Antimicrobial Activity of Peptides Derived from Olive Flounder Lipopolysaccharide Binding Protein/Bactericidal Permeability-Increasing Protein (LBP/BPI)

Abstract
:1. Introduction
2. Results and Discussion
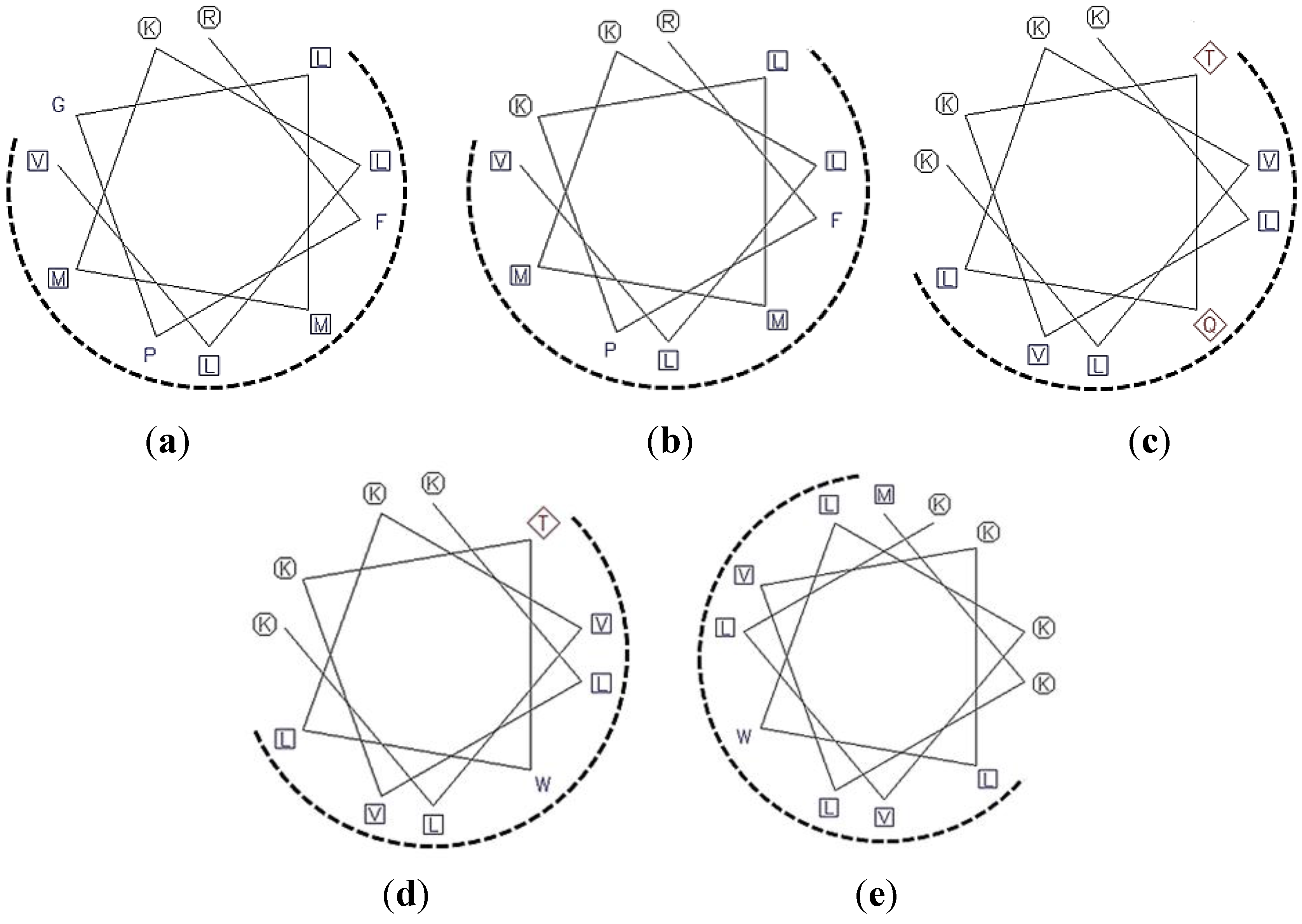
2.1. Peptide Design and Synthesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domain | Amino Acid Sequence in α-Helix Predicted | Position/AA Length | pI Value | Net Charge | Boman Index * (kcal/mol) | Hydrophobic Ratio * (%) | Hydrophobicity* (kcal/mol) | Modification |
|---|---|---|---|---|---|---|---|---|
| N-terminal domain | LEYGRQLGMASIQQKLK | 32–48/17 | 10.21 | +2 | 1.6 | 35 | 3.71 | |
| VEYSLSHMQIVKL | 67–79/13 | 7.76 | 0 | 0.33 | 46 | 1.56 | ||
| WLYNLFKNFIDKALRNALQKQ | 170–190/21 | 10.44 | +3 | 1.66 | 47 | 0.17 | ||
| VNELNPHLKTLNVLAKVDQYAE | 199–220/22 | 5.37 | −1 | 1.4 | 40 | 7.22 | ||
| SIDLN | 235–239/5 | 3.0 | −1 | 1.78 | 40 | 0.91 | ||
| C-terminal domain | AYSVNSAAFVYNK | 276–288/13 | 9.65 | +1 | 0.72 | 46 | −0.27 | |
| EISKRFPGLMMKLLVQ | 320–335/16 | 10.59 | +2 | 0.58 | 50 | 2.47 | ||
| RFPGLMMKLLV a | 324–334/11 | 11.51 | +2 | −0.63 | 63 | −0.94 | ||
| RFPKLMMKLLV b | 324–334/11 | 11.67 | +3 | −0.04 | 63 | 0.04 | Gly→Lys | |
| MRVAGAVSLN | 389–398/10 | 10.9 | +1 | 0.5 | 60 | 1.06 | ||
| FKVRSLDNILQMVLKVVVI | 412–430/19 | 10.53 | +1 | 0.52 | 63 | 1.84 | ||
| KMKLVKTQLKVLKDYMLI | 448–465/18 | 10.45 | +4 | 0.4 | 50 | 3.09 | ||
| KLVKTQLKVLKc | 450–460/11 | 10.98 | +4 | 0.67 | 45 | 3.14 | ||
| KLVKTWLKVLKd | 450–460/11 | 10.98 | +4 | −0.03 | 54 | 0.71 | Gln→Trp | |
| MKLVKTWLKVLKe | 449–460/12 | 10.99 | +4 | −0.22 | 58 | 0.48 | Gln→Trp |
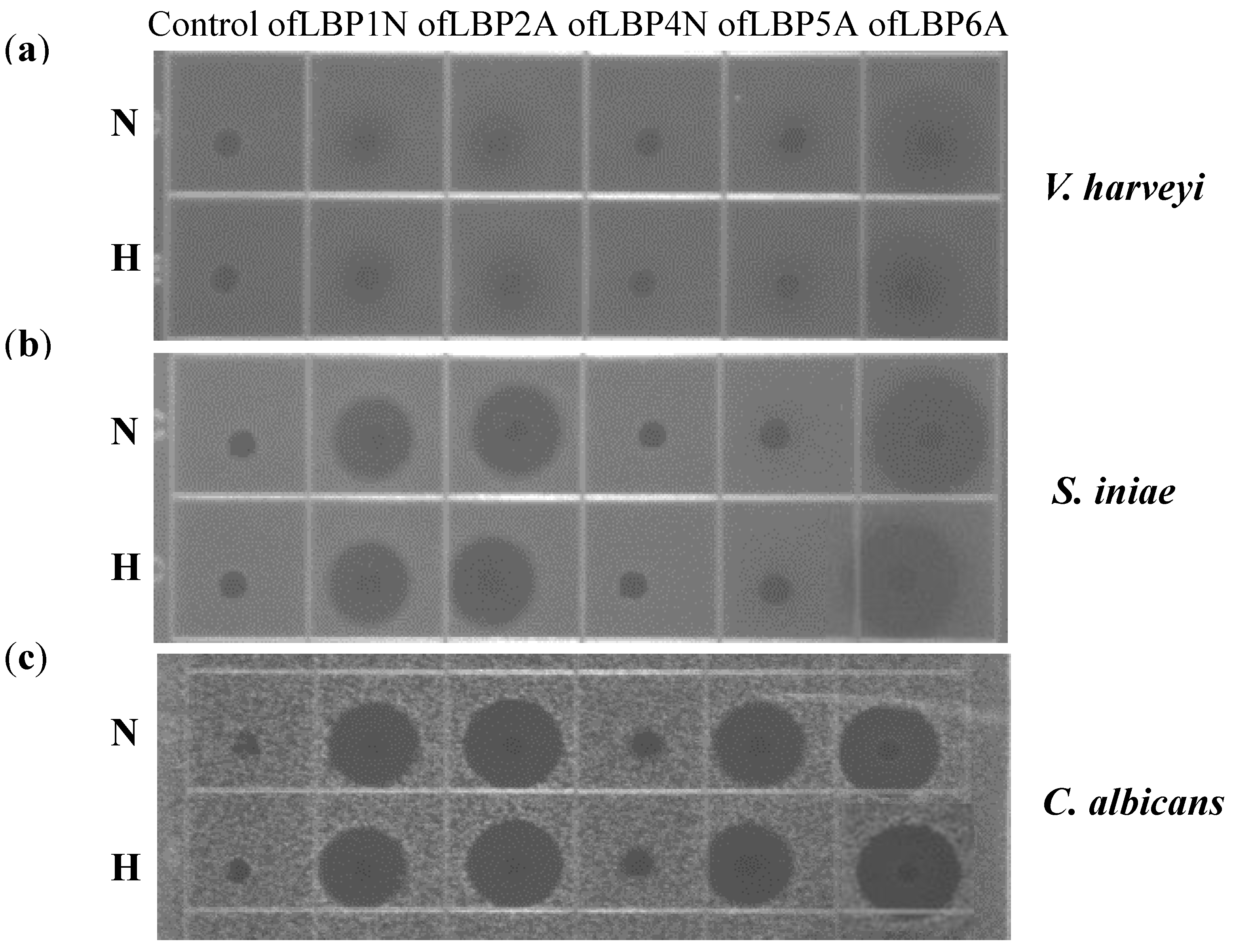
2.2. Antimicrobial and Hemolytic Activities

| Microbes | Gram | Minimal Effectives Concentration (μg/mL) | |||||
|---|---|---|---|---|---|---|---|
| ofLBP 1N | ofLBP 2A | ofLBP 4N | ofLBP 5A | ofLBP 6A | Piscidin 1 | ||
| B. subtilis KCTC1021 | + | 1.4 | 0.8 | 62.5 | 0.7 | 1.4 | 2.3 |
| B. subtilis RM125 | + | 1.0 | 1.2 | 250.0 | 4.2 | 1.8 | 4.5 |
| M. luteus ATCC9341 | + | 5.3 | 5.7 | 250.0 | 8.0 | 2.2 | 2.3 |
| S. aureus RM4220 | + | 5.6 | 5.0 | 62.5 | 3.7 | 2.7 | 2.2 |
| E. coli D31 | − | 1.3 | 0.5 | 62.5 | 0.9 | 2.6 | 2.0 |
| E. coli KCTC1116 | − | 1.7 | 1.2 | 1.5 | 1.6 | 3.8 | 7.0 |
| E. coli ML35p | − | 0.8 | 0.6 | 250.0 | 1.6 | 1.8 | 2.3 |
| P. aeruginosa KCTC2004 | − | 6.4 | 6.6 | >250.0 | 8.0 | 4.6 | 8.0 |
| S. enterica KCTC2514 | − | 1.2 | 1.2 | 250.0 | 3.5 | 2.6 | 7.0 |
| Fish pathogen | |||||||
| A. hydrophila KCTC2358 | − | 4.8 | 3.6 | >250.0 | 5.0 | 2.8 | 10.0 |
| E. tarda H-4 | − | 125.0 | 62.5 | >250.0 | 250.0 | 62.5 | 125.0 |
| S. iniae FP5229 | + | 5.3 | 3.0 | >250.0 | 2.7 | 1.2 | 6.5 |
| V. parahemolyticus KCCM41664 | − | 2.2 | 1.7 | 1.3 | 1.2 | 0.9 | 1.8 |
| C. albicans KCTC7965 | Yeast | 5.9 | 5.6 | 125.0 | 4.0 | 2.7 | >62.5 |

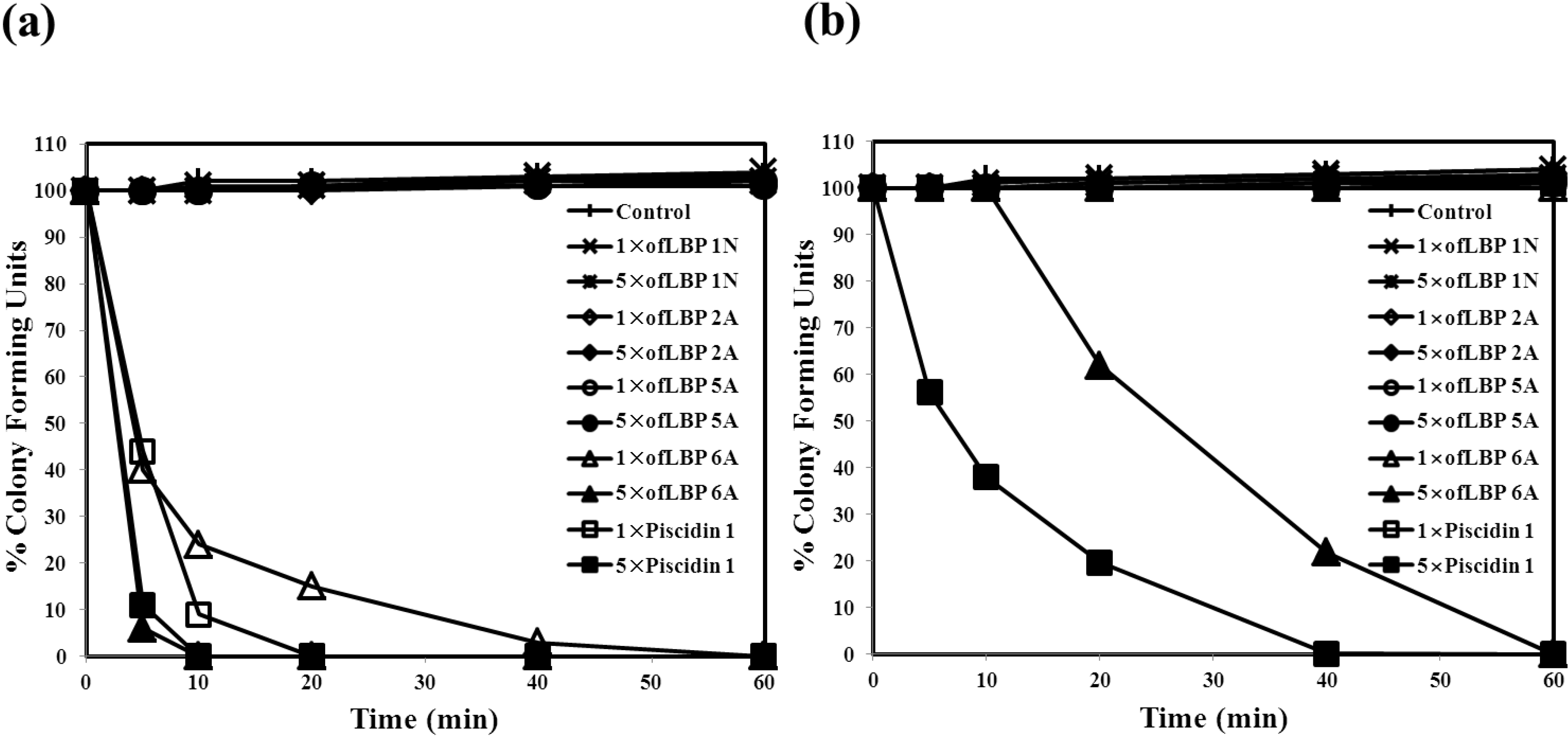
2.3. Killing Kinetic Study

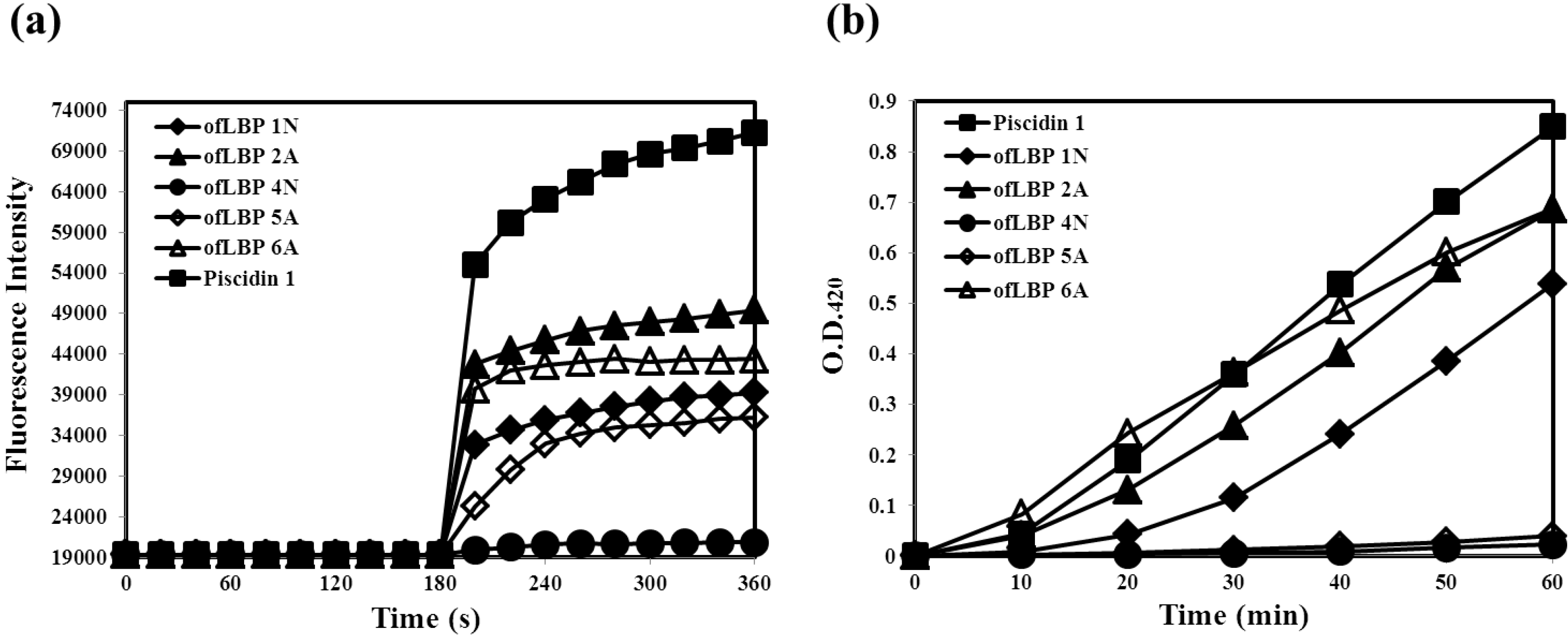
2.4. Membrane Permeabilization Ability

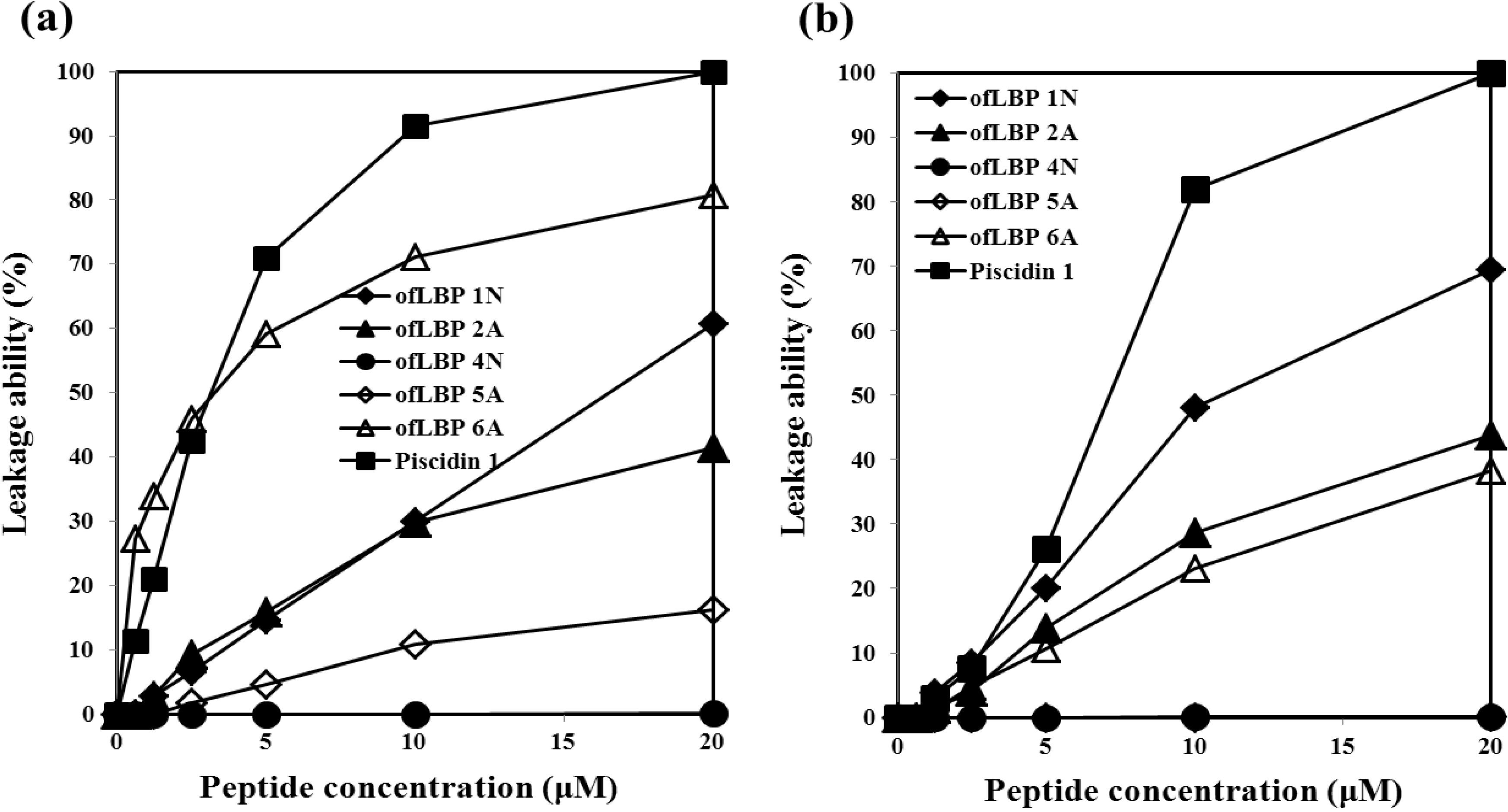
2.5. Leakage Ability of AMPs

2.6. Effect of Temperature on Antimicrobial Activity
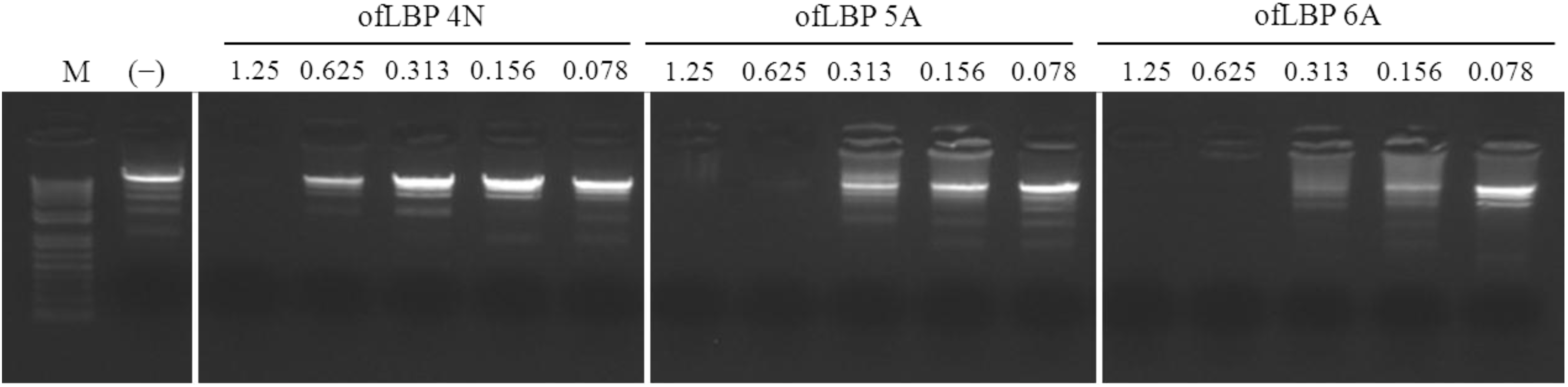
2.7. DNA-Binding Assay

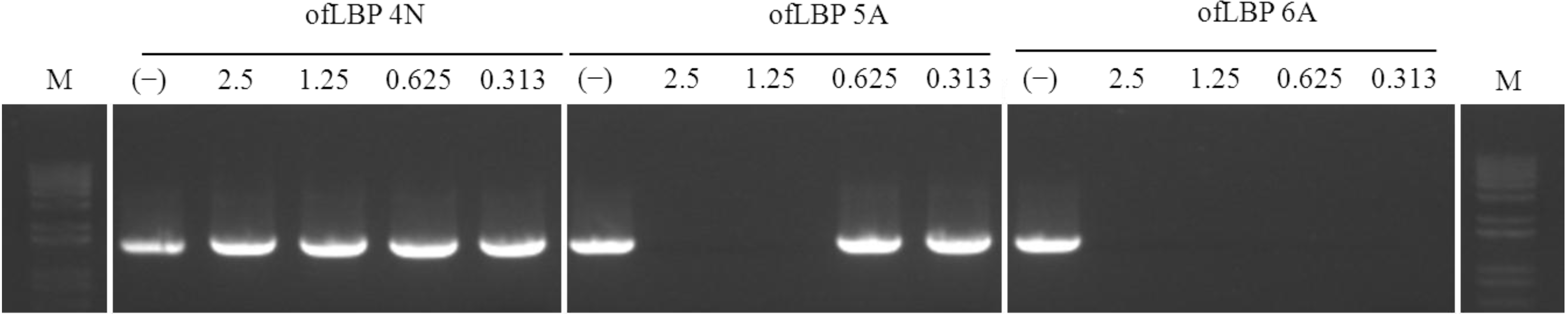
2.8. DNA polymerase Inhibition Assay


3. Experimental Section
3.1. Peptide Synthesis and Purification
3.2. Ultrasensitive Radial Diffusion Assay for Antimicrobial Potency
3.3. Minimal Effective Concentration of the ofLBP-Derived Analogs
3.4. Hemolytic Activity Assay
3.5. Killing Kinetic Assay
3.6. Membrane Permeabilization
3.7. Liposome Preparation
3.8. Leakage Assay
3.9. Effect of Temperature on Antimicrobial Activity
3.10. DNA-Binding Assay
3.11. DNA Polymerase Inhibition Assay
3.12. Structure Prediction
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hancock, R.E.W.; Patrzykat, A. Clinical development of cationic antimicrobial peptides, from natural to novel antibiotics. Curr. Drug Targets 2002, 2, 79–83. [Google Scholar]
- Jiang, S.; Hong, Z.; Guo, W.; Xiaoyun, G.; Gengfa, L.; Yongning, L.; Guangxia, X. A synthetic peptide derived from bactericidal/permeability-increasing protein neutralizes endotoxin in vitro and in vivo. Int. Immunopharmacol. 2004, 4, 527–537. [Google Scholar]
- He, J.; Eckert, R.; Pharm, T.; Simanian, M.D.; Hu, C.; Yarbrough, D.K.; Qi, F.; Anderson, M.H.; Shi, W. Novel synthetic antimicrobial peptides against Streptococcus mutans. Antimicrob. Agents Chemother. 2007, 51, 1351–1358. [Google Scholar]
- Schumann, R.R.; Leong, S.R.; Flaggs, G.W.; Gray, P.W.; Wright, S.D.; Mathison, J.C.; Tobias, P.S.; Ulevitch, R.J. Structure and function of lipopolysaccharide binding protein. Science 1990, 249, 1429–1431. [Google Scholar]
- Beamer, L.J.; Carroll, S.F.; Eisenberg, D. Crystal structure of human BPI and two bound phospholipids at 2.4 angstrom resolution. Science 1997, 276, 1861–1864. [Google Scholar]
- Kirschning, C.J.; Au-Young, J.; Lamping, N.; Reuter, D.; Pfeil, D.; Seilhamer, J.J.; Schumann, R.R. Similar organization of the lipopolysaccharide-binding protein (LBP) and phospholipid transfer protein (PLTP) genes suggests a common gene family of lipid-binding proteins. Genomics 1997, 46, 416–425. [Google Scholar]
- Elsbach, P.; Weiss, J. Prospects for use of recombinant BPI in the treatment of gram-negative bacterial infections. Infect. Agents Dis. 1995, 4, 102–109. [Google Scholar]
- Inagawa, H.; Honda, T.; Kohchi, C.; Nishizawa, T.; Yoshiura, Y.; Nakanishi, T.; Yokomizo, Y.; Soma, G. Cloning and characterization of the homolog of mammalian lipopolysaccharide binding protein and bactericidal permeability-increasing protein in rainbow trout Oncorhynchus mykiss. J. Immunol. 2002, 168, 5638–5644. [Google Scholar]
- Kono, T.; Sakai, M. Molecular cloning of a novel bactericidal permeability-increasing protein/lipopolysaccharide-binding protein (BPI/LBP) from common carp Cyprinus carpio L. and its expression. Mol. Immunol. 2003, 40, 269–278. [Google Scholar]
- Stenvik, J.; Soltrad, T.; Strand, C.; Leiros, I.; Jorgensen, T. Cloning and analyses of BPI/LBP cDNA of the Atlantic cod (Gadus morhua L.). Dev. Comp. Immunol. 2004, 28, 307–323. [Google Scholar]
- Nam, B.H.; Ahn, K.J.; Kim, Y.O.; Kong, H.J.; Kim, W.J.; Kim, H.S.; Lee, S.J.; Kim, K.K. Molecular cloning and characterization of LPS-binding protein/bactericidal permeability-increasing protein (LBP/BPI) from olive flounder, Paralichthys olivaceus. Vet. Immunol. Immunopathol. 2010, 133, 256–263. [Google Scholar]
- Fenton, M.J.; Golenbock, D.T. LPS-binding proteins and receptors. J. Leuk. Biol. 1998, 64, 25–32. [Google Scholar]
- Gazzano-Santoro, H.; Parent, J.B.; Grinna, L.; Horwitz, A.; Parsons, T.; Theofan, G.; Elsbach, P.; Weiss, J.; Conlon, P.J. High affinity binding of the bactericidal/permeability-increasing protein from human neutrophils. Infect. Immun. 1992, 60, 4754–4761. [Google Scholar]
- Weiss, J.; Elsbach, P.; Shu, C.; Castillo, J.; Grinna, L.; Horwits, A.; Theofan, G. Human bactericidal/permeability-increasing protein and a recombinant NH2-terminal fragment cause killing of serum-resistant Gram-negative bacteria in whole blood and inhibit tumor necrosis factor release induced by the bacteria. J. Clin. Invest. 1992, 90, 1122–1130. [Google Scholar]
- Little, R.G.; Kelner, D.N.; Lim, E.; Burke, D.J.; Conlon, P.J. Functional domains of recombinant bactericidal/permeability increasing protein (rBPI23). J. Biol. Chem. 1994, 269, 1865–1872. [Google Scholar]
- Iovine, N.M.; Elsbach, P.; Weiss, J. An opsonic function of the neutrophil bactericidal/permeability-increasing protein depends on both its N- and C-terminal domains. Proc. Natl. Acad. Sci. USA 1997, 94, 10973–10978. [Google Scholar]
- ExPASy. Available online: http://www.expasy.org/tools/ (accessed on 25 June 2013).
- Boman, H. Antibacterial peptides, basic facts and emerging concepts. J. Int. Med. 2003, 254, 197–215. [Google Scholar]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar]
- Cantisani, M.; Leone, M.; Mignogna, E.; Kampanaraki, K.; Falanga, A.; Morelli, G.; Galdiero, M.; Galdiero, S. Structure–activity relations of myxinidin, an antibacterial peptide derived from the epidermal mucus of hagfish. Antimicrob. Agents Chemother. 2013, 57, 5665–5673. [Google Scholar]
- Koo, Y.S.; Kim, J.M.; Park, I.Y.; Yu, B.J.; Jang, S.A.; Kim, K.S.; Park, C.B.; Cho, J.H.; Kim, S.C. Structure-activity relations of parasin I, a histone H2A-related antimicrobial peptide. Peptides 2008, 29, 1102–1108. [Google Scholar]
- Park, N.G.; Silphaduang, U.; Moon, H.S.; Seo, J.K.; Corrales, J.; Noga, E.J. Structure-activity relationships of piscidin 4, a piscine antimicrobial peptide. Biochemistry 2011, 50, 3288–3299. [Google Scholar]
- Barthe, C.; Hocquellet, A.; Garbay, B. Bacteriostatic activity of the proregion of human hepcidin. Prot. Pept. Lett. 2011, 18, 36–40. [Google Scholar]
- Seo, J.K.; Crawford, J.M.; Stone, K.L.; Noga, E.J. Purification of a novel arthropod defensin from the American oyster, Crassostrea virginica. Biochem. Biophys. Res. Commun. 2005, 338, 1998–2004. [Google Scholar]
- Lehrer, R.I.; Rosenman, M.; Harwig, S.S.L.; Jackson, R.; Eisenhaur, P. Ultrasensitive assay for endogenous antimicrobial polypeptides. J. Immunol. Methods 1991, 137, 167–173. [Google Scholar]
- Silphaduang, U.; Noga, E.J. Peptide antibiotics in mast cells of fish. Nature 2001, 414, 268–269. [Google Scholar]
- Skerlavaj, B.; Romeo, D.; Gennaro, R. Rapid membrane permeabilization and inhibition of vital functions of Gram-negative bacteria by bactenecins. Infect. Immun. 1990, 58, 3724–3730. [Google Scholar]
- Hsu, C.H.; Chen, C.; Jou, M.L.; Lee, A.Y.L.; Lin, Y.C.; Yu, Y.P.; Huang, W.T.; Wu, S.H. Structural and DNA-binding studies on the bovine antimicrobial peptide, indolicidin, evidence for multiple conformations involved in binding to membranes and DNA. Nucl. Acids Res. 2005, 33, 4053–4064. [Google Scholar]
- Alfred, R.L.; Palombo, E.A.; Panozzo, J.F.; Bhave, M. The antimicrobial domains of wheat puroindolines are cell-penetrating peptides with possible intracellular mechanism of action. PLoS One 2013, 8, e75488. [Google Scholar]
- ExPAsy. Available online: http://www.expasy.org/protparam/ (accessed on 25 June 2013).
- Ramachandran, G.N.; Sasisekharan, V. Conformation of polypeptides and proteins. Adv. Prot. Chem. 1968, 23, 283–437. [Google Scholar]
- Antimicrobial Peptide Database. Available online: http://asp.unmc.edu/AP/main.php/ (accessed on 25 June 2013).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, B.-H.; Moon, J.-Y.; Park, E.-H.; Kim, Y.-O.; Kim, D.-G.; Kong, H.J.; Kim, W.-J.; Jee, Y.J.; An, C.M.; Park, N.G.; et al. Antimicrobial Activity of Peptides Derived from Olive Flounder Lipopolysaccharide Binding Protein/Bactericidal Permeability-Increasing Protein (LBP/BPI). Mar. Drugs 2014, 12, 5240-5257. https://doi.org/10.3390/md12105240
Nam B-H, Moon J-Y, Park E-H, Kim Y-O, Kim D-G, Kong HJ, Kim W-J, Jee YJ, An CM, Park NG, et al. Antimicrobial Activity of Peptides Derived from Olive Flounder Lipopolysaccharide Binding Protein/Bactericidal Permeability-Increasing Protein (LBP/BPI). Marine Drugs. 2014; 12(10):5240-5257. https://doi.org/10.3390/md12105240
Chicago/Turabian StyleNam, Bo-Hye, Ji-Young Moon, Eun-Hee Park, Young-Ok Kim, Dong-Gyun Kim, Hee Jeong Kong, Woo-Jin Kim, Young Ju Jee, Cheul Min An, Nam Gyu Park, and et al. 2014. "Antimicrobial Activity of Peptides Derived from Olive Flounder Lipopolysaccharide Binding Protein/Bactericidal Permeability-Increasing Protein (LBP/BPI)" Marine Drugs 12, no. 10: 5240-5257. https://doi.org/10.3390/md12105240
APA StyleNam, B. -H., Moon, J. -Y., Park, E. -H., Kim, Y. -O., Kim, D. -G., Kong, H. J., Kim, W. -J., Jee, Y. J., An, C. M., Park, N. G., & Seo, J. -K. (2014). Antimicrobial Activity of Peptides Derived from Olive Flounder Lipopolysaccharide Binding Protein/Bactericidal Permeability-Increasing Protein (LBP/BPI). Marine Drugs, 12(10), 5240-5257. https://doi.org/10.3390/md12105240