and Sequence Analysis of CcPrx4 cDNA

3.2. Sequence Alignment and Phylogenetic Analysis of the CcPrx4 Protein


{kind=link}
{kind=link}
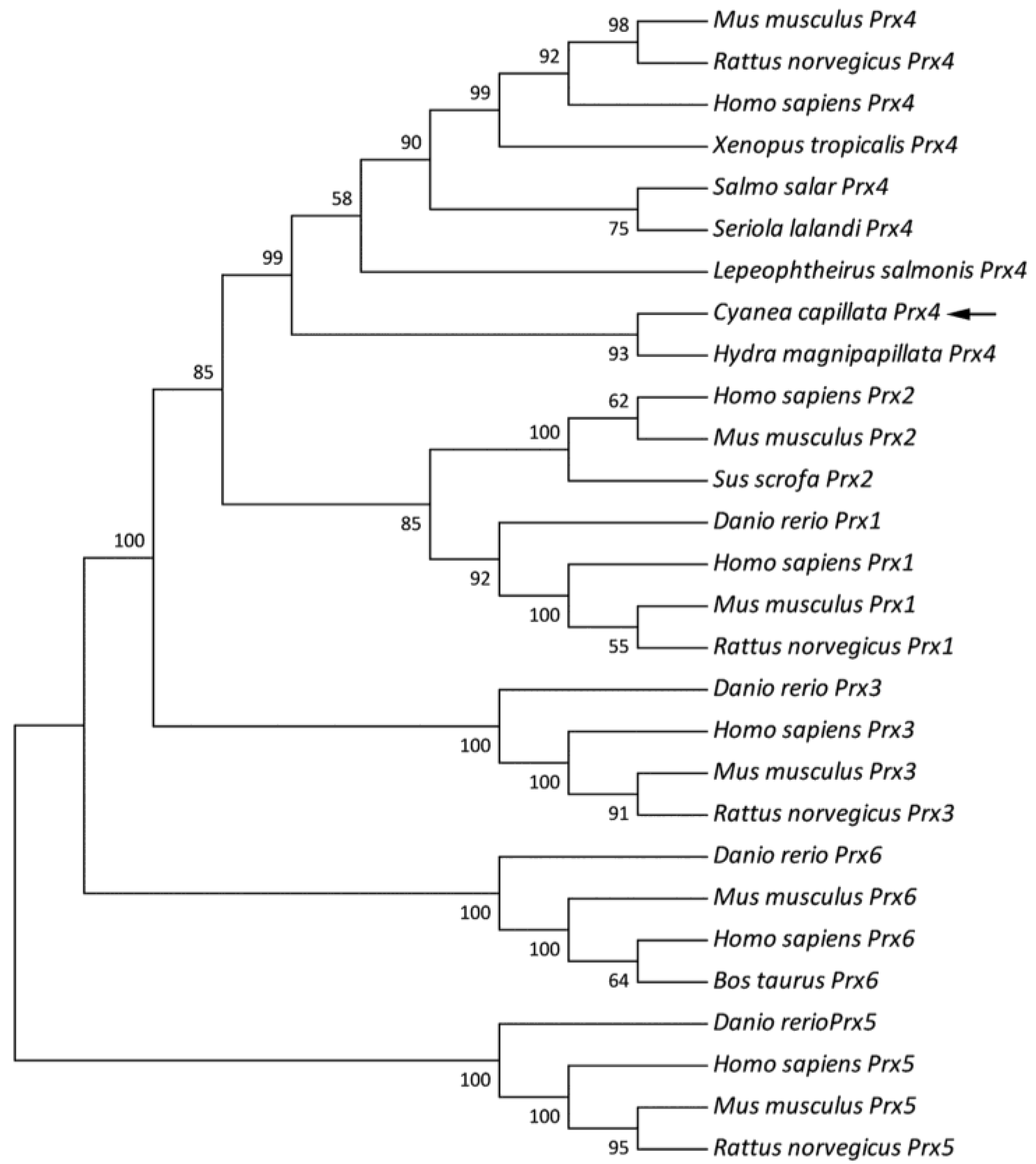{kind=link}
{kind=link}
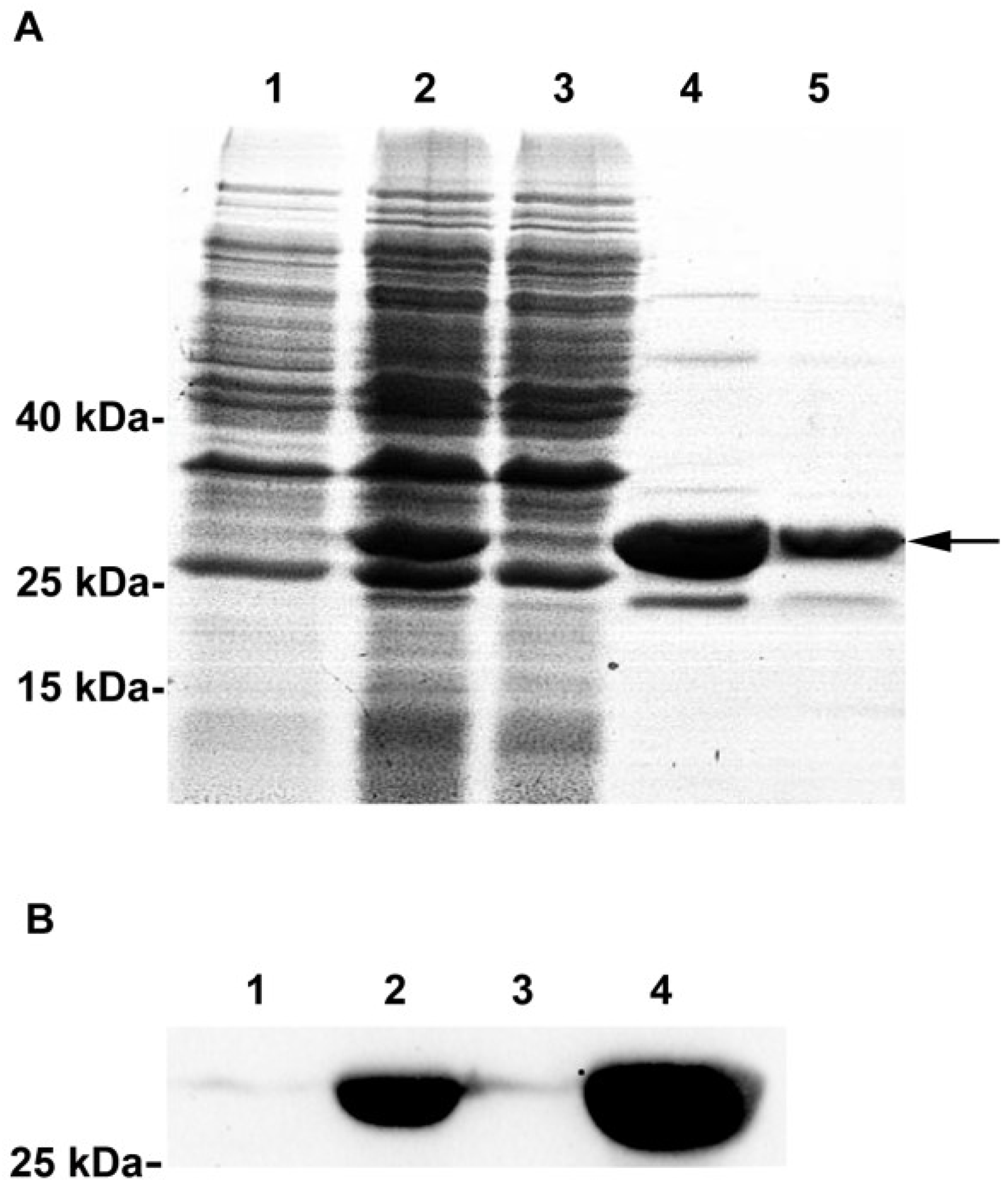{kind=link}
{kind=link}
| Species Name | Common Name | Accession Number | Sequence Size (aa) | Identity (%) | Similarity (%) |
|---|---|---|---|---|---|
| C. capillata | Jellyfish | KF201511 | 247 | - | - |
| H. magnipapillata | Hydra | XP_004207404 | 247 | 74.1 | 81.0 |
| L. salmonis | Sea louse | ACO12581 | 236 | 65.6 | 78.5 |
| M. musculus | Mouse | NP_058044 | 274 | 66.4 | 76.2 |
| R. norvegicus | Norway rat | NP_445964 | 273 | 65.2 | 75.7 |
| H. sapiens | Human | NP_006397 | 271 | 65.0 | 77.0 |
| S. scrofa | Pig | XP_001927404 | 272 | 64.4 | 76.7 |
| X. tropicalis | Western clawed frog | NP_001006812 | 271 | 69.7 | 78.8 |
| S. lalandi | Yellowtail kingfish | ACM47312 | 264 | 68.2 | 77.9 |
| S. salar | Atlantic salmon | ACI69656 | 262 | 68.3 | 77.4 |

3.3. Tissue Distribution of CcPrx4

3.4. Recombinant Expression and Purification of the CcPrx4 Protein

3.5. In Vitro Peroxidase Activity of the CcPrx4 Protein

3.6. Ability of the CcPrx4 Protein to Protect Supercoiled DNA
4. Discussion
5. Conclusions
Abbreviations
| BLAST | Basic Local Alignment Search Tool |
| CDD | Conserved Domain Database |
| MEGA | Molecular Evolutionary Genetics Analysis |
| ROX | Carboxy-X-rhodamine |
| SYBR | Synergy Brands |
| Ct | cycle threshold |
| TOP 10 | dh10 β Escherichia coli strain |
| HRP | horse radish peroxidase |
| G:BOX | Syngene system for fluorescence and visible applications |
| PDB | Protein Data Bank |
| YF motif | Tyr-Phe motif |
Acknowledgments
Conflicts of Interest
References
- Aispuro-Hernandez, E.; Garcia-Orozco, K.D.; Muhlia-Almazan, A.; Del-Toro-Sanchez, L.; Robles-Sanchez, R.M.; Hernandez, J.; Gonzalez-Aguilar, G.; Yepiz-Plascencia, G.; Sotelo-Mundo, R.R. Shrimp thioredoxin is a potent antioxidant protein. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 148, 94–99. [Google Scholar] [CrossRef]
- Mates, J.M.; Perez-Gomez, C.; Nunez de Castro, I. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, B.; Wang, Q.; Zhang, Z.; Nie, F.; Liu, G.; Zheng, J.; Xiao, L.; Zhang, L. Pharmacological studies of tentacle extract from the jellyfish Cyanea capillata in isolated rat aorta. Mar. Drugs 2013, 11, 3335–3349. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef]
- Ataya, F.S.; Fouad, D.; Al-Olayan, E.; Malik, A. Molecular cloning, characterization and predicted structure of a putative copper-zinc SOD from the camel, Camelus dromedarius. Int. J. Mol. Sci. 2012, 13, 879–900. [Google Scholar] [CrossRef]
- Brinkman, D.; Burnell, J. Identification, cloning and sequencing of two major venom proteins from the box jellyfish, Chironex fleckeri. Toxicon 2007, 50, 850–860. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, J.; Torrens, E.; Segura-Puertas, L. Partial purification and characterization of a novel neurotoxin and three cytolysins from box jellyfish (Carybdea marsupialis) nematocyst venom. Arch. Toxicol. 2006, 80, 163–168. [Google Scholar] [CrossRef]
- Leone, A.; Lecci, R.M.; Durante, M.; Piraino, S. Extract from the zooxanthellate jellyfish Cotylorhiza tuberculata modulates gap junction intercellular communication in human cell cultures. Mar. Drugs 2013, 11, 1728–1762. [Google Scholar] [CrossRef]
- Li, R.; Yu, H.; Xing, R.; Liu, S.; Qing, Y.; Li, K.; Li, B.; Meng, X.; Cui, J.; Li, P. Isolation, identification and characterization of a novel antioxidant protein from the nematocyst of the jellyfish Stomolophus meleagris. Int. J. Biol. Macromol. 2012, 51, 274–278. [Google Scholar] [CrossRef]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish (Rhopilema esculentum) on mice skin photoaging induced by UV irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar] [CrossRef]
- Park, S.G.; Cha, M.K.; Jeong, W.; Kim, I.H. Distinct physiological functions of thiol peroxidase isoenzymes in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 5723–5732. [Google Scholar] [CrossRef]
- Chae, J.I.; Cho, Y.K.; Cho, S.K.; Kim, J.H.; Han, Y.M.; Koo, D.B.; Lee, K.K. Proteomic analysis of pancreas derived from adult cloned pig. Biochem. Biophys. Res. Commun. 2008, 366, 379–387. [Google Scholar] [CrossRef]
- Dayer, R.; Fischer, B.B.; Eggen, R.I.; Lemaire, S.D. The peroxiredoxin and glutathione peroxidase families in Chlamydomonas reinhardtii. Genetics 2008, 179, 41–57. [Google Scholar] [CrossRef]
- Seo, M.S.; Kang, S.W.; Kim, K.; Baines, I.C.; Lee, T.H.; Rhee, S.G. Identification of a new type of mammalian peroxiredoxin that forms an intramolecular disulfide as a reaction intermediate. J. Biol. Chem. 2000, 275, 20346–20354. [Google Scholar] [CrossRef]
- Rhee, S.G.; Chae, H.Z.; Kim, K. Peroxiredoxins: A historical overview and speculative preview of novel mechanisms and emerging concepts in cell signaling. Free Radic. Biol. Med. 2005, 38, 1543–1552. [Google Scholar] [CrossRef]
- Lee, K.S.; Kim, S.R.; Park, N.S.; Kim, I.; Kang, P.D.; Sohn, B.H.; Choi, K.H.; Kang, S.W.; Je, Y.H.; Lee, S.M.; et al. Characterization of a silkworm thioredoxin peroxidase that is induced by external temperature stimulus and viral infection. Insect Biochem. Mol. Biol. 2005, 35, 73–84. [Google Scholar] [CrossRef]
- Randall, L.M.; Ferrer-Sueta, G.; Denicola, A. Peroxiredoxins as preferential targets in H2O2-induced signaling. Methods Enzymol. 2013, 527, 41–63. [Google Scholar]
- Jarvis, R.M.; Hughes, S.M.; Ledgerwood, E.C. Peroxiredoxin 1 functions as a signal peroxidase to receive, transduce, and transmit peroxide signals in mammalian cells. Free Radic. Biol. Med. 2012, 53, 1522–1530. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar]
- NCBI. Basic Local Alignment Search Tool (BLAST) Home Page. Available online: http://www.ncbi.nlm.gov/Blast.cgi (accessed on 13 April 2013).
- NCBI. Open Reading Frame (ORF) Finder Home Page. Available online: http://www.ncbi.nlm.nih.gov/gorf/gorf.html (accessed on 13 April 2013).
- EMBL-EBI. ClustalW2 Home Page. Available online: http://www.ebi.ac.uk/Tools/msa/clustalw2/ (accessed on 13 April 2013).
- Marchler-Bauer, A.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; et al. CDD: Specific functional annotation with the Conserved Domain Database. Nucleic. Acids Res. 2009, 37, D205–D210. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic. Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef]
- EMBL-EBI. InterProScan 4 Home Page. Available online: http://www.ebi.ac.uk/Tools/pfa/iprscan/ (accessed on 18 April 2013).
- NCBI. Conserved Domain Database (CDD) Home Page. Available online: http://www.ncbi.nlm.nih.gov/cdd (accessed on 18 April 2013).
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- CBS. SignalP 4.1 Server Home Page. Available online: http://www.cbs.dtu.dk/services/SignalP/ (accessed on 6 May 2013).
- ExPASy. ProtParam Tool Home Page. Available online: http://web.expasy.org/protparam/ (accessed on 6 May 2013).
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef]
- Kelley, L.A.; Sternberg, M.J. Protein structure prediction on the Web: A case study using the Phyre server. Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef]
- Structural Bioinformatics Group; Imperial College. Phyre2 Home Page. Available online: http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index (accessed on 30 May 2013).
- ExPASy. SWISS-MODEL Home Page. Available online: http://swissmodel.expasy.org/ (accessed on 30 May 2013).
- Bramucci, E.; Paiardini, A.; Bossa, F.; Pascarella, S. PyMod: Sequence similarity searches, multiple sequence-structure alignments, and homology modeling within PyMOL. BMC Bioinforma. 2012, 13, S2:1–S2:6. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pushpamali, W.A.; De Zoysa, M.; Kang, H.S.; Oh, C.H.; Whang, I.; Kim, S.J.; Lee, J. Comparative study of two thioredoxin peroxidases from disk abalone (Haliotis discus discus): cloning, recombinant protein purification, characterization of antioxidant activities and expression analysis. Fish Shellfish Immunol. 2008, 24, 294–307. [Google Scholar] [CrossRef]
- Lim, Y.S.; Cha, M.K.; Kim, H.K.; Uhm, T.B.; Park, J.W.; Kim, K.; Kim, I.H. Removals of hydrogen peroxide and hydroxyl radical by thiol-specific antioxidant protein as a possible role in vivo. Biochem. Biophys. Res. Commun. 1993, 192, 273–280. [Google Scholar] [CrossRef]
- Lim, Y.S.; Cha, M.K.; Yun, C.H.; Kim, H.K.; Kim, K.; Kim, I.H. Purification and characterization of thiol-specific antioxidant protein from human red blood cell: a new type of antioxidant protein. Biochem. Biophys. Res. Commun. 1994, 199, 199–206. [Google Scholar]
- Li, J.; Zhang, W.B.; Loukas, A.; Lin, R.Y.; Ito, A.; Zhang, L.H.; Jones, M.; McManus, D.P. Functional expression and characterization of Echinococcus granulosus thioredoxin peroxidase suggests a role in protection against oxidative damage. Gene 2004, 326, 157–165. [Google Scholar] [CrossRef]
- Loo, G.H.; Schuller, K.A. Cloning and functional characterization of a peroxiredoxin 4 from yellowtail kingfish (Seriola lalandi). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 156, 244–253. [Google Scholar] [CrossRef]
- Wood, Z.A.; Poole, L.B.; Karplus, P.A. Peroxiredoxin evolution and the regulation of hydrogen peroxide signaling. Science 2003, 300, 650–653. [Google Scholar] [CrossRef]
- Konig, J.; Lotte, K.; Plessow, R.; Brockhinke, A.; Baier, M.; Dietz, K.J. Reaction mechanism of plant 2-Cys peroxiredoxin: Role of the C terminus and the quaternary structure. J. Biol. Chem. 2003, 278, 24409–24420. [Google Scholar]
- Kil, I.S.; Lee, S.K.; Ryu, K.W.; Woo, H.A.; Hu, M.C.; Bae, S.H.; Rhee, S.G. Feedback control of adrenal steroidogenesis via H2O2-dependent, reversible inactivation of peroxiredoxin III in mitochondria. Mol. Cell 2012, 46, 584–594. [Google Scholar] [CrossRef]
- Cho, C.S.; Lee, S.; Lee, G.T.; Woo, H.A.; Choi, E.J.; Rhee, S.G. Irreversible inactivation of glutathione peroxidase 1 and reversible inactivation of peroxiredoxin II by H2O2 in red blood cells. Antioxid. Redox Signal. 2010, 12, 1235–1246. [Google Scholar] [CrossRef]
- Suttiprapa, S.; Loukas, A.; Laha, T.; Wongkham, S.; Kaewkes, S.; Gaze, S.; Brindley, P.J.; Sripa, B. Characterization of the antioxidant enzyme, thioredoxin peroxidase, from the carcinogenic human liver fluke, Opisthorchis viverrini. Mol. Biochem. Parasitol. 2008, 160, 116–122. [Google Scholar] [CrossRef]
- Wang, T.; Wen, X.J.; Mei, X.B.; Wang, Q.Q.; He, Q.; Zheng, J.M.; Zhao, J.; Xiao, L.; Zhang, L.M. Lipid peroxidation is another potential mechanism besides pore-formation underlying hemolysis of tentacle extract from the jellyfish Cyanea capillata. Mar. Drugs 2013, 11, 67–80. [Google Scholar] [CrossRef]
- Jeon, S.J.; Ishikawa, K. Characterization of novel hexadecameric thioredoxin peroxidase from Aeropyrum pernix K1. J. Biol. Chem. 2003, 278, 24174–24180. [Google Scholar] [CrossRef]
- Nelson, K.J.; Knutson, S.T.; Soito, L.; Klomsiri, C.; Poole, L.B.; Fetrow, J.S. Analysis of the peroxiredoxin family: Using active-site structure and sequence information for global classification and residue analysis. Proteins 2011, 79, 947–964. [Google Scholar] [CrossRef]
- Cheng, X.J.; Yoshihara, E.; Takeuchi, T.; Tachibana, H. Molecular characterization of peroxiredoxin from Entamoeba moshkovskii and a comparison with Entamoeba histolytica. Mol. Biochem. Parasitol. 2004, 138, 195–203. [Google Scholar] [CrossRef]
- Joung, M.; Yoon, S.; Choi, K.; Kim, J.Y.; Park, W.Y.; Yu, J.R. Characterization of the thioredoxin peroxidase from Cryptosporidium parvum. Exp. Parasitol. 2011, 129, 331–336. [Google Scholar] [CrossRef]
- Hakimi, H.; Asada, M.; Angeles, J.M.; Inoue, N.; Kawazu, S. Cloning and characterization of Plasmodium vivax thioredoxin peroxidase-1. Parasitol. Res. 2012, 111, 525–529. [Google Scholar] [CrossRef]
- Henkle-Duhrsen, K.; Kampkotter, A. Antioxidant enzyme families in parasitic nematodes. Mol. Biochem. Parasitol. 2001, 114, 129–142. [Google Scholar] [CrossRef]
- Fujii, J.; Ikeda, Y. Advances in our understanding of peroxiredoxin, a multifunctional, mammalian redox protein. Redox Rep. 2002, 7, 123–130. [Google Scholar] [CrossRef]
- Hofmann, B.; Hecht, H.J.; Flohe, L. Peroxiredoxins. Biol. Chem. 2002, 383, 347–364. [Google Scholar]
- Dietz, K.J. Plant peroxiredoxins. Annu. Rev. Plant Biol. 2003, 54, 93–107. [Google Scholar] [CrossRef]
- Wood, Z.A.; Schroder, E.; Robin Harris, J.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- McGonigle, S.; Dalton, J.P.; James, E.R. Peroxidoxins: A new antioxidant family. Parasitol. Today 1998, 14, 139–145. [Google Scholar] [CrossRef]
- Haridas, V.; Ni, J.; Meager, A.; Su, J.; Yu, G.L.; Zhai, Y.; Kyaw, H.; Akama, K.T.; Hu, J.; Van Eldik, L.J.; et al. TRANK, a novel cytokine that activates NF-kappa B and c-Jun N-terminal kinase. J. Immunol. 1998, 161, 1–6. [Google Scholar]
- Qiu, L.; Ma, Z.; Jiang, S.; Wang, W.; Zhou, F.; Huang, J.; Li, J.; Yang, Q. Molecular cloning and mRNA expression of peroxiredoxin gene in black tiger shrimp (Penaeus monodon). Mol. Biol. Rep. 2010, 37, 2821–2827. [Google Scholar] [CrossRef]
- Sun, J.; Liu, X.; Li, Q. Molecular cloning, expression and antioxidant activity of a peroxiredoxin 2 homologue from Lampetra japonica. Fish Shellfish Immunol. 2010, 28, 795–801. [Google Scholar] [CrossRef]
- Shi, G.Q.; Yu, Q.Y.; Shi, L.; Zhang, Z. Molecular cloning and characterization of peroxiredoxin 4 involved in protection against oxidative stress in the silkworm Bombyx mori. Insect Mol. Biol. 2012, 21, 581–592. [Google Scholar] [CrossRef]
- Yu, F.; Kang, M.; Meng, F.; Guo, X.; Xu, B. Molecular cloning and characterization of a thioredoxin peroxidase gene from Apis cerana cerana. Insect Mol. Biol. 2011, 20, 367–378. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ruan, Z.; Liu, G.; Wang, B.; Zhou, Y.; Lu, J.; Wang, Q.; Zhao, J.; Zhang, L. First Report of a Peroxiredoxin Homologue in Jellyfish: Molecular Cloning, Expression and Functional Characterization of CcPrx4 from Cyanea capillata. Mar. Drugs 2014, 12, 214-231. https://doi.org/10.3390/md12010214
Ruan Z, Liu G, Wang B, Zhou Y, Lu J, Wang Q, Zhao J, Zhang L. First Report of a Peroxiredoxin Homologue in Jellyfish: Molecular Cloning, Expression and Functional Characterization of CcPrx4 from Cyanea capillata. Marine Drugs. 2014; 12(1):214-231. https://doi.org/10.3390/md12010214
Chicago/Turabian StyleRuan, Zengliang, Guoyan Liu, Beilei Wang, Yonghong Zhou, Jia Lu, Qianqian Wang, Jie Zhao, and Liming Zhang. 2014. "First Report of a Peroxiredoxin Homologue in Jellyfish: Molecular Cloning, Expression and Functional Characterization of CcPrx4 from Cyanea capillata" Marine Drugs 12, no. 1: 214-231. https://doi.org/10.3390/md12010214
APA StyleRuan, Z., Liu, G., Wang, B., Zhou, Y., Lu, J., Wang, Q., Zhao, J., & Zhang, L. (2014). First Report of a Peroxiredoxin Homologue in Jellyfish: Molecular Cloning, Expression and Functional Characterization of CcPrx4 from Cyanea capillata. Marine Drugs, 12(1), 214-231. https://doi.org/10.3390/md12010214