Applications of Mass Spectrometry to Structural Analysis of Marine Oligosaccharides



Abstract
:1. Introduction

2. MS for Characterizing Galactan Oligosaccharides from Red Algae
2.1. MS of Carra-Oligosaccharides and Neocarra-Oligosaccharides

{kind=link}
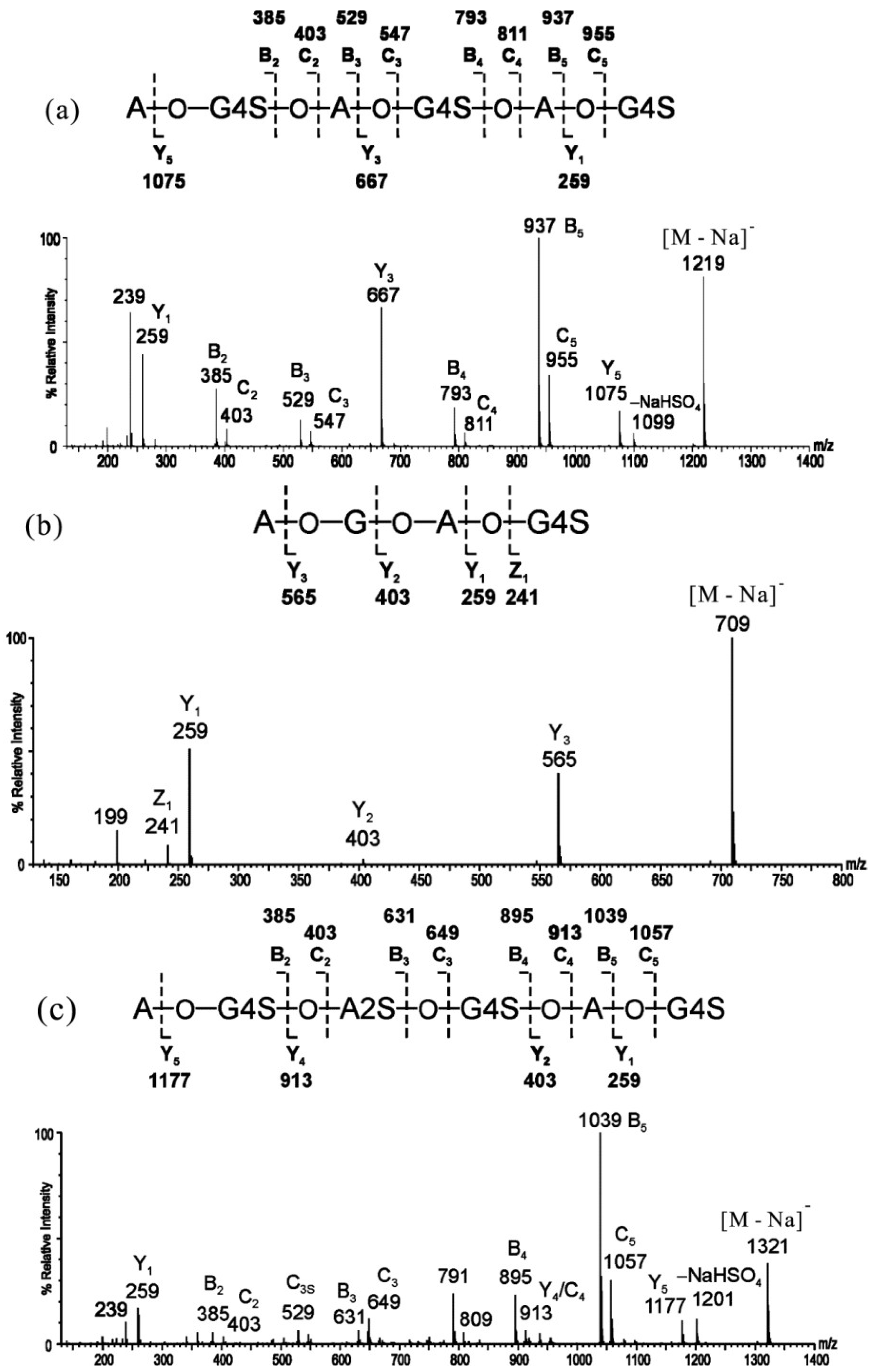{kind=link}
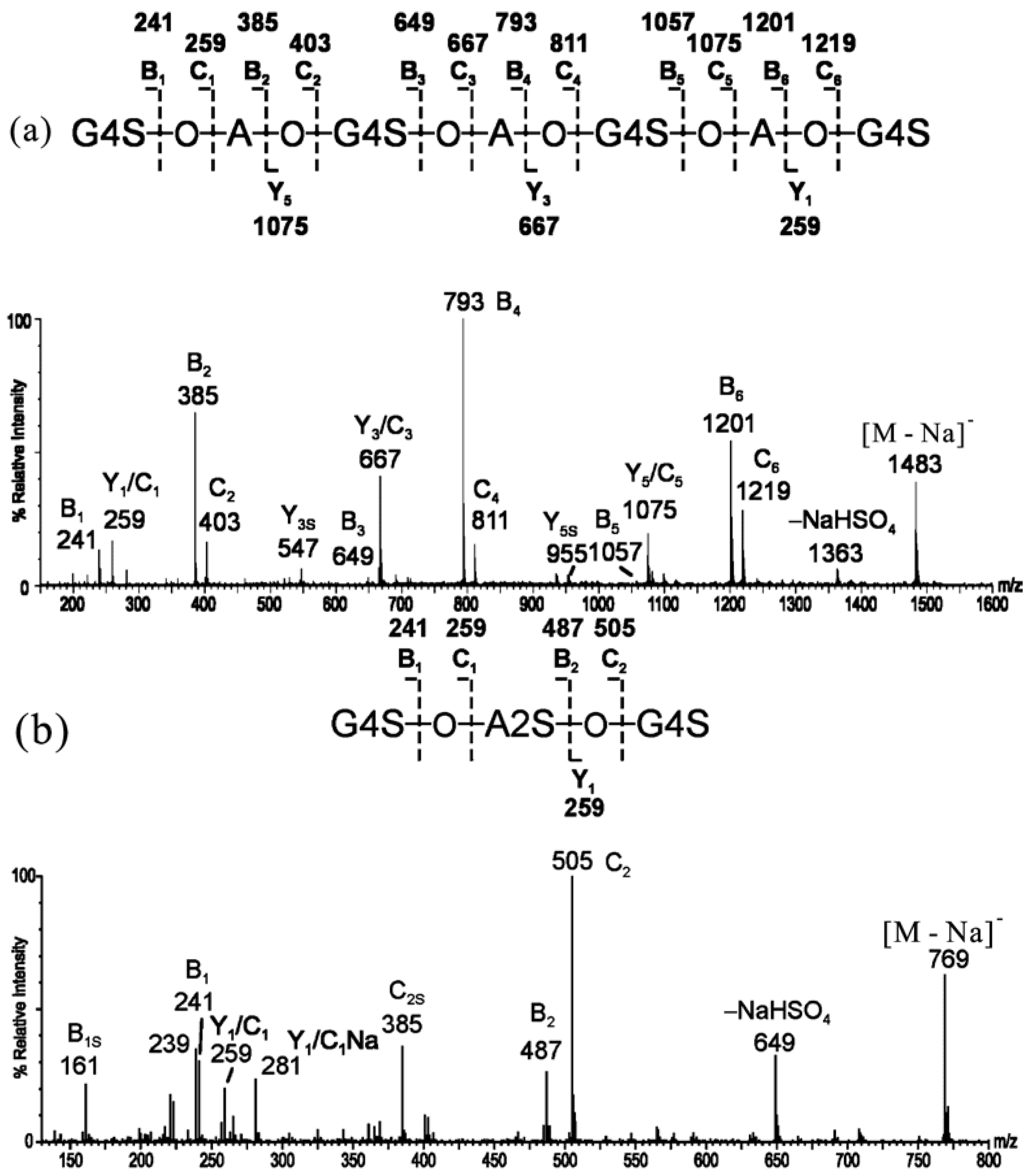{kind=link}
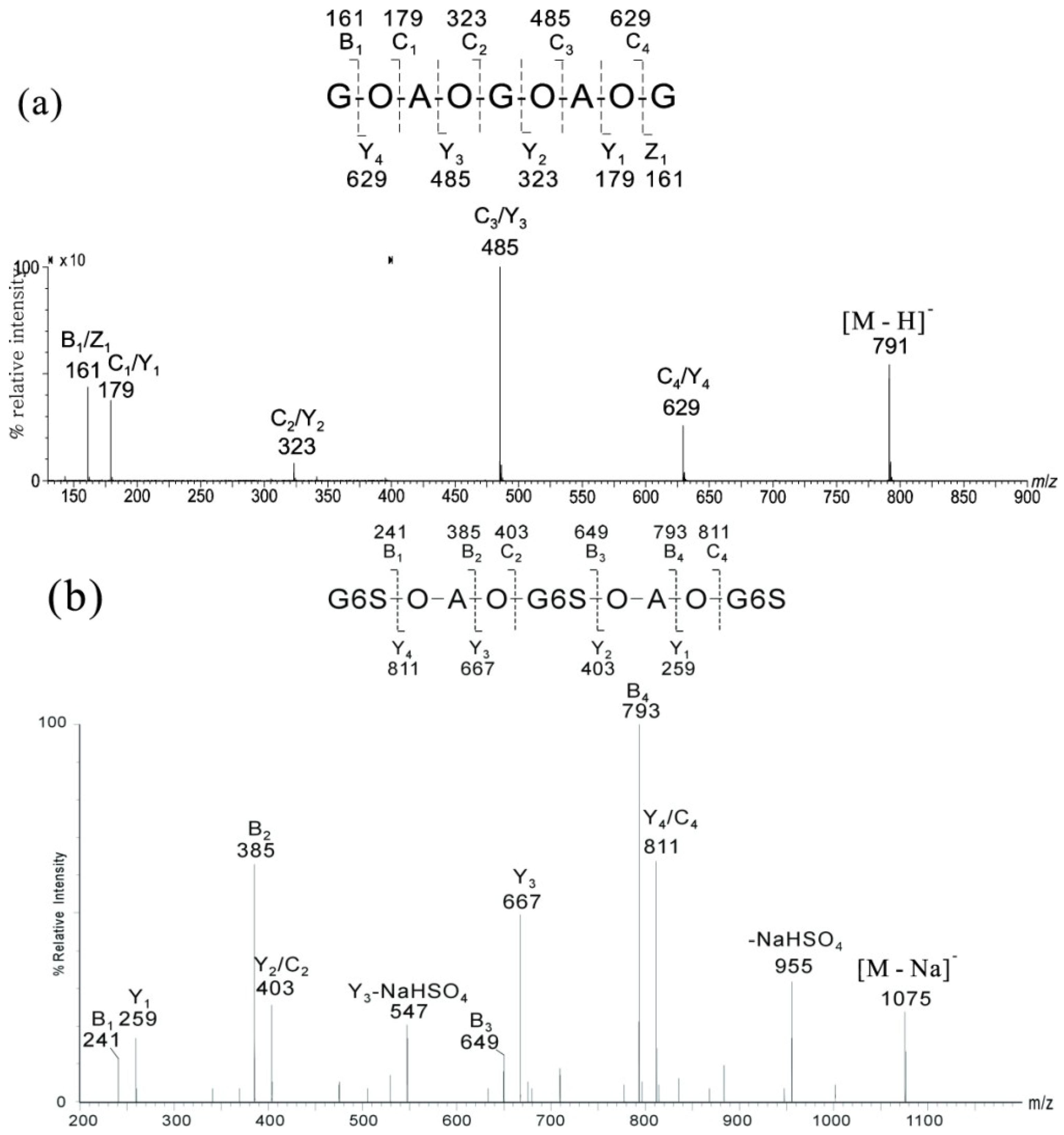{kind=link}
{kind=link}
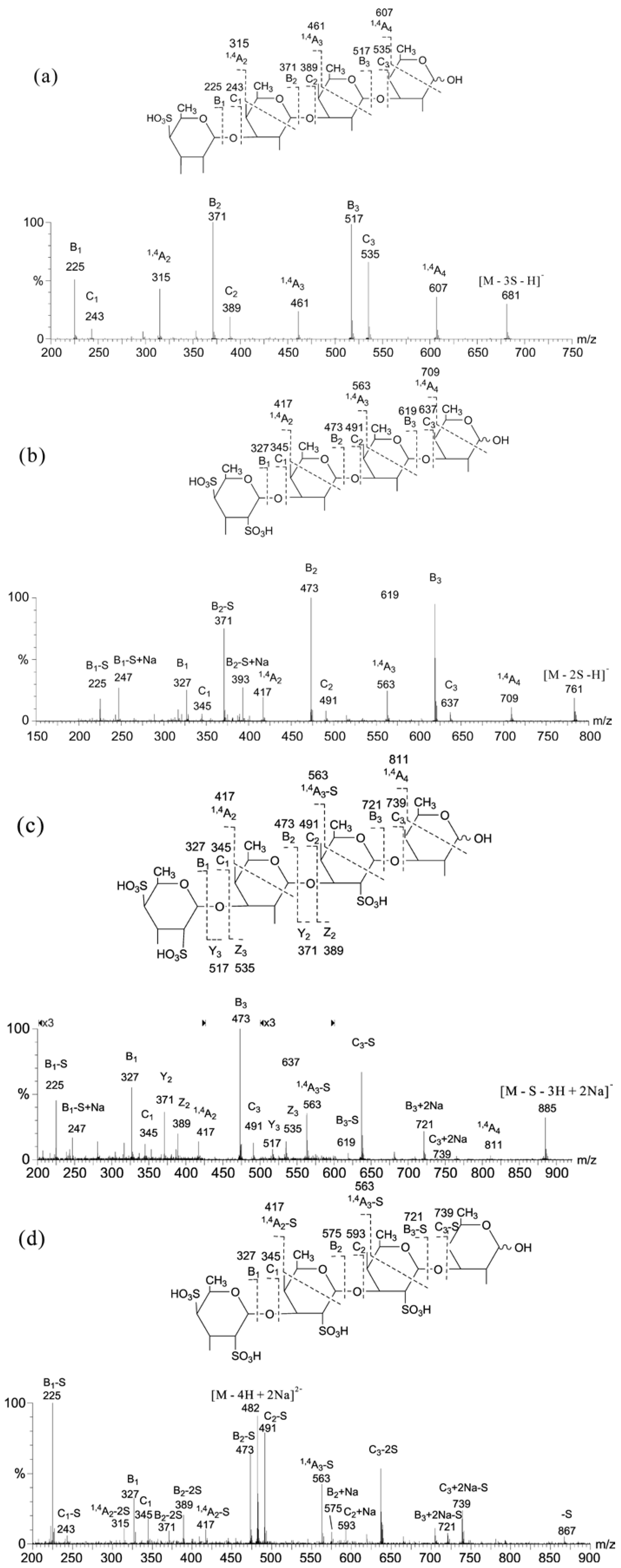{kind=link}
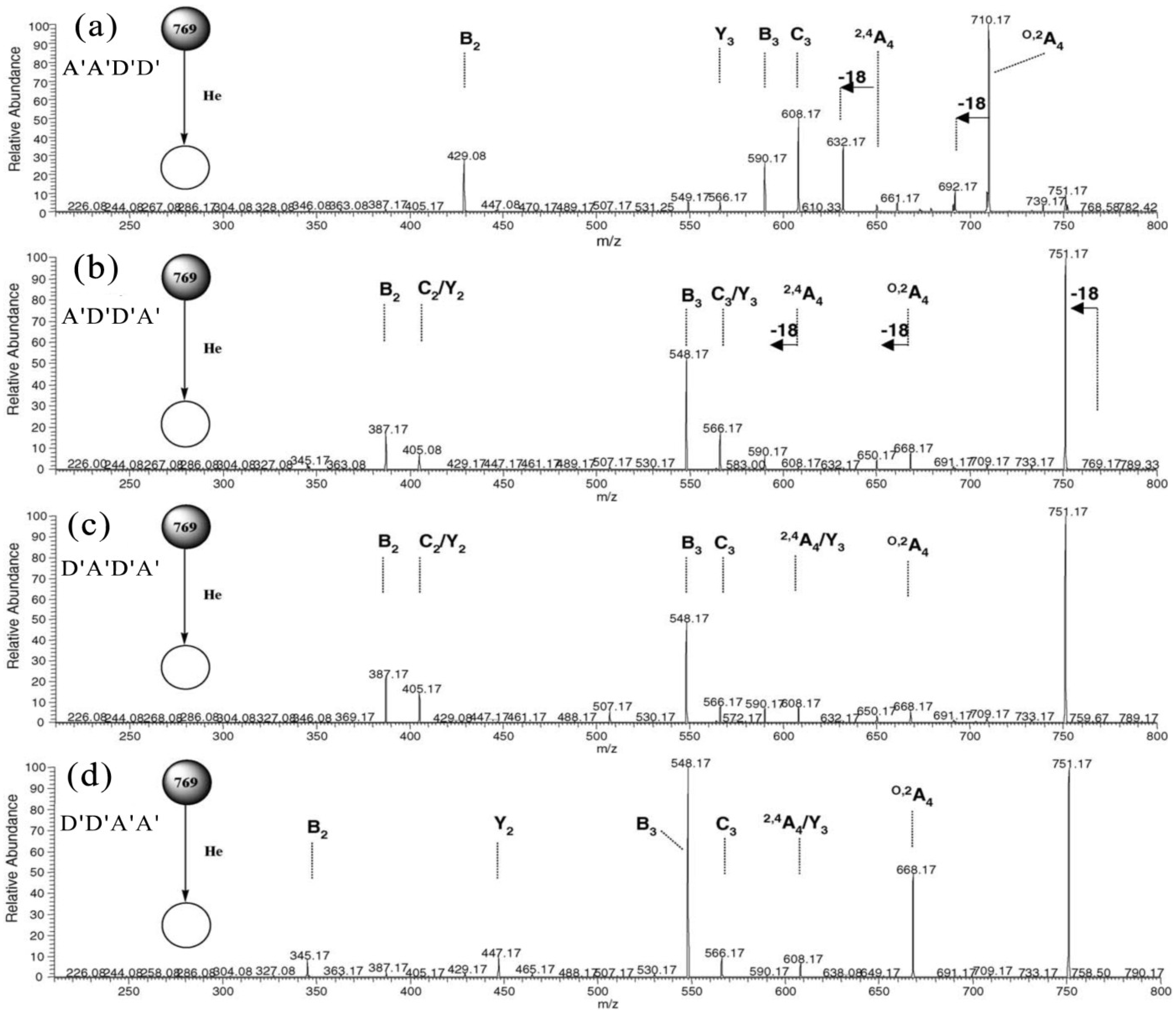{kind=link}
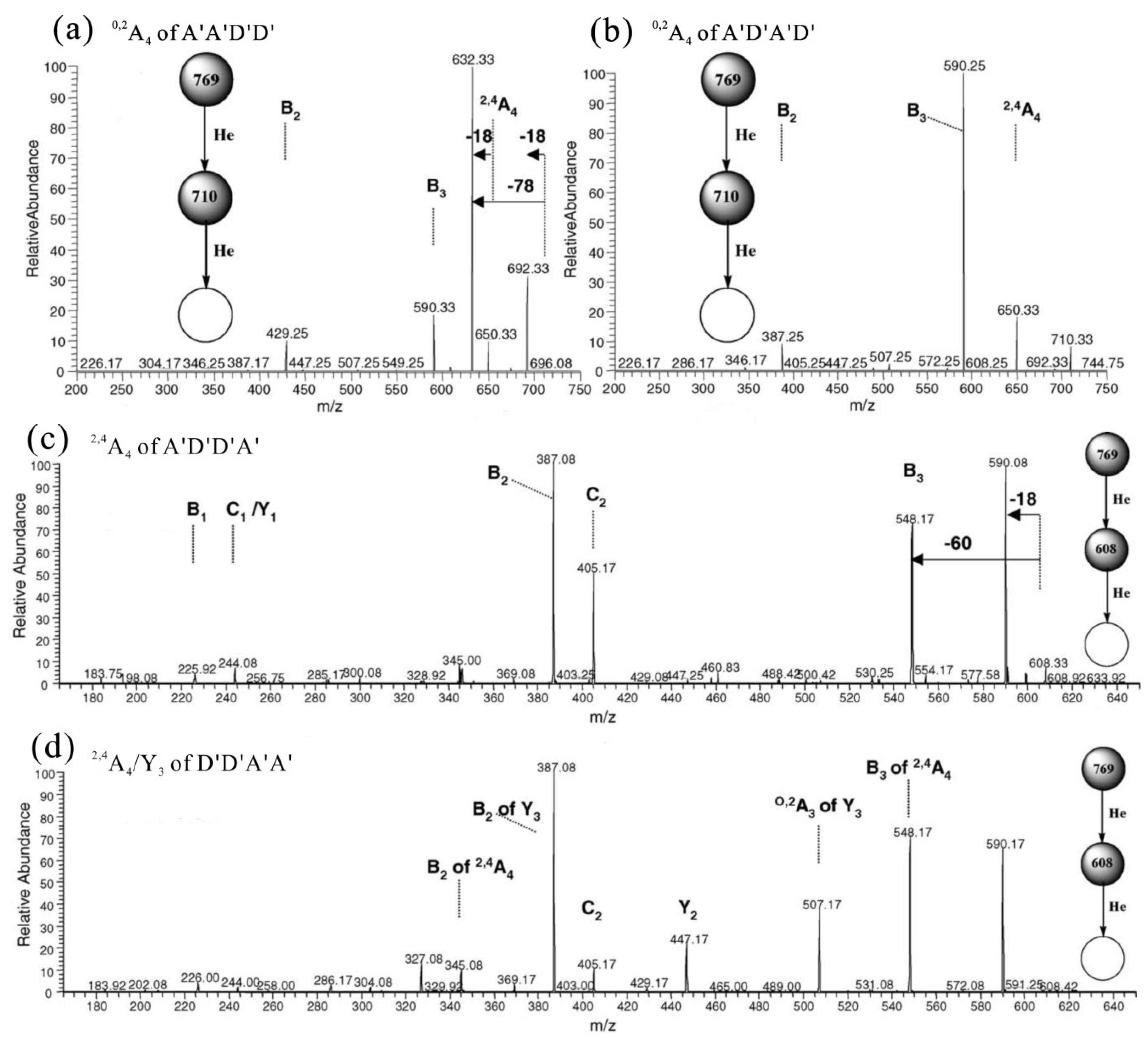{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligosaccharides | Assignment | Nominal Molecular Masses | ||
|---|---|---|---|---|
| Monosaccharide | Biose | Free Acid | Na Salt | |
| Neocarra-tetra-1S | A-G-A-G4S | β′-κ′ | 710 | 732 |
| Neocarra-hexa-3S | A-G4S-A-G4S-A-G4S | κ′-κ′-κ′ | 1176 | 1242 |
| Neocarra-hexa-4S | A-G4S-A2S-G4S-A-G4S | κ′-ι′-κ′ | 1256 | 1344 |
| Carra-tri-3S | G4S-A2S-G4S | ι-G4S | 726 | 792 |
| Carra-hepta-4S | G4S-A-G4S-A-G4S-A-G4S | κ-κ-κ-G4S | 1418 | 1506 |


2.2. MS of Agaro-Oligosaccharides
| Fractions | Found MNa+ | Assignment | Theoretical MNa+ | |
|---|---|---|---|---|
| DP | Sequences | |||
| A1 | 509.4 | 3 | G-A-G | 509.1 |
| A2 | 815.2 | 5 | G-A-G-A-G | 815.2 |
| A3 | 1121.3 | 7 | G-A-G-A-G-A-G | 1121.3 |
| A4 | 1427.5 | 9 | G-A-G-A-G-A-G-A-G | 1427.4 |
| A5 | 1733.4 | 11 | G-A-G-A-G-A-G-A-G-A-G | 1733.5 |

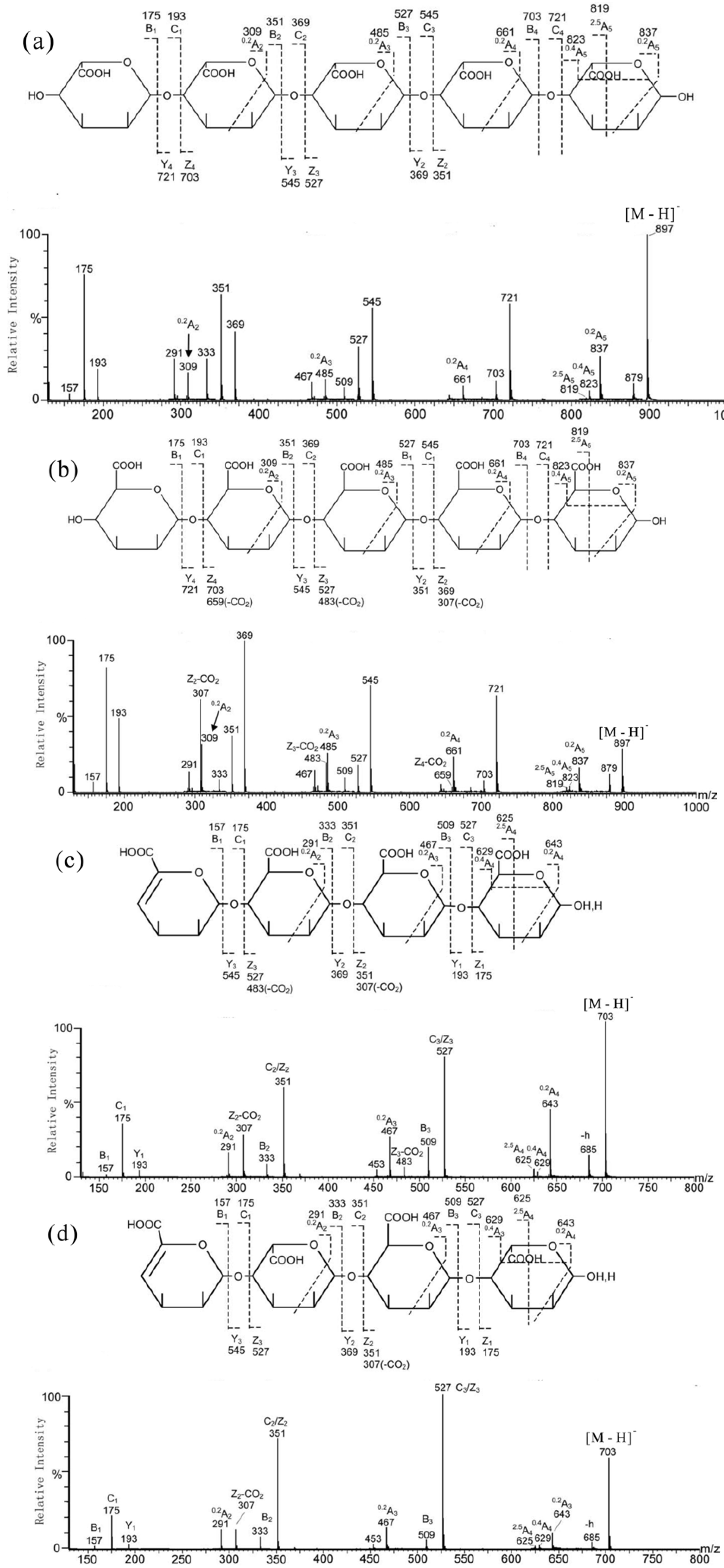
3. MS for Characterizing Alginate Oligosaccharides from Brown Algae

4. MS for Characterizing Sulfated Fucan Oligosaccharides from Echinoderm

5. MS for Characterizing Chitosan Oligosaccharides from Crustaceans


6. MS for Characterizing Glycosaminoglycans Oligosaccharides from Marine Animals
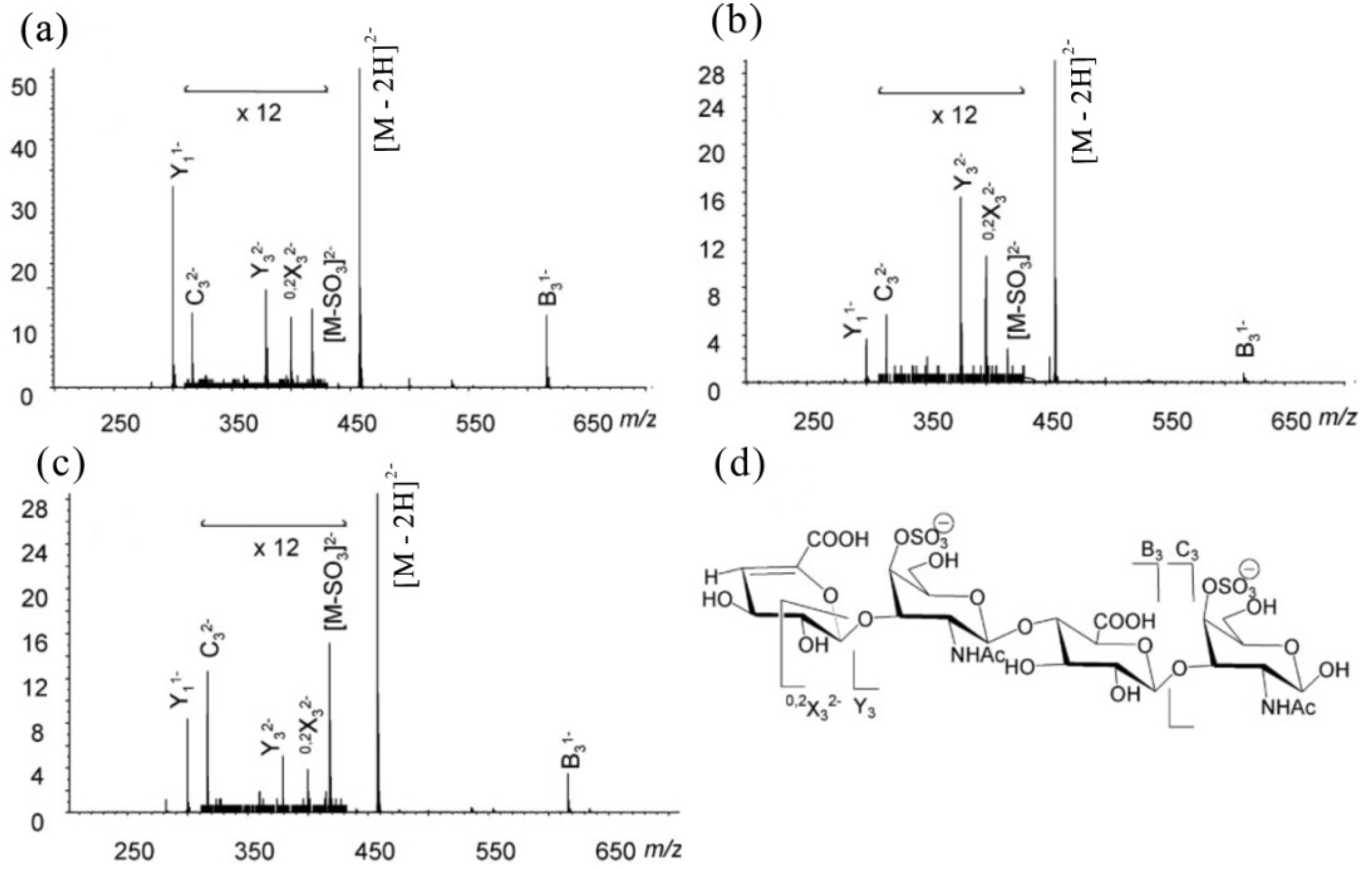
6.1. ESI-MS of CS/DS-Oligosaccharides
| Ion | CSA | CSB | CSC |
|---|---|---|---|
| Y11− | 33.91 ± 0.61 | 10.14 ± 0.56 | 19.88 ± 0.48 |
| B31− | 11.22 ± 0.33 | 2.92 ± 0.73 | 7.74 ± 0.52 |
| Y32− | 1.63 ± 0.27 | 3.18 ± 0.15 | 0.68 ± 0.05 |
| 0,2X32− | 0.06 ± 0.05 | 2.41 ± 0.31 | 0.44 ± 0.04 |
| C32− | 0.96 ± 0.03 | 0.78 ± 0.08 | 2.06 ± 0.18 |
| [M − SO3]2− | 0.89 ± 0.09 | 0.72 ± 0.64 | 2.23 ± 0.05 |

6.2. MALDI MS of CS/DS-Oligosaccharides

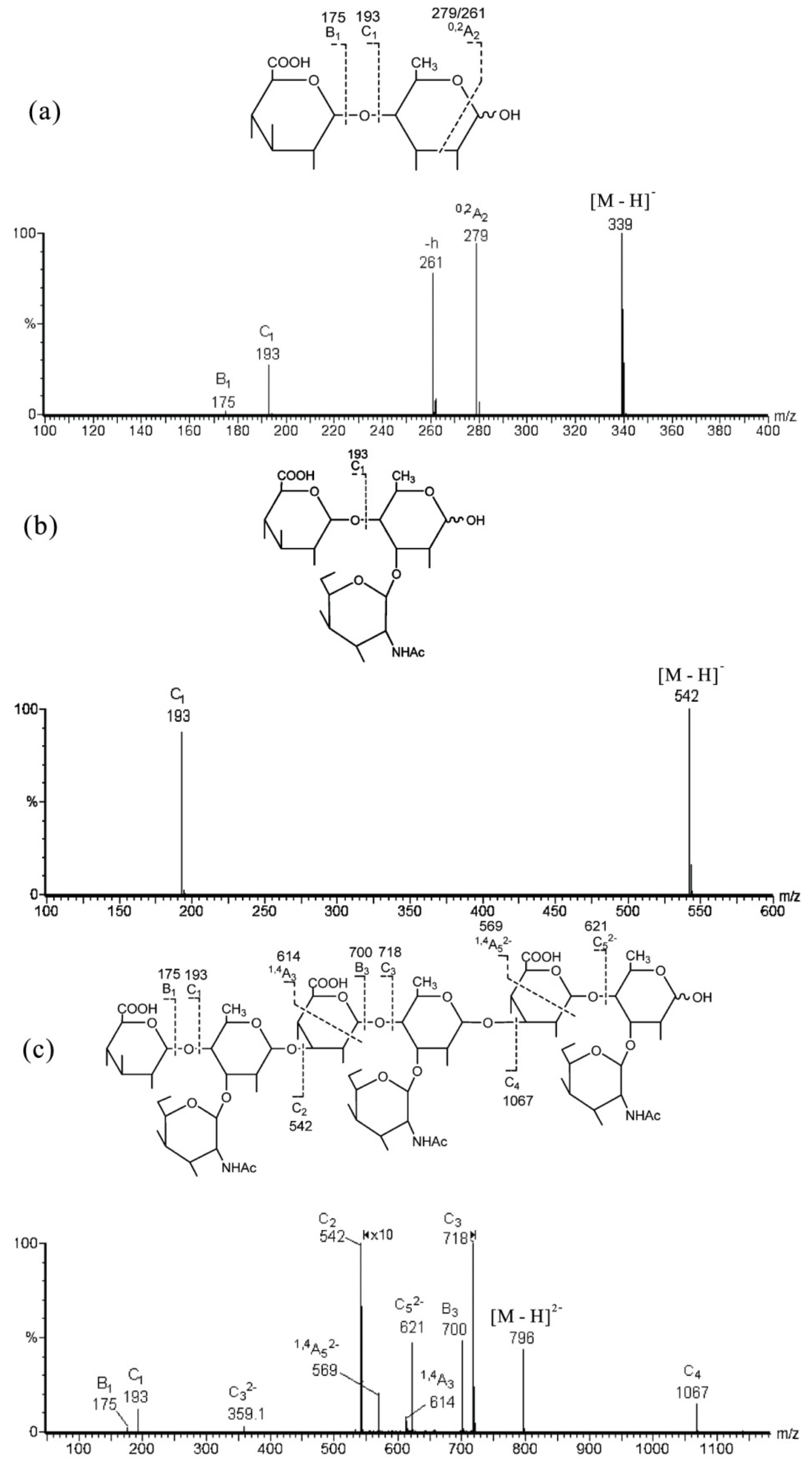
7. MS for Characterizing GAG-Like Oligosaccharides from Marine Invertebrates
| Fractions | Observed Ions | Assignments | ||
|---|---|---|---|---|
| [M − H]− | [M − 2H]2− | [M − 3H]3− | Compositions | |
| OFII | 796.0 | 530.3 | GlcA3·Fuc3·GalNAc3 | |
| OFIV | 542.2 | GlcA1·Fuc1·GalNAc1 | ||
| OFIVa | 339.1 | GlcA1·Fuc1 | ||

8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dewapriya, P.; Kim, S. Marine microorganisms: An emerging avenue in modern nutraceuticals and functional foods. Food Res. Int. 2014, 56, 115–125. [Google Scholar] [CrossRef]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.; Guan, H. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Q. Recent development in carbohydrate-based cancer vaccines. Curr. Opin. Chem. Biol. 2009, 13, 608–617. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S. Beneficial effects of marine algal compounds in cosmeceuticals. Mar. Drugs 2013, 11, 146–164. [Google Scholar] [CrossRef]
- Kim, S.; Ravichandran, Y.D.; Khan, S.B.; Kim, Y.T. Prospective of the cosmeceuticals derived from marine organisms. Biotechnol. Bioprocess Eng. 2008, 13, 511–523. [Google Scholar] [CrossRef]
- Silva, T.H.; Alves, A.; Ferreira, B.M.; Oliveira, J.M.; Reys, L.L.; Ferreira, R.; Sousa, R.A.; Silva, S.S.; Mano, J.F.; Reis, R.L. Materials of marine origin: A review on polymers and ceramics of biomedical interest. Int. Mater. Rev. 2012, 57, 276–306. [Google Scholar] [CrossRef]
- D’Ayala, G.G.; Malinconico, M.; Laurienzo, P. Marine derived polysaccharides for biomedical applications: Chemical modification approaches. Molecules 2008, 13, 2069–2106. [Google Scholar] [CrossRef]
- Silva, T.H.; Alves, A.; Popa, E.G.; Reys, L.L.; Gomes, M.E.; Sousa, R.A.; Silva, S.S.; Mano, J.F.; Reis, R.L. Marine algae sulfated polysaccharides for tissue engineering and drug delivery approaches. Biomatter 2012, 2, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Rodríguez-Amado, I.; Montemayor, M.I.; Fraguas, J.; González, M.D.P.; Murado, M.A. Chondroitin sulfate, hyaluronic acid and chitin/chitosan production using marine waste sources: Characteristics, applications and eco-friendly processes: A review. Mar. Drugs 2013, 11, 747–774. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Ciric, J.; Oostland, J.; de Vries, J.W.; Woortman, A.J.; Loos, K. Size exclusion chromatography with multi detection in combination with matrix-assisted laser desorption ionization-time-of-flight mass spectrometry as a tool for unraveling the mechanism of the enzymatic polymerization of polysaccharides. Anal. Chem. 2012, 84, 10463–10470. [Google Scholar] [CrossRef]
- El-Aneed, A.; Cohen, A.; Banoub, J. Mass spectrometry, review of the basics: Electrospray, MALDI, and commonly used mass analyzers. Appl. Spectrosc. Rev. 2009, 44, 210–230. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–409. [Google Scholar]
- Zaia, J. Mass spectrometry of oligosaccharides. Mass Spectrom. Rev. 2004, 23, 161–227. [Google Scholar] [CrossRef]
- Mischnick, P. Mass spectrometric characterization of oligo- and polysaccharides and their derivatives. Adv. Polym. Sci. 2012, 248, 105–174. [Google Scholar] [CrossRef]
- Stahl, M.; von Brocke, A.; Bayer, E. Mass spectrometry of oligosaccharides. J. Chromatogr. Libr. 2002, 66, 961–1042. [Google Scholar] [CrossRef]
- Han, L.; Costello, C.E. Mass spectrometry of glycans. Biochemistry 2013, 78, 710–720. [Google Scholar]
- Sisu, E.; Flangea, C.; Serb, A.; Zamfir, A.D. Modern developments in mass spectrometry of chondroitin and dermatan sulfate glycosaminoglycans. Amino Acids 2011, 41, 235–256. [Google Scholar] [CrossRef]
- Leymarie, N.; Zaia, J. Effective use of mass spectrometry for glycan and glycopeptide structural analysis. Anal. Chem. 2012, 84, 3040–3048. [Google Scholar] [CrossRef]
- Zaia, J. Glycosaminoglycan glycomics using mass spectrometry. Mol. Cell. Proteomics 2013, 12, 885–892. [Google Scholar] [CrossRef]
- Wuhrer, M. Glycomics using mass spectrometry. Glycoconj. J. 2013, 30, 11–22. [Google Scholar] [CrossRef]
- Oudenhove, L.; Devreese, B. A review on recent developments in mass spectrometry instrumentation and quantitative tools advancing bacterial proteomics. Appl. Microbiol. Biotechnol. 2013, 97, 4749–4762. [Google Scholar] [CrossRef]
- De Ruiter, G.A.; Rudolph, B. Carrageenan biotechnology. Trends Food Sci. Technol. 1997, 8, 389–395. [Google Scholar] [CrossRef]
- Usov, A.I. Structural analysis of red seaweed galactans of agar and carrageenan groups. Food Hydrocoll. 1998, 12, 301–308. [Google Scholar] [CrossRef]
- Duarte, M.E.R.; Noseda, D.G.; Noseda, M.D.; Tulio, S.; Pujol, C.A.; Damonte, E.B. Inhibitory effect of sulfated galactans from the marine alga Bostrychia montagnei on herpes simplex virus replication in vitro. Phytomedicine 2001, 8, 53–58. [Google Scholar] [CrossRef]
- Talarico, L.B.; Zibetti, R.G.M.; Faria, P.C.S.; Scolaro, L.A.; Duarte, M.E.R.; Noseda, M.D.; Pujol, C.A.; Damonte, E.B. Anti-herpes simplex virus activity of sulfated galactans from the red seaweeds Gymnogongrus griffithsiae and Cryptonemia crenulata. Int. J. Biol. Macromol. 2004, 34, 63–71. [Google Scholar] [CrossRef]
- Knutsen, S.H.; Myslabodski, D.E.; Larsen, B.; Usov, A.I. A modified system of nomenclature for red algal galactans. Bot. Mar. 1994, 37, 163–170. [Google Scholar]
- Gonçalves, A.G.; Ducatti, D.R.; Grindley, T.B.; Duarte, M.E.R.; Noseda, M.D. ESI-MS differential fragmentation of positional isomers of sulfated oligosaccharides derived from carrageenans and agarans. J. Am. Soc. Mass Spectrom. 2010, 21, 1404–1416. [Google Scholar] [CrossRef]
- Yu, G.; Zhao, X.; Yang, B.; Ren, S.; Guan, H.; Zhang, Y.; Lawson, A.M.; Chai, W. Sequence determination of sulfated carrageenan-derived oligosaccharides by high-sensitivity negative-ion electrospray tandem mass spectrometry. Anal. Chem. 2006, 78, 8499–8505. [Google Scholar] [CrossRef]
- Yang, B.; Yu, G.; Zhao, X.; Jiao, G.; Ren, S.; Chai, W. Mechanism of mild acid hydrolysis of galactan polysaccharides with highly ordered disaccharide repeats leading to a complete series of exclusively odd-numbered oligosaccharides. FEBS J. 2009, 276, 2125–2137. [Google Scholar] [CrossRef]
- Hu, Y.; Yu, G.; Zhao, X.; Wang, Y.; Sun, X.; Jiao, G.; Zhao, X.; Chai, W. Structural characterization of natural ideal 6-O-sulfated agarose from red alga Gloiopeltis furcata. Carbohydr. Polym. 2012, 89, 883–889. [Google Scholar] [CrossRef]
- Lahaye, M. Developments on gelling algal galactans, their structure and physico-chemistry. J. Appl. Phycol. 2001, 13, 173–184. [Google Scholar] [CrossRef]
- Aarstad, O.A.; Tondervik, A.; Sletta, H.; Skjak-Braek, G. Alginate sequencing: An analysis of block distribution in alginates using specific alginate degrading enzymes. Biomacromolecules 2012, 13, 106–116. [Google Scholar] [CrossRef]
- Zhang, Z.; Yu, G.; Zhao, X.; Liu, H.; Guan, H.; Lawson, A.M.; Chai, W. Sequence analysis of alginate-derived oligosaccharides by negative-ion electrospray tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2006, 17, 621–630. [Google Scholar] [CrossRef]
- Pomin, V.H.; Mourao, P.A.S. Structure, biology, evolution, and medical importance of sulfated fucans and galactans. Glycobiology 2008, 18, 1016–1027. [Google Scholar] [CrossRef]
- Kariya, Y.; Mulloy, B.; Imai, K.; Tominaga, A.; Kaneko, T.; Asari, A.; Suzuki, K.; Masuda, H.; Kyogashima, M.; Ishii, T. Isolation and partial characterization of fucan sulfates from the body wall of sea cucumber Stichopus japonicus and their ability to inhibit osteoclastogenesis. Carbohydr. Res. 2004, 339, 1339–1346. [Google Scholar] [CrossRef]
- Mulloy, B.; Ribeiro, A.; Alves, A.; Vieira, R.P.; Mourao, P.A.S. Sulfated fucans from echinoderms have a regular tetrasaccharide repeating unit defined by specific patterns of sulfation at the 0–2 and 0–4 positions. J. Biol. Chem. 1994, 269, 22113–22123. [Google Scholar]
- Berteau, O.; Mulloy, B. Sulfated fucans, fresh perspectives: Structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 2003, 13, 29R–40R. [Google Scholar] [CrossRef]
- Zaia, J.; Costello, C.E. Tandem mass spectrometry of sulfated heparin-like glycosaminoglycan oligosaccharides. Anal. Chem. 2003, 75, 2445–2455. [Google Scholar] [CrossRef]
- Chi, L.; Amster, J.; Linhardt, R.J. Mass spectrometry for the analysis of highly charged sulfated carbohydrates. Curr. Anal. Chem. 2005, 1, 223–240. [Google Scholar] [CrossRef]
- Chen, S.; Hu, Y.; Ye, X.; Li, G.; Yu, G.; Xue, C.; Chai, W. Sequence determination and anticoagulant and antithrombotic activities of a novel sulfated fucan isolated from the sea cucumber Isostichopus badionotus. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 989–1000. [Google Scholar]
- Hund, R.; Cherif, C.; Hund, H.; Aibibu, D.; Toskas, G.; Richter, K.; Richter, T. Chitin, chitosan, bio-fibers-preparation, properties and use. Chem. Fibers Int. 2013, 63, 211–213. [Google Scholar]
- Ahlafi, H.; Moussout, H.; Boukhlifi, F.; Echetna, M.; Bennani, M.N.; Slimane, S.M. Kinetics of N-deacetylation of chitin extracted from shrimp shells collected from coastal area of Morocco. Mediterr. J. Chem. 2013, 2, 503–513. [Google Scholar] [CrossRef]
- Younes, I.; Ghorbel-Bellaaj, O.; Nasri, R.; Chaabouni, M.; Rinaudo, M.; Nasri, M. Chitin and chitosan preparation from shrimp shells using optimized enzymatic deproteinization. Process Biochem. (Oxf. UK) 2012, 47, 2032–2039. [Google Scholar] [CrossRef]
- Kumar, M.N.V.R. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Jayakumar, R.; Prabaharan, M.; Sudheesh Kumar, P.T.; Nair, S.V.; Tamura, H. Biomaterials based on chitin and chitosan in wound dressing applications. Biotechnol. Adv. 2011, 29, 322–337. [Google Scholar] [CrossRef]
- Kurita, K. Chitin and chitosan: Functional biopolymers from marine crustaceans. Mar. Biotechnol. 2006, 8, 203–226. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A.; Muzzarelli, C. Chitosan chemistry: Relevance to the biomedical sciences. Adv. Polym. Sci. 2005, 186, 151–209. [Google Scholar] [CrossRef]
- Vijayakrishnan, B.; Issaree, A.; Corilo, Y.E.; Ferreira, C.R.; Eberlin, M.N.; Peter, M.G. MSn of the six isomers of (GlcN)2(GlcNAc)2 aminoglucan tetrasaccharides (diacetylchitotetraoses): Rules of fragmentation for the sodiated molecules and application to sequence analysis of hetero-chitooligosaccharides. Carbohydr. Polym. 2011, 84, 713–726. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K.; Ozbek, S. Evolution of glycosaminoglycans: Comparative biochemical study. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef]
- Deepa, S.S.; Yamada, S.; Fukui, S.; Sugahara, K. Structural determination of novel sulfated octasaccharides isolated from chondroitin sulfate of shark cartilage and their application for characterizing monoclonal antibody epitopes. Glycobiology 2007, 17, 631–645. [Google Scholar] [CrossRef]
- Field, I.C.; Meekan, M.G.; Buckworth, R.C.; Bradshaw, C.J. Susceptibility of sharks, rays and chimaeras to global extinction. Adv. Mar. Biol. 2009, 56, 275–363. [Google Scholar] [CrossRef]
- Zhou, X.; Weiser, P.; Pan, J.; Qian, Y.; Lu, H.; Zhang, L. Chondroitin sulfate and abnormal contact system in rheumatoid arthritis. Prog. Mol. Biol. Transl. Sci. 2010, 93, 423–442. [Google Scholar] [CrossRef]
- Wang, H.; Kong, X.; He, J.; Ye, T.; Jang, J. An immunochemical study of chondroitin sulfate (CS) in cirrhotic liver tissue and hepatocellular carcinoma. Shanghai Yixue 1995, 18, 68–70. [Google Scholar]
- Makeudom, A.; Kulpawaropas, S.; Montreekachon, P.; Khongkhunthian, S.; Sastraruji, T.; Pothacharoen, P.; Kongtawelert, P.; Krisanaprakornkit, S. Positive correlations between hCAP18/LL-37 and chondroitin sulphate levels in chronic periodontitis. J. Clin. Periodontol. 2014, 41, 252–261. [Google Scholar] [CrossRef]
- Zaia, J.; Miller, M.J.; Seymour, J.L.; Costello, C.E. The role of mobile protons in negative ion CID of oligosaccharides. J. Am. Soc. Mass Spectrom. 2007, 18, 952–960. [Google Scholar] [CrossRef]
- Zaia, J.; McClellan, J.E.; Costello, C.E. Tandem mass spectrometric determination of the 4S/6S sulfation sequence in chondroitin sulfate oligosaccharides. Anal. Chem. 2001, 73, 6030–6039. [Google Scholar] [CrossRef]
- Miller, M.J.; Costello, C.E.; Malmstrom, A.; Zaia, J. A tandem mass spectrometric approach to determination of chondroitin/dermatan sulfate oligosaccharide glycoforms. Glycobiology 2006, 16, 502–513. [Google Scholar] [CrossRef]
- Laremore, T.N.; Zhang, F.; Linhardt, R.J. Ionic liquid matrix for direct UV-MALDI-TOF-MS analysis of dermatan sulfate and chondroitin sulfate oligosaccharides. Anal. Chem. 2007, 79, 1604–1610. [Google Scholar] [CrossRef]
- Laremore, T.N.; Murugesan, S.; Park, T.; Avci, F.Y.; Zagorevski, D.V.; Linhardt, R.J. Matrix-assisted laser desorption/ionization mass spectrometric analysis of uncomplexed highly sulfated oligosaccharides using ionic liquid matrices. Anal. Chem. 2006, 78, 1774–1779. [Google Scholar]
- Tissot, B.R.R.; Gasiunas, N.; Powell, A.K.; Ahmed, Y.; Zhi, Z.; Haslam, S.M.; Morris, H.R.; Turnbull, J.E.; Gallagher, J.T.; Dell, A. Towards GAG glycomics: Analysis of highly sulfated heparins by MALDI-TOF mass spectrometry. Glycobiology 2007, 17, 972–982. [Google Scholar] [CrossRef]
- Laremore, T.N.; Linhardt, R.J. Improved matrix-assisted laser desorption/ionization mass spectrometric detection of glycosaminoglycan disaccharides as cesium salts. Rapid Commun. Mass Spectrom. 2007, 21, 1315–1320. [Google Scholar] [CrossRef]
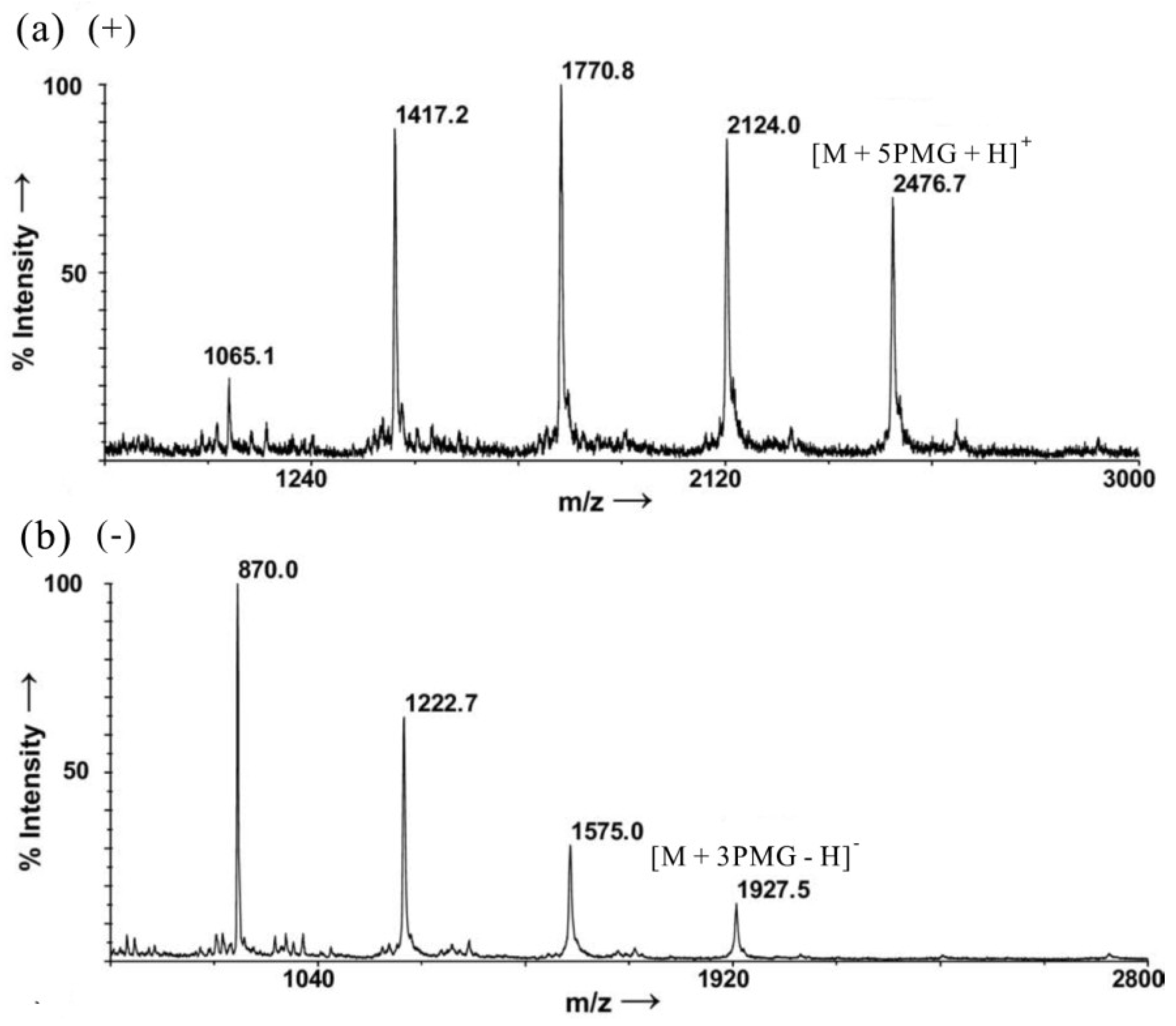
- Ohara, K.; Jacquinet, J.C.; Jouanneau, D.; Helbert, W.; Smietana, M.; Vasseur, J.J. Matrix-assisted laser desorption/ionization mass spectrometric analysis of polysulfated-derived oligosaccharides using pyrenemethylguanidine. J. Am. Soc. Mass Spectrom. 2009, 20, 131–137. [Google Scholar]
- Santos, J.C.; Mesquita, J.M.F.; Belmiro, C.L.R.; da Silveira, C.B.M.; Viskov, C.; Mourier, P.A.; Pavao, M.S.G. Isolation and characterization of a heparin with low antithrombin activity from the body of Styela plicata (Chordata-Tunicata). Distinct effects on venous and arterial models of thrombosis. Thromb. Res. 2007, 121, 213–223. [Google Scholar] [CrossRef]
- Gomes, A.M.; Kozlowski, E.O.; Pomin, V.H.; de Barros, C.M.; Zaganeli, J.L.; Pavao, M.S.G. Unique extracellular matrix heparan sulfate from the bivalve Nodipecten nodosus (Linnaeus, 1758) safely inhibits arterial thrombosis after photochemically induced endothelial lesion. J. Biol. Chem. 2010, 285, 7312–7323. [Google Scholar]
- Borsig, L.; Wang, L.; Cavalcante, M.C.M.; Cardilo-Reis, L.; Ferreira, P.L.; Mourao, P.A.S.; Esko, J.D.; Pavao, M.S.G. Selectin blocking activity of a fucosylated chondroitin sulfate glycosaminoglycan from sea cucumber. Effect on tumor metastasis and neutrophil recruitment. J. Biol. Chem. 2007, 282, 14984–14991. [Google Scholar] [CrossRef]
- Belmiro, C.L.R.; Castelo-Branco, M.T.L.; Melim, L.M.C.; Schanaider, A.; Elia, C.; Madi, K.; Pavao, M.S.G.; de Souza, H.S.P. Unfractionated heparin and new heparin analogues from ascidians (chordate-tunicate) ameliorate colitis in rats. J. Biol. Chem. 2009, 284, 11267–11278. [Google Scholar] [CrossRef]
- Bao, X.; Pavao, M.S.G.; Dos, S.J.C.; Sugahara, K. A functional dermatan sulfate epitope containing iduronate(2-O-sulfate)alpha1–3GalNAc(6-O-sulfate) disaccharide in the mouse brain: Demonstration using a novel monoclonal antibody raised against dermatan sulfate of ascidian Ascidia nigra. J. Biol. Chem. 2005, 280, 23184–23193. [Google Scholar] [CrossRef]
- Hikino, M.; Mikami, T.; Faissner, A.; Vilela-Silva, A.E.S.; Pavao, M.S.G.; Sugahara, K. Oversulfated dermatan sulfate exhibits neurite outgrowth-promoting activity toward embryonic mouse hippocampal neurons: Implications of dermatan sulfate in neuritogenesis in the brain. J. Biol. Chem. 2003, 278, 43744–43754. [Google Scholar]
- Nandini, C.D.; Mikami, T.; Ohta, M.; Itoh, N.; Akiyama-Nambu, F.; Sugahara, K. Structural and functional characterization of oversulfated chondroitin sulfate/dermatan sulfate hybrid chains from the notochord of hagfish: Neuritogenic and binding activities for growth factors and neurotrophic factors. J. Biol. Chem. 2004, 279, 50799–50809. [Google Scholar]
- Shetty, A.K.; Kobayashi, T.; Mizumoto, S.; Narumi, M.; Kudo, Y.; Yamada, S.; Sugahara, K. Isolation and characterization of a novel chondroitin sulfate from squid liver integument rich in N-acetylgalactosamine(4,6-disulfate) and glucuronate(3-sulfate) residues. Carbohydr. Res. 2009, 344, 1526–1532. [Google Scholar] [CrossRef]
- Tamura, J.; Arima, K.; Imazu, A.; Tsutsumishita, N.; Fujita, H.; Yamane, M.; Matsumi, Y. Sulfation patterns and the amounts of chondroitin sulfate in the diamond squid, Thysanoteuthis rhombus. Biosci. Biotechnol. Biochem. 2009, 73, 1387–1391. [Google Scholar]
- Li, G.; Chen, S.; Wang, Y.; Xue, Y.; Chang, Y.; Li, Z.; Wang, J.; Xue, C. A novel glycosaminoglycan-like polysaccharide from abalone Haliotis discus hannai Ino: Purification, structure identification and anticoagulant activity. Int. J. Biol. Macromol. 2011, 49, 1160–1166. [Google Scholar] [CrossRef]
- Chai, W.; Piskarev, V.; Lawson, A.M. Negative-ion electrospray mass spectrometry of neutral underivatized oligosaccharides. Anal. Chem. 2001, 73, 651–657. [Google Scholar] [CrossRef]
- Chen, S.; Xu, J.; Xue, C.; Dong, P.; Sheng, W.; Yu, G.; Chai, W. Sequence determination of a non-sulfated glycosaminoglycan-like polysaccharide from melanin-free ink of the squid Ommastrephes bartrami by negative-ion electrospray tandem mass spectrometry and NMR spectroscopy. Glycoconj. J. 2008, 25, 481–492. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lang, Y.; Zhao, X.; Liu, L.; Yu, G. Applications of Mass Spectrometry to Structural Analysis of Marine Oligosaccharides. Mar. Drugs 2014, 12, 4005-4030. https://doi.org/10.3390/md12074005
Lang Y, Zhao X, Liu L, Yu G. Applications of Mass Spectrometry to Structural Analysis of Marine Oligosaccharides. Marine Drugs. 2014; 12(7):4005-4030. https://doi.org/10.3390/md12074005
Chicago/Turabian StyleLang, Yinzhi, Xia Zhao, Lili Liu, and Guangli Yu. 2014. "Applications of Mass Spectrometry to Structural Analysis of Marine Oligosaccharides" Marine Drugs 12, no. 7: 4005-4030. https://doi.org/10.3390/md12074005
APA StyleLang, Y., Zhao, X., Liu, L., & Yu, G. (2014). Applications of Mass Spectrometry to Structural Analysis of Marine Oligosaccharides. Marine Drugs, 12(7), 4005-4030. https://doi.org/10.3390/md12074005