Understanding Nanocalcification: A Role Suggested for Crystal Ghosts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. “Maturation” of Crystals
1.2. Early Inorganic Structures


1.3. Organic-Inorganic Relationships
1.4. Decalcification Procedures

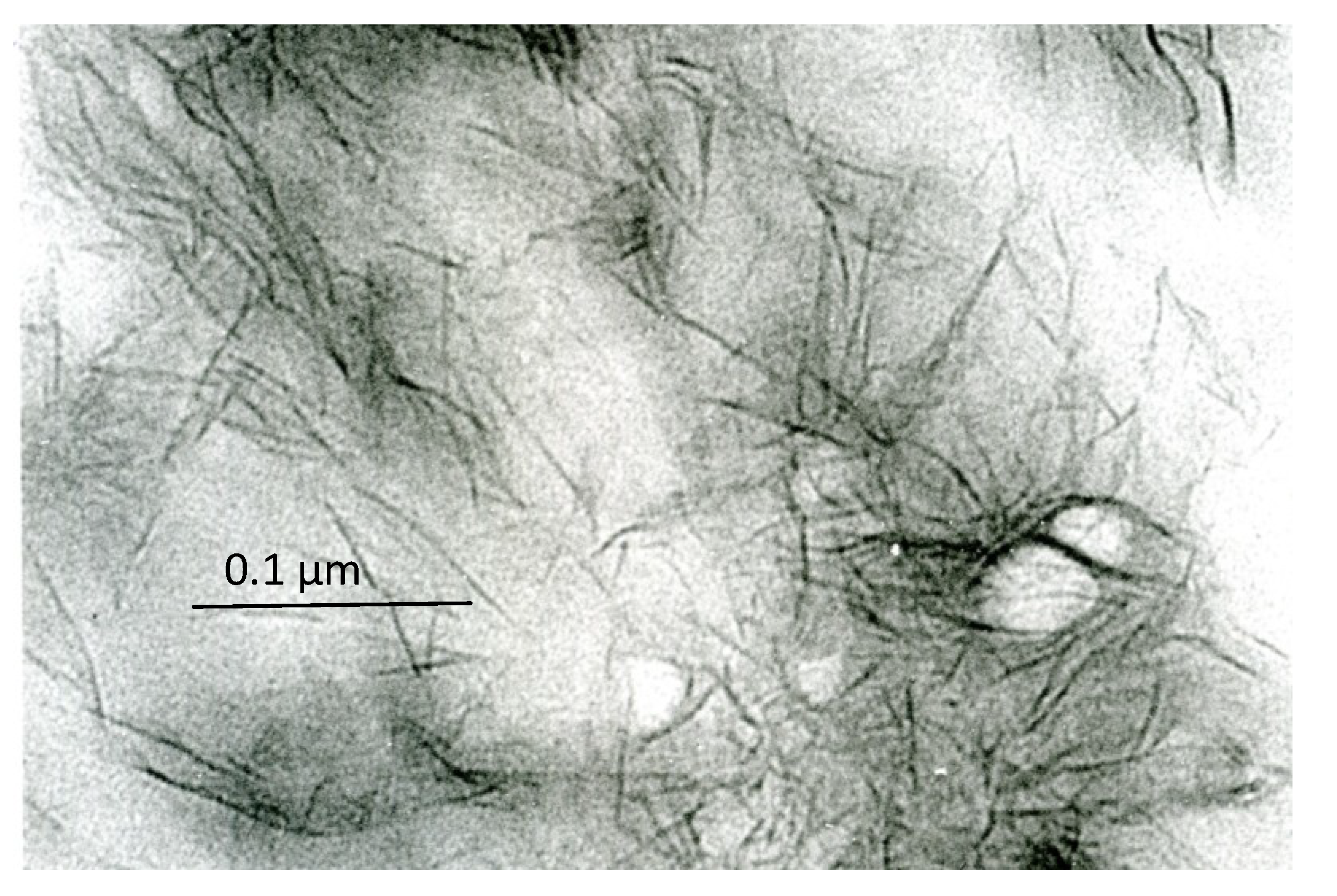
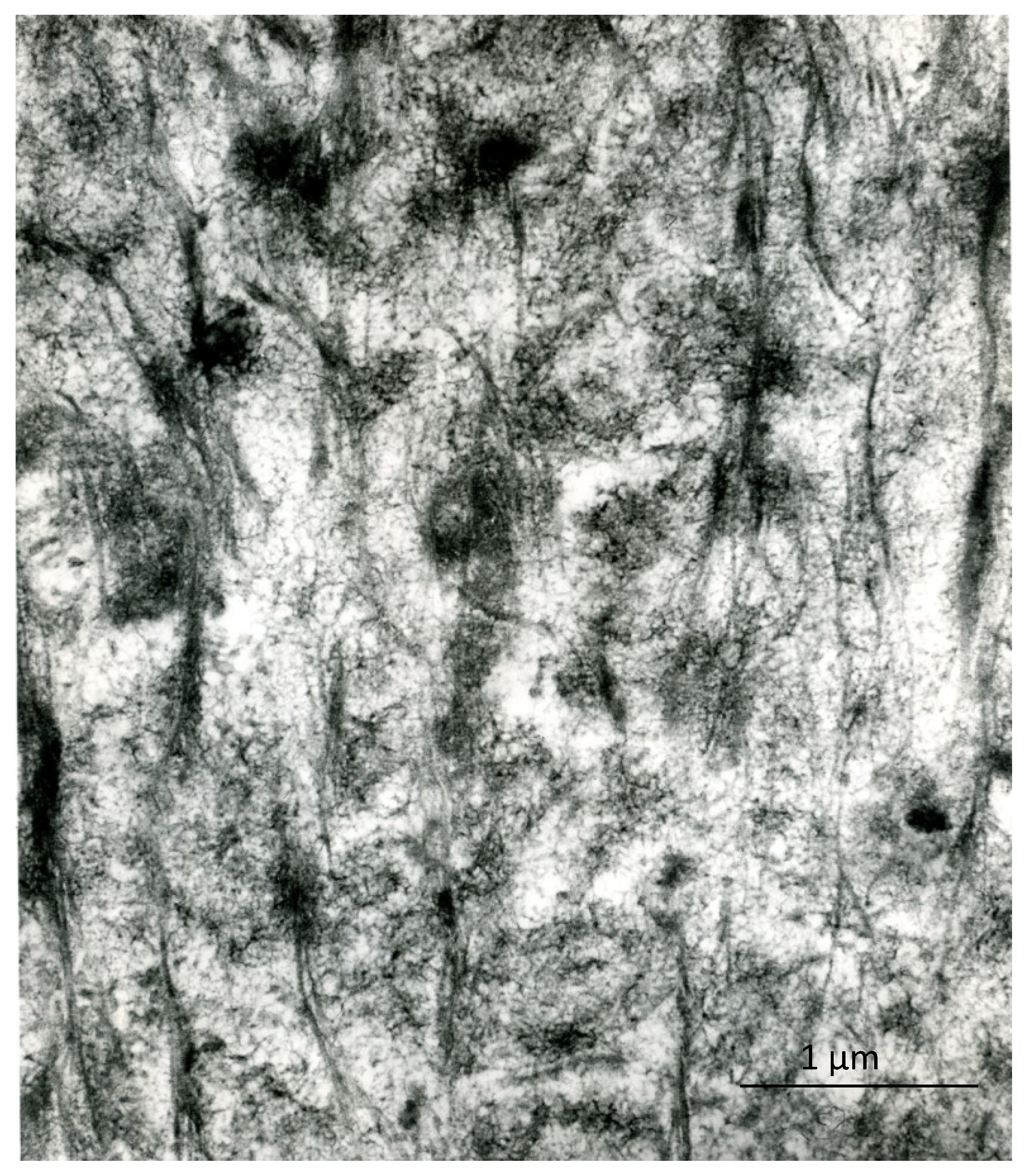
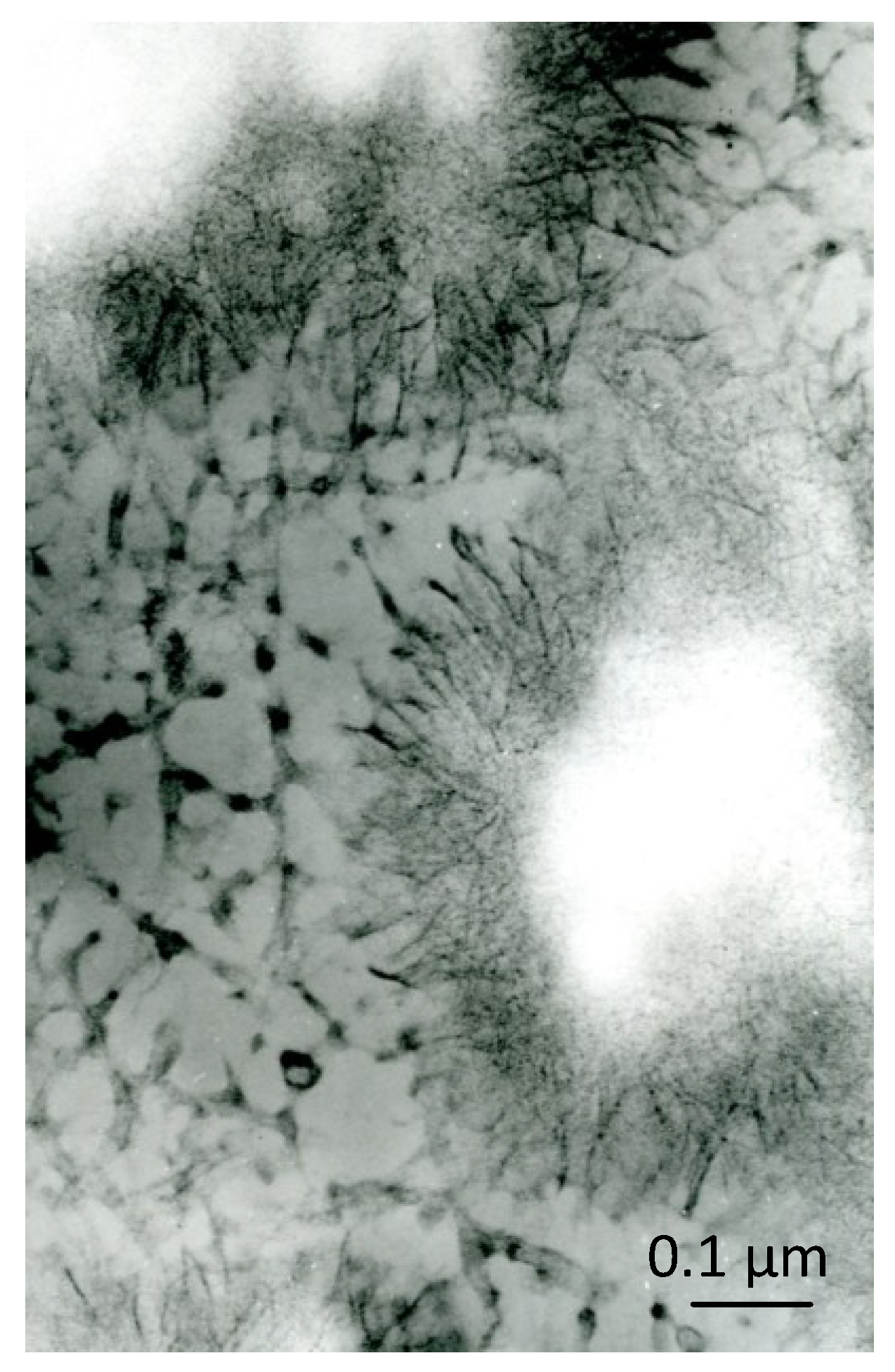
1.5. Crystal Ghosts


1.6. Supposed Function of Crystal Ghosts

2. Conclusions
Conflicts of Interest
References
- De Jong, W.F. La substance minérale dans les os. Rec. Trav. Chim. 1926, 45, 445–446. [Google Scholar] [CrossRef]
- Roseberry, H.H.; Hastings, A.B.; Morse, J.K. X-ray analysis of bone and teeth. J. Biol. Chem. 1931, 90, 335–407. [Google Scholar]
- Ascenzi, A.; Bonucci, E. A quantitative investigation of the birefringence of the osteon. Acta Anat. 1961, 44, 236–262. [Google Scholar] [CrossRef]
- Ascenzi, A.; Bonucci, E.; Bocciarelli, S.D. An electron microscope study of osteon calcification. J. Ultrastruct. Res. 1965, 12, 287–303. [Google Scholar] [CrossRef]
- Ascenzi, A.; Bonucci, E.; Bocciarelli, S.D. An electron microscope study on primary periosteal bone. J. Ultrastruct. Res. 1967, 18, 605–618. [Google Scholar] [CrossRef]
- Bonucci, E. Biological calcification. In Normal and Pathological Processes in the Early Stages; Springer-Verlag: Berlin, Heidelberg, Germany, 2007. [Google Scholar]
- Arnott, H.J.; Pautard, F.G.E. Osteoblast function and fine structure. Israel J. Med. Sci. 1967, 3, 657–670. [Google Scholar]
- Posner, A.S.; Harper, R.A.; Muller, S.A.; Menczel, J. Age changes in the crystal chemistry of bone apatite. Ann. N. Y. Acad. Sci. 1965, 131, 737–742. [Google Scholar] [CrossRef]
- Bonar, L.C.; Roufosse, A.H.; Sabine, W.K.; Grynpas, M.D.; Glimcher, M.J. X-ray diffraction studies of the crystallinity of bone mineral in newly synthesized and density fractionated bone. Calcif. Tissue Int. 1983, 35, 202–209. [Google Scholar] [CrossRef]
- Burnell, J.M.; Teubner, E.J.; Miller, A.G. Normal maturation changes in bone matrix, mineral, and crystal size in the rat. Calcif. Tissue Int. 1980, 31, 13–19. [Google Scholar] [CrossRef]
- Legros, R.; Balmain, N.; Bonel, G. Age-related changes in mineral of rat and bovine cortical bone. Calcif. Tissue Int. 1987, 41, 137–144. [Google Scholar] [CrossRef]
- Smith, C.B.; Smith, D.A. An X-ray diffraction investigation of age-related changes in the crystal structure of bone apatite. Calcif. Tissue Res. 1976, 22, 219–226. [Google Scholar] [CrossRef]
- Wu, Y.; Ackerman, J.L.; Kim, H.-M.; Rey, C.; Barroug, A.; Glimcher, M.J. Nuclear magnetic resonance spin-spin relaxation of the crystals of bone, dental enamel, and synthetic hydroxyapatites. J. Bone Miner. Res. 2002, 17, 472–480. [Google Scholar] [CrossRef]
- Matsushima, N.; Hikichi, K. Age changes in the crystallinity of bone mineral and in the disorder of its crystal. Biochim. Biophys. Acta 1989, 992, 155–159. [Google Scholar] [CrossRef]
- Wheeler, E.J.; Lewis, D. An X-ray study of the paracrystalline nature of bone apatite. Calcif. Tissue Res. 1977, 24, 243–248. [Google Scholar] [CrossRef]
- Arnold, S.; Plate, U.; Wiesmann, H.P.; Stratmann, U.; Kohl, H.; Höhling, H.J. Quantitative analyses of the biomineralization of different hard tissues. J. Microsc. 2001, 202, 488–494. [Google Scholar] [CrossRef]
- Arnold, S.; Plate, U.; Wiesmann, H.P.; Stratmann, U.; Kohl, H.; Höhling, H.J. Quantitative electron spectroscopic diffraction analyses of the crystal formation in dentine. J. Microsc. 1999, 195, 58–63. [Google Scholar] [CrossRef]
- Landis, W.J.; Glimcher, M.J. Electron diffraction and electron probe microanalysis of the mineral phase of bone tissue prepared by anhydrous techniques. J. Ultrastruct. Res. 1978, 63, 188–223. [Google Scholar] [CrossRef]
- Wergedal, J.E.; Baylink, D.J. Electron microprobe measurements of bone mineralization rate in vivo. Am. J. Physiol. 1974, 226, 345–352. [Google Scholar]
- Nanci, A. TenCate’s Oral Histology: Development, Structure, and Function, 6th ed.; Mosby: St. Louis, MO, USA, 2003. [Google Scholar]
- Landis, W.J.; Burke, G.Y.; Neuringer, J.R.; Paine, M.C.; Nanci, A.; Bai, P.; Warshawsky, H. Earliest enamel deposits of the rat incisor examined by electron microscopy, electron diffraction, and electron probe microanalysis. Anat. Record 1988, 220, 233–238. [Google Scholar] [CrossRef]
- Beniash, E.; Metzler, R.A.; Lam, R.S.; Gilbert, P.U. Transient amorphous calcium phosphate in forming enamel. J. Struct. Biol. 2009, 166, 133–143. [Google Scholar] [CrossRef]
- Diekwisch, T.G.H. Subunit compartments of secretory stage enamel matrix. Connect. Tissue Res. 1998, 38, 101–111. [Google Scholar] [CrossRef]
- Simmer, J.P.; Richardson, A.S.; Hu, Y.Y.; Smith, C.E.; Ching-Chun Hu, J. A post-classical theory of enamel biomineralization and why we need one. Int. J. Oral. Sci. 2012, 4, 129–134. [Google Scholar] [CrossRef]
- Mahamid, J.; Sharir, A.; Addadi, L.; Weiner, S. Amorphous calcium phosphate is a major component of the forming fin bones of zebrafish: Indications for an amorphous precursor phase. Proc. Natl. Acad. Sci. USA 2008, 105, 12748–12753. [Google Scholar] [CrossRef]
- Glimcher, M.J. The nature of the mineral component of bone and the mechanism of calcification. In Disorders of Bone and Mineral Metabolism; Coe, F.L., Favus, M.J., Eds.; Raven Press: New York, NY, USA, 1992; pp. 265–286. [Google Scholar]
- Bonucci, E. Bone mineralization. Front. Biosci. 2012, 17, 100–128. [Google Scholar] [CrossRef]
- Bonucci, E. The mineralization of bone and its analogies with other hard tissues. In Advanced Topics in Crystal Growth; Ferreira, S.O., Ed.; InTech: Rijeka, Croazia, 2013; pp. 145–184. [Google Scholar]
- Bonucci, E. Fine structure of early cartilage calcification. J. Ultrastruct. Res. 1967, 20, 33–50. [Google Scholar] [CrossRef]
- Bonucci, E.; Reurink, J. The fine structure of decalcified cartilage and bone: A comparison between decalcification procedures performed before and after embedding. Calcif. Tissue Res. 1978, 25, 179–190. [Google Scholar] [CrossRef]
- Shepard, N. Role of proteoglycans in calcification. In Calcification in Biological Systems; Bonucci, E., Ed.; CRC Press: Boca Raton, FL, USA, 1992; pp. 41–58. [Google Scholar]
- Bonucci, E. Crystal ghosts and biological mineralization: Fancy spectres in an old castle, or neglected structures worthy of belief? J. Bone Miner. Metab. 2002, 20, 249–265. [Google Scholar] [CrossRef]
- Bonucci, E.; Gomez, S. Cartilage calcification. In Advanced Topics in Biomineralization; Seto, J., Ed.; InTech: Rijeka, Croazia, 2012; pp. 85–110. [Google Scholar]
- Prostak, K.S.; Lees, S. Visualization of crystal-matrix structure. In situ demineralization of mineralized turkey leg tendon and bone. Calcif. Tissue Int. 1996, 59, 474–479. [Google Scholar] [CrossRef]
- Goldberg, M.; Noblot, M.M.; Septier, D. Effets de deux méthodes de démineralisation sur la préservation des glycoprotéines et des protéoglycanes dans les dentines intercanaliculaires et péricanaliculaires chez le cheval. J. Biol. Buccale 1980, 8, 315–330. [Google Scholar]
- Hayashi, Y. Ultrastructural demonstration of the carbohydrate in developing mantle dentine with soybean agglutinin-gold complexes. J. Electron. Microsc. 1988, 37, 150–154. [Google Scholar]
- Bishop, M.A.; Warshawsky, H. Electron microscopic studies on the potential loss of crystallites from routinely processed sections of young enamel in the rat incisor. Anat. Record 1982, 202, 177–186. [Google Scholar] [CrossRef]
- Hayashi, Y.; Bianco, P.; Shimokawa, H.; Termine, J.D.; Bonucci, E. Organic-inorganic relationships, and immunohistochemical localization of amelogenins and enamelins in developing enamel. Basic Appl. Histochem. 1986, 30, 291–299. [Google Scholar]
- Kallenbach, E. Crystal-associated matrix components in rat incisor enamel. An electron-microscopic study. Cell Tissue Res. 1986, 246, 455–461. [Google Scholar] [CrossRef]
- Nanci, A.; Bai, P.; Warshawsky, H. The effect of osmium postfixation and uranyl and lead staining on the ultrastructure of young enamel in the rat incisor. Anat. Record 1983, 207, 1–16. [Google Scholar] [CrossRef]
- Smales, F.C. Structural subunit in prisms of immature rat enamel. Nature 1975, 258, 772–774. [Google Scholar] [CrossRef]
- Davis, W.L.; Jones, R.G.; Hagler, H.K. An electron microscopic histochemical and analytical X-ray microprobe study of calcification in Bruch’s membrane from human eyes. J. Histochem. Cytochem. 1981, 29, 601–608. [Google Scholar] [CrossRef]
- Kuhar, K.J.; Eisenmann, D.R. Fluoride-induced mineralization within vacuoles in maturative ameloblasts of the rat. Anat. Record 1978, 191, 91–102. [Google Scholar] [CrossRef]
- Boothroyd, B. The problem of demineralisation in thin sections of fully calcified bone. J. Cell Biol. 1964, 20, 165–173. [Google Scholar]
- Dong, W.; Warshawsky, H. Failure to demonstrate a protein coat on enamel crystallites by morphological means. Archs Oral Biol. 1995, 40, 321–330. [Google Scholar] [CrossRef]
- Warshawsky, H. External shape of enamel crystals. Scann. Microsc. 1987, 1, 1913–1923. [Google Scholar]
- Warshawsky, H. Organization of crystals in enamel. Anat. Record 1989, 224, 242–262. [Google Scholar] [CrossRef]
- Albeck, S.; Addadi, L.; Weiner, S. Regulation of calcite crystal morphology by intracrystalline acidic proteins and glycoproteins. Connect. Tissue Res. 1996, 35, 365–370. [Google Scholar] [CrossRef]
- Berman, A.; Addadi, L.; Kvick, Å.; Leiserowitz, L.; Nelson, M.; Weiner, S. Intercalation of sea urchin proteins in calcite: Study of a crystalline composite material. Science 1990, 250, 664–667. [Google Scholar]
- Berman, A.; Addadi, L.; Weiner, S. Interactions of sea-urchin skeleton macromolecules with growing calcite crystals—A study of intracrystalline proteins. Nature 1988, 331, 546–548. [Google Scholar] [CrossRef]
- Pokroy, B.; Fitch, A.N.; Lee, P.L.; Quintana, J.P.; Caspi, E.N.; Zolotoyabko, E. Anisotropic lattice distortions in the mollusk-made aragonite: A widespread phenomenon. J. Struct. Biol. 2006, 153, 145–150. [Google Scholar] [CrossRef]
- Tong, H.; Hu, J.; Ma, W.; Zhong, G.; Yao, S.; Cao, N. In situ analysis of the organic framework in the prismatic layer of mollusc shell. Biomaterials 2002, 23, 2593–2598. [Google Scholar] [CrossRef]
- Bonucci, E.; Silvestrini, G.; Di Grezia, R. Histochemical properties of the “crystal ghosts” of calcifying epiphyseal cartilage. Connect. Tissue Res. 1989, 22, 43–50. [Google Scholar]
- Bonucci, E.; Silvestrini, G. Immunohistochemical investigation on the presence of chondroitin sulfate in calcification nodules of epiphyseal cartilage. Eur. J. Histochem. 1992, 36, 407–422. [Google Scholar]
- Appleton, J. Ultrastructural observations on the inorganic/organic relationships in early cartilage calcification. Calcif. Tissue Res. 1971, 7, 307–317. [Google Scholar] [CrossRef]
- Davis, W.L.; Jones, R.G.; Knight, J.P.; Hagler, H.K. Cartilage calcification: An ultrastructural, histochemical, and analytical X-ray microprobe study of the zone of calcification in normal avian epiphyseal growth plate. J. Histochem. Cytochem. 1982, 30, 221–234. [Google Scholar] [CrossRef]
- Chardin, H.; Septier, D.; Goldberg, M. Visualization of glycosaminoglycans in rat incisor predentin and dentin with cetylpyridinium chloride-glutaraldehyde as fixative. J. Histochem. Cytochem. 1990, 38, 885–894. [Google Scholar] [CrossRef]
- Sauren, Y.M.H.F.; Mieremet, R.H.P.; Groot, C.G.; Scherft, J.P. An electron microscopical study on the presence of proteoglycans in the calcified bone matrix by use of cuprolinic blue. Bone 1989, 10, 287–294. [Google Scholar] [CrossRef]
- Hayashi, Y. Ultrastructural demonstration of the carbohydrate in developing rat enamel using soybean agglutinin-gold complexes. Arch. Oral Biol. 1989, 34, 517–522. [Google Scholar] [CrossRef]
- Gomez, S.; Lopez-Cepero, J.M.; Silvestrini, G.; Mocetti, P.; Bonucci, E. Matrix vesicles and focal proteoglycan aggregates are the nucleation sites revealed by the lanthanum incubation method: A correlated study on the hypertrophic zone of the rat epiphyseal cartilage. Calcif. Tissue Int. 1996, 58, 273–282. [Google Scholar] [CrossRef]
- Butler, W.T. Matrix macromolecules of bone and dentin. Collagen Rel. Res. 1984, 4, 297–307. [Google Scholar] [CrossRef]
- Deutsch, D.; Catalano-Sherman, J.; Dafni, L.; David, S.; Palmon, A. Enamel matrix proteins and ameloblast biology. Connect. Tissue Res. 1995, 32, 97–107. [Google Scholar] [CrossRef]
- Gorski, J.P. Biomineralization of bone: A fresh view of the roles of non-collagenous proteins. Front. Biosci. 2011, 17, 2598–2621. [Google Scholar] [CrossRef]
- Gotliv, B.A.; Addadi, L.; Weiner, S. Mollusk shell acidic proteins: In search of individual functions. Chembiochem 2003, 4, 522–529. [Google Scholar] [CrossRef]
- Marin, F.; Narayanappa, P.; Motreuil, S. Acidic shell proteins of the mediterranean fan mussel Pinna nobilis. Progr. Mol. Subcell Biol. 2011, 52, 353–395. [Google Scholar] [CrossRef]
- Rahman, M.A.; Oomori, T. In vitro regulation of CaCO3 crystal growth by the highly acidic proteins of calcitic sclerites in soft coral, Sinularia polydactyla. Connect. Tissue Res. 2010, 50, 285–293. [Google Scholar] [CrossRef]
- Veis, A. Mineral-matrix interactions in bone and dentin. J. Bone Miner. Res. 1993, 8 (suppl. 2), S493–S497. [Google Scholar] [CrossRef]
- Weiner, S. Aspartic acid-rich proteins: Major components of the soluble organic matrix of mollusk shells. Calcif. Tissue Int. 1979, 29, 163–167. [Google Scholar] [CrossRef]
- Mann, S. Biomineralization: Principles and Concepts. In Bioinorganic Materials Chemistry, 1st ed.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Addadi, L.; Weiner, S. Interactions between acidic proteins and crystals: Stereochemical requirements in biomineralization. Proc. Natl. Acad. Sci. USA 1985, 82, 4110–4114. [Google Scholar] [CrossRef]
- Aizenberg, J.; Hanson, J.; Ilan, M.; Leiserowitz, L.; Koetzle, T.F.; Addadi, L.; Weiner, S. Morphogenesis of calcitic sponge spicules: A role for specialized proteins interacting with growing crystals. FASEB J. 1995, 9, 262–268. [Google Scholar]
- Fang, P.A.; Margolis, H.C.; Conway, J.F.; Simmer, J.P.; Beniash, E. CryoTEM study of effects of phosphorylation on the hierarchical assembly of porcine amelogenin and its regulation of mineralization in vitro. J. Struct. Biol. 2013, 183, 250–257. [Google Scholar] [CrossRef]
- Kawska, A.; Hochrein, O.; Brickmann, J.; Kniep, D.; Zahn, D. The nucleation mechanism of fluorapatite-collagen composites: Ion association and motif control by collagen proteins. Angew. Chem. Int. Ed. 2008, 47, 4982–4985. [Google Scholar] [CrossRef]
- Wang, Y.; Azaïs, T.; Robin, M.; Vallée, A.; Catania, C.; Legriel, P.; Pehau-Arnaudet, G.; Babonneau, F.; Giraud-Guille, M.M.; Nassif, N. The predominant role of collagen in the nucleation, growth, structure and orientation of bone apatite. Nat. Mat. 2012, 11, 724–733. [Google Scholar] [CrossRef]
- Moradian-Oldak, J. Protein-mediated enamel mineralization. Front. Biosci. 2012, 17, 1996–2023. [Google Scholar] [CrossRef]
- Glimcher, M.J.; Brickley-Parsons, D.; Levine, P.T. Studies of enamel proteins during maturation. Calcif. Tissue Res. 1977, 24, 259–270. [Google Scholar] [CrossRef]
- Pugliarello, M.C.; Vittur, F.; de Bernard, B.; Bonucci, E.; Ascenzi, A. Chemical modifications in osteones during calcification. Calcif. Tissue Res. 1970, 5, 108–114. [Google Scholar] [CrossRef]
- Lohmander, S.; Hjerpe, A. Proteoglycans of mineralizing rib and epiphyseal cartilage. Biochim. Biophys. Acta 1975, 404, 93–109. [Google Scholar] [CrossRef]
- Althoff, J.; Quint, P.; Krefting, E.R.; Höhling, H.J. Morphological studies on the epiphyseal growth plate combined with biochemical and X-ray microprobe analyses. Histochemistry 1982, 74, 541–552. [Google Scholar] [CrossRef]
- Campo, R.D.; Dziewiatkowski, D.D. Turnover of the organic matrix of cartilage and bone as visualized by autoradiography. J. Cell Biol. 1963, 18, 19–29. [Google Scholar] [CrossRef]
- Campo, R.D.; Romano, J.E. Changes in cartilage proteoglycans associated with calcification. Calcif. Tissue Int. 1986, 39, 175–184. [Google Scholar] [CrossRef]
- Hirschman, A.; Dziewiatkowski, D.D. Protein-polysaccharide loss during endochondral ossification: Immunochemical evidence. Science 1966, 154, 393–395. [Google Scholar]
- Matukas, V.J.; Krikos, G.A. Evidence for changes in protein polysaccharide associated with the onset of calcification in cartilage. J. Cell Biol. 1968, 39, 43–48. [Google Scholar] [CrossRef]
- Aizenberg, J.; Lambert, G.; Addadi, L.; Weiner, S. Stabilization of amorphous calcium carbonate by specialized macromolecules in biological and synthetic precipitates. Adv. Mater. 1996, 8, 222–226. [Google Scholar] [CrossRef]
- Bentov, S.; Weil, S.; Glazer, L.; Sagi, A.; Berman, A. Stabilization of amorphous calcium carbonate by phosphate rich organic matrix proteins and by single phosphoamino acids. J. Struct. Biol. 2010, 171, 207–215. [Google Scholar] [CrossRef]
- Gajjeraman, S.; Narayanan, K.; Hao, J.; George, A. Matrix macromolecules in hard tissues controls the nucleation and hierarchical assembly of hydroxyapatite. J. Biol. Chem. 2007, 282, 1193–1204. [Google Scholar] [CrossRef]
- Politi, Y.; Arad, T.; Klein, E.; Weiner, S.; Addadi, L. Sea urchin spine calcite forms via a transient amorphous calcium carbonate phase. Science 2004, 306, 1161–1164. [Google Scholar] [CrossRef]
- Weiner, S.; Levi-Kalisman, Y.; Raz, S.; Addadi, L. Biologically formed amorphous calcium carbonate. Connect. Tissue Res. 2003, 44 (suppl. 1), 214–218. [Google Scholar] [CrossRef]
- Zhang, G.; Xu, J. From colloidal nanoparticles to a single crystal: New insights into the formation of nacre’s aragonite tablets. J. Struct. Biol. 2013, 182, 36–43. [Google Scholar] [CrossRef]
- Dey, A.; Bomans, P.H.; Müller, F.A.; Will, J.; Frederik, P.M.; de With, G.; Sommerdijk, N.A. The role of prenucleation clusters in surface-induced calcium phosphate crystallization. Nat. Mater. 2010, 9, 1010–1014. [Google Scholar] [CrossRef]
- Kwak, S.Y.; Green, S.; Wiedemann-Bidlack, F.B.; Beniash, E.; Yamakoshi, Y.; Simmer, J.P.; Margolis, H.C. Regulation of calcium phosphate formation by amelogenins under physiological conditions. Eur. J. Oral Sci. 2011, 119 (suppl. 1), 103–111. [Google Scholar] [CrossRef] [Green Version]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bonucci, E. Understanding Nanocalcification: A Role Suggested for Crystal Ghosts. Mar. Drugs 2014, 12, 4231-4246. https://doi.org/10.3390/md12074231
Bonucci E. Understanding Nanocalcification: A Role Suggested for Crystal Ghosts. Marine Drugs. 2014; 12(7):4231-4246. https://doi.org/10.3390/md12074231
Chicago/Turabian StyleBonucci, Ermanno. 2014. "Understanding Nanocalcification: A Role Suggested for Crystal Ghosts" Marine Drugs 12, no. 7: 4231-4246. https://doi.org/10.3390/md12074231
APA StyleBonucci, E. (2014). Understanding Nanocalcification: A Role Suggested for Crystal Ghosts. Marine Drugs, 12(7), 4231-4246. https://doi.org/10.3390/md12074231