Endotoxin Structures in the Psychrophiles Psychromonas marina and Psychrobacter cryohalolentis Contain Distinctive Acyl Features

Abstract
:
1. Introduction
2. Results
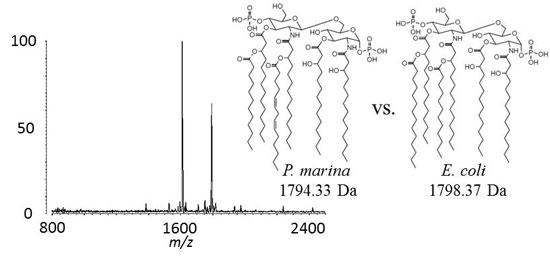
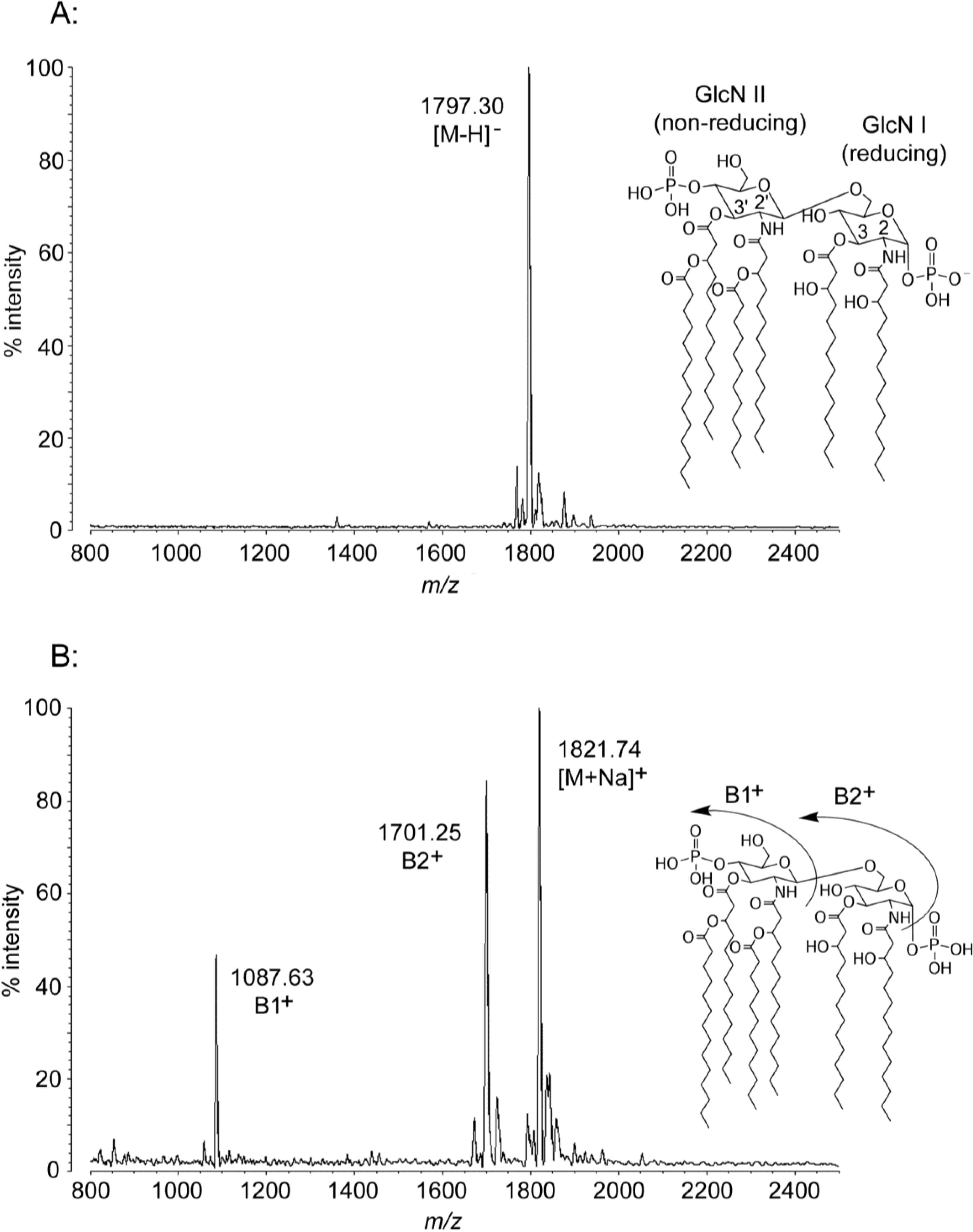
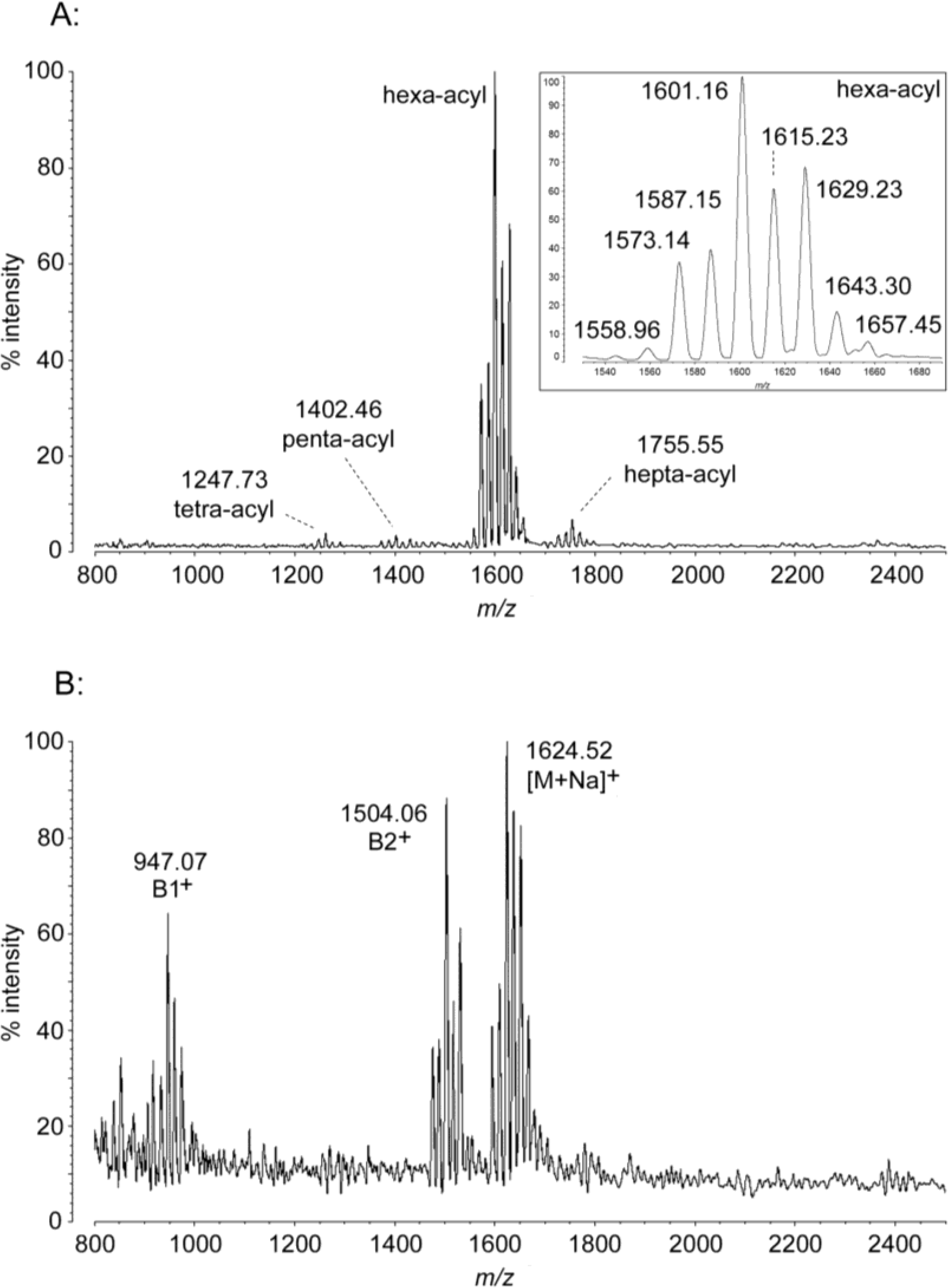
2.1. MALDI-TOF MS of E. coli Lipid A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | [M − H]− Predicted Mass (Da) | [M − H]− Observed m/z | B1+ Predicted Mass (Da) | B1+ Observed m/z | B2+ Predicted Mass (Da) | B2+ Observed m/z | [M + Na]+ Predicted Mass (Da) | [M + Na]+ Observed m/z |
|---|---|---|---|---|---|---|---|---|
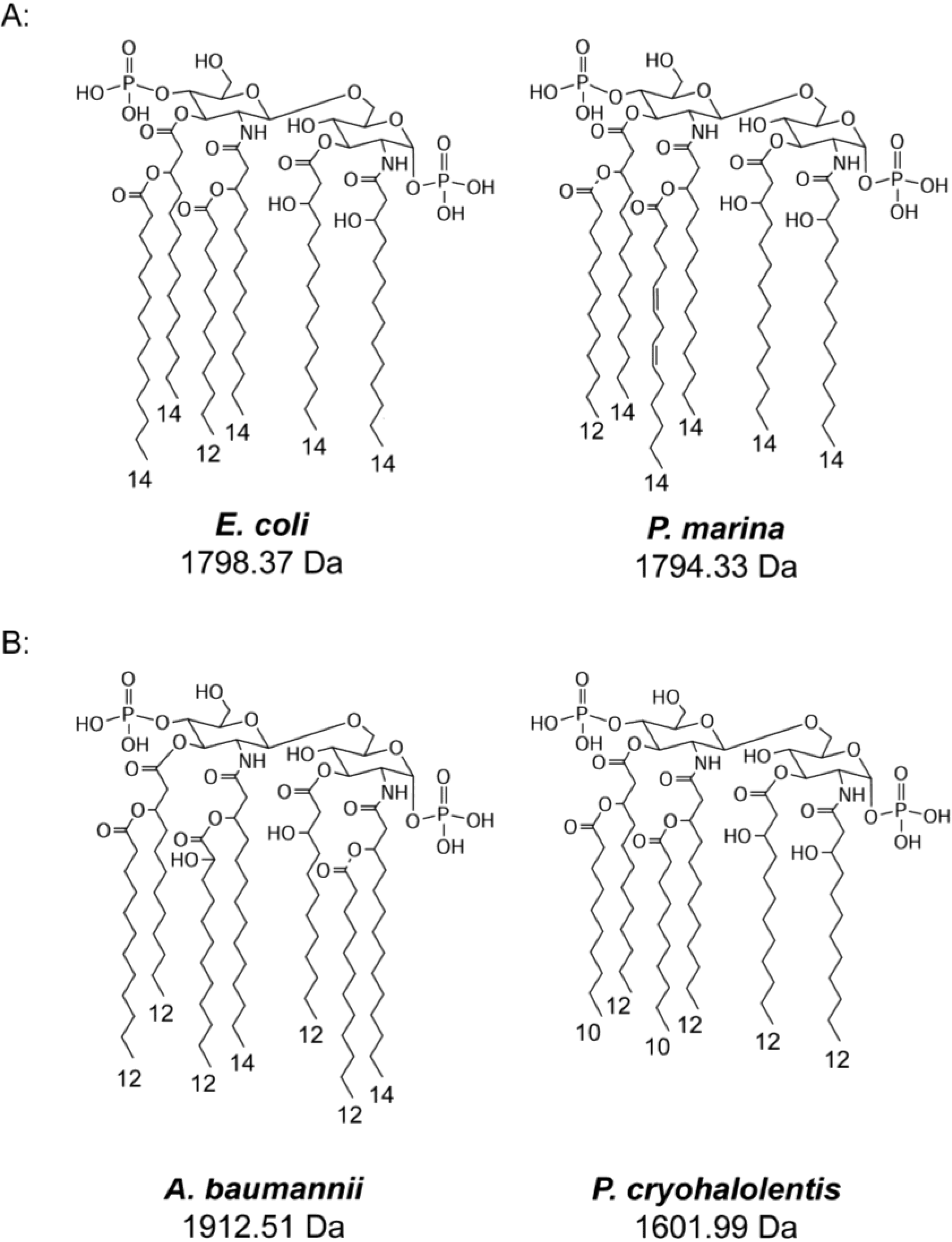
| E. coli hexa-acyl | 1797.36 | 1797.30 | 1087.51 | 1087.63 | 1701.38 | 1701.25 | 1821.35 | 1821.74 |
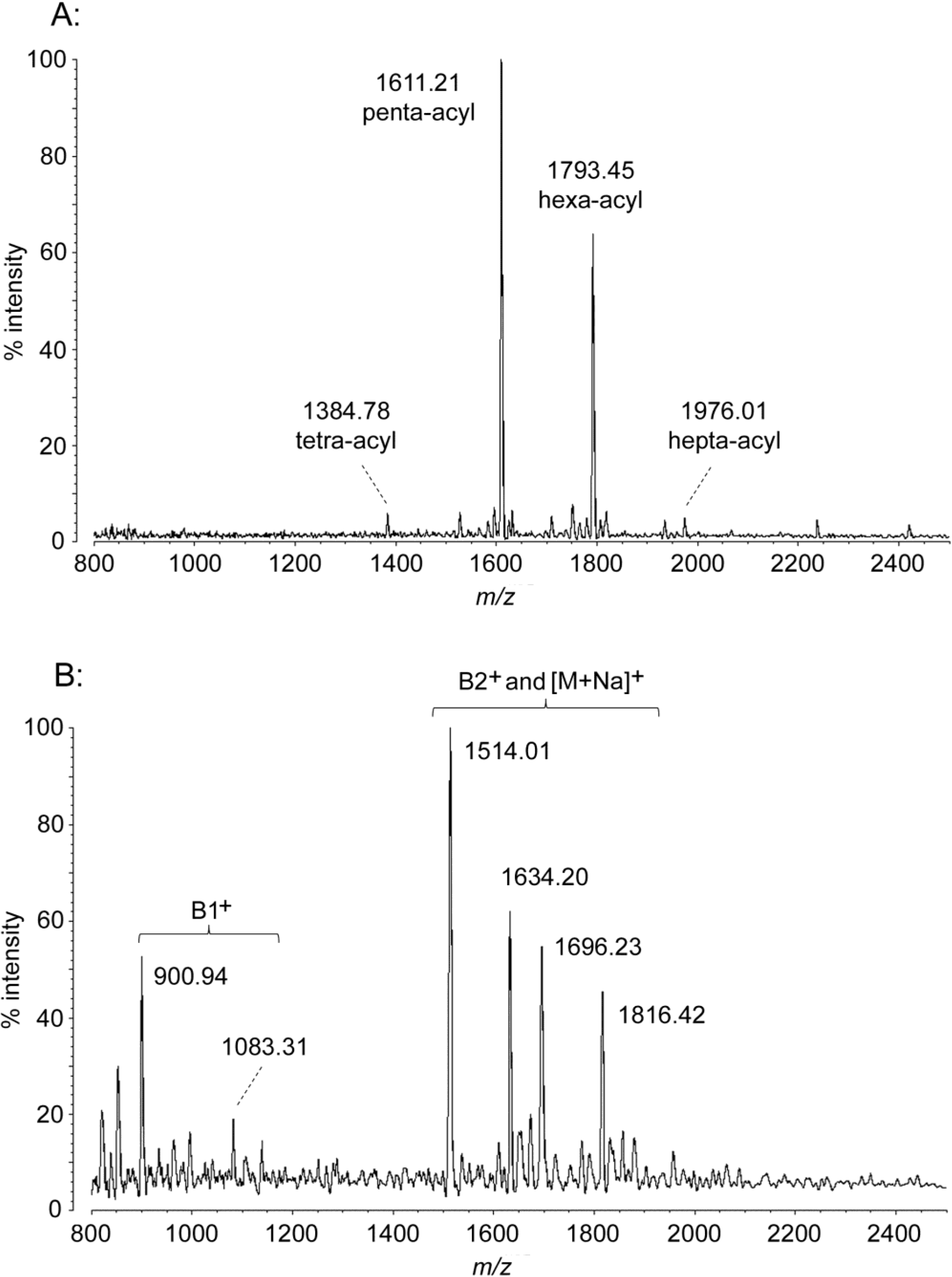
| P. marina penta-acyl | 1611.02 | 1611.21 | 901.21 | 900.94 | 1515.04 | 1514.01 | 1635.02 | 1634.20 |
| hexa-acyl | 1793.33 | 1793.45 | 1083.48 | 1083.31 | 1697.35 | 1696.23 | 1817.32 | 1816.42 |
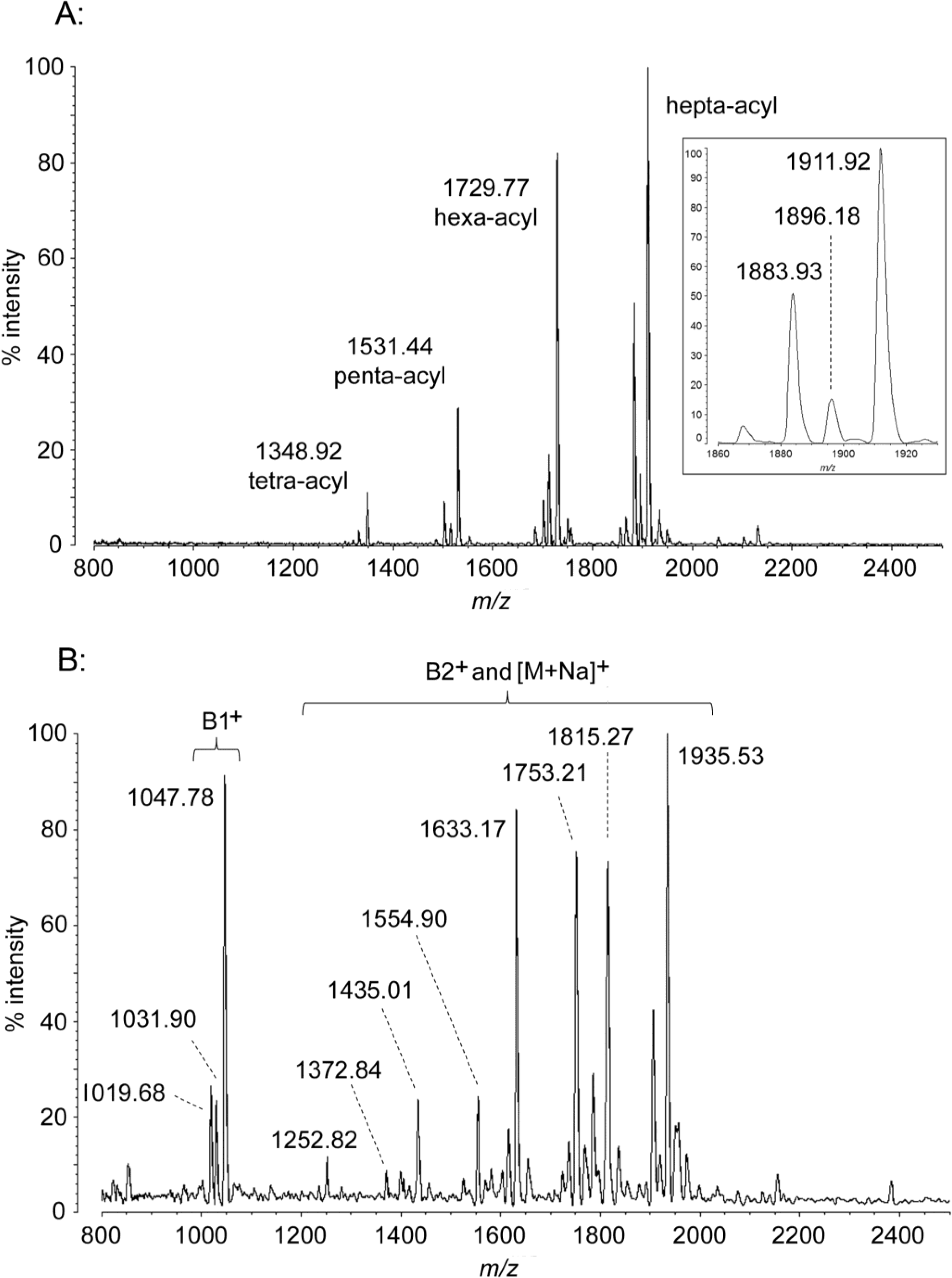
| A. baumannii hexa-acyl | 1730.20 | 1729.77 | 1047.40 | 1047.78 | 1633.22 | 1633.17 | 1753.19 | 1753.21 |
| hexa-acyl − (OH) | 1714.21 | 1713.80 | 1031.41 | 1031.90 | 1617.22 | 1616.93 | 1737.20 | 1737.31 |
| hexa-acyl − (CH2)2 | 1702.15 | 1701.71 | 1019.35 | 1019.68 | 1605.16 | 1604.72 | 1725.14 | 1724.97 |
| hepta-acyl | 1911.50 | 1911.92 | 1047.40 | 1047.78 | 1815.52 | 1815.27 | 1935.50 | 1935.53 |
| hepta-acyl − (OH) | 1895.50 | 1896.18 | 1031.41 | 1031.90 | 1799.52 | -- | 1919.50 | 1920.34 |
| hepta-acyl − (CH2)2 | 1883.45 | 1883.93 | 1019.35 | 1019.68 | 1787.47 | 1787.30 | 1907.44 | 1907.36 |
| P. cryohalolentis − (CH2)3 | 1558.91 | 1558.96 | 905.17 | 907.11 | 1462.93 | 1461.39 | 1582.90 | 1581.95 |
| −(CH2)2 | 1572.93 | 1573.14 | 919.19 | 919.09 | 1476.95 | 1476.26 | 1596.93 | 1596.22 |
| −CH2 | 1586.96 | 1587.15 | 933.22 | 934.13 | 1490.98 | 1489.93 | 1610.96 | 1610.55 |
| hexa-acyl | 1600.98 | 1601.16 | 947.25 | 947.07 | 1505.01 | 1504.06 | 1624.98 | 1624.52 |
| +CH2 | 1615.01 | 1615.23 | 961.27 | 961.36 | 1519.03 | 1517.92 | 1639.01 | 1638.86 |
| +(CH2)2 | 1629.04 | 1629.23 | 975.30 | 975.01 | 1533.06 | 1532.13 | 1653.04 | 1652.71 |
| +(CH2)3 | 1643.06 | 1643.30 | 989.33 | 990.63 | 1547.09 | 1546.00 | 1667.06 | 1667.18 |
| +(CH2)4 | 1657.09 | 1657.13 | 1003.35 | 1003.68 | 1561.11 | -- | 1681.09 | 1680.35 |
2.2. MALDI-TOF MS of P. marina Lipid A


2.3. MALDI-TOF MS of A. baumannii Lipid A
2.4. MALDI-TOF MS of P. cryohalolentis Lipid A

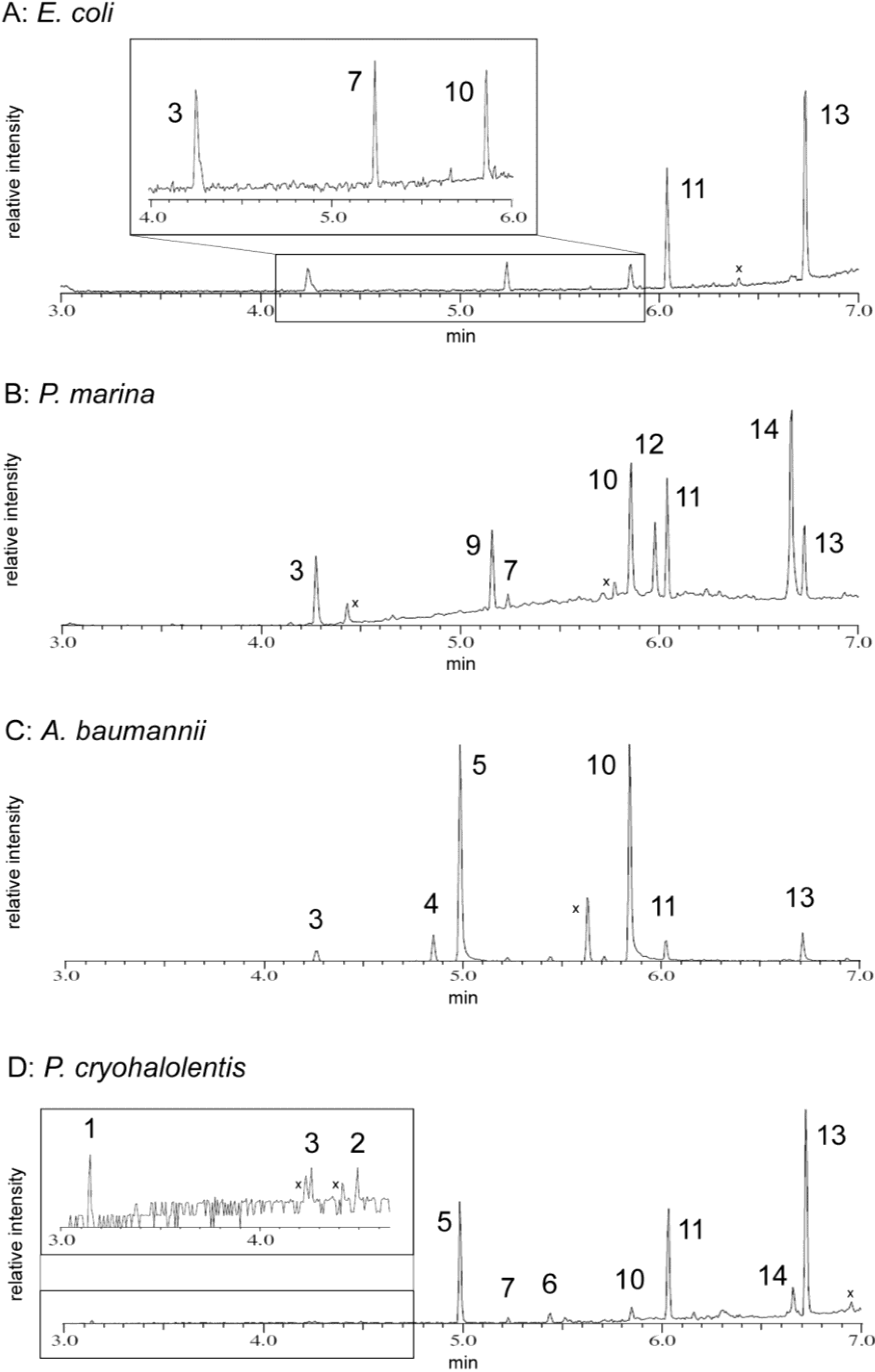
2.5. FAME GC-MS Analysis of Lipid Acyl Content
| Acyl Residue | Peak ID | Standard Retention Time | E. coli | P. marina | A. baumannii | P. cryohalolentis |
|---|---|---|---|---|---|---|
| From Lipid A: | ||||||
| decanoate (C10:0) | 1 | 3.160 | -- | -- | -- | 3.147 |
| 3-hydroxyundecanoate (3-OH C11:0) | 2 | 4.5 * | -- | -- | -- | 4.493 |
| dodecanoate (C12:0) | 3 | 4.264 | 4.265 | 4.273 | 4.264 | 4.260 |
| 2-hydroxydodecanoate (2-OH C12:0) | 4 | 4.853 | -- | -- | 4.855 | -- |
| 3-hydroxydodecanoate (3-OH C12:0) | 5 | 4.990 | -- | -- | 4.990 | 4.989 |
| 3-hydroxytridecanoate (3-OH C13:0) | 6 | 5.4 * | -- | -- | -- | 5.440 |
| tetradecanoate (C14:0) | 7 | 5.237 | 5.240 | 5.240 | -- | 5.230 |
| tetradecenoate (C14:1) | 8 | 5.193 | -- | -- | -- | -- |
| tetradecadienoate (C14:2) | 9 | not available | -- | 5.160 | -- | -- |
| 3-hydroxytetradecanoate (3-OH C14:0) | 10 | 5.856 | 5.860 | 5.853 | 5.843 | 5.847 |
| From Phospholipids: | ||||||
| hexadecanoate (C16:0) | 11 | 6.026 | 6.040 | 6.040 | 6.024 | 6.033 |
| hexadecenoate (C16:1) | 12 | 5.975 | -- | 5.980 | -- | -- |
| octadecanoate (C18:0) | 13 | 6.733 | 6.733 | 6.733 | 6.715 | 6.720 |
| octadecenoate (C18:1) | 14 | 6.667 | -- | 6.667 | -- | 6.653 |
2.5.1. E. coli Acyl Analysis
2.5.2. P. marina Acyl Analysis
2.5.3. A. baumannii Acyl Analysis
2.5.4. P. cryohalolentis Acyl Analysis

2.6. Discussion

3. Materials and Methods
3.1. Culture Conditions, Strains, and Reagents
3.2. Lipid A Isolation and DE52 Purification
3.3. MALDI-TOF MS
3.4. FAME (Fatty Acid Methyl Ester) GC-MS
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Beutler, B.; Rietschel, E.T. Innate immune sensing and its roots: The story of endotoxin. Nat. Rev. Immunol. 2003, 3, 169–176. [Google Scholar] [CrossRef]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar]
- Raetz, C.R.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A modification systems in gram-negative bacteria. Annu. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef]
- Montminy, S.W.; Khan, N.; McGrath, S.; Walkowicz, M.J.; Sharp, F.; Conlon, J.E.; Fukase, K.; Kusumoto, S.; Sweet, C.; Miyake, K.; et al. Virulence factors of Yersinia pestis are overcome by a strong lipopolysaccharide response. Nat. Immunol. 2006, 7, 1066–1073. [Google Scholar] [CrossRef]
- Zhou, Z.; Ribeiro, A.A.; Lin, S.; Cotter, R.J.; Miller, S.I.; Raetz, C.R. Lipid A modifications in polymyxin-resistant Salmonella typhimurium: PMRA-dependent 4-amino-4-deoxy-l-arabinose, and phosphoethanolamine incorporation. J. Biol. Chem. 2001, 276, 43111–43121. [Google Scholar]
- Murata, T.; Tseng, W.; Guina, T.; Miller, S.I.; Nikaido, H. PhoPQ-mediated regulation produces a more robust permeability barrier in the outer membrane of Salmonella enterica serovar typhimurium. J. Bacteriol. 2007, 189, 7213–7222. [Google Scholar] [CrossRef]
- Li, Y.; Powell, D.A.; Shaffer, S.A.; Rasko, D.A.; Pelletier, M.R.; Leszyk, J.D.; Scott, A.J.; Masoudi, A.; Goodlett, D.R.; Wang, X.; et al. LPS remodeling is an evolved survival strategy for bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 8716–8721. [Google Scholar] [CrossRef]
- Leone, S.; Silipo, A.; Nazarenko Evgeny, L.; Lanzetta, R.; Parrilli, M.; Molinaro, A. Molecular structure of endotoxins from Gram-negative marine bacteria: An update. Mar. Drugs 2007, 5, 85–112. [Google Scholar] [CrossRef]
- Gibbons, H.S.; Kalb, S.R.; Cotter, R.J.; Raetz, C.R. Role of Mg2+ and pH in the modification of Salmonella lipid A after endocytosis by macrophage tumour cells. Mol. Microbiol. 2005, 55, 425–440. [Google Scholar]
- Zhou, Z.; Ribeiro, A.A.; Raetz, C.R. High-resolution NMR spectroscopy of lipid A molecules containing 4-amino-4-deoxy-l-arabinose and phosphoethanolamine substituents. Different attachment sites on lipid A molecules from NH4VO3-treated Escherichia coli versus kdsA mutants of Salmonella typhimurium. J. Biol. Chem 2000, 275, 13542–13551. [Google Scholar] [CrossRef]
- Bishop, R.E.; Gibbons, H.S.; Guina, T.; Trent, M.S.; Miller, S.I.; Raetz, C.R. Transfer of palmitate from phospholipids to lipid A in outer membranes of gram-negative bacteria. EMBO J. 2000, 19, 5071–5080. [Google Scholar] [CrossRef]
- Phadtare, S. Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 2004, 6, 125–136. [Google Scholar]
- Graumann, P.L.; Marahiel, M.A. Cold shock response in Bacillus subtilis. J. Mol. Microbiol. Biotechnol. 1999, 1, 203–209. [Google Scholar]
- Miller, D.J.; Zhang, Y.M.; Subramanian, C.; Rock, C.O.; White, S.W. Structural basis for the transcriptional regulation of membrane lipid homeostasis. Nat. Struct. Mol. Biol. 2010, 17, 971–975. [Google Scholar] [CrossRef]
- Singh, A.K.; Zhang, Y.M.; Zhu, K.; Subramanian, C.; Li, Z.; Jayaswal, R.K.; Gatto, C.; Rock, C.O.; Wilkinson, B.J. FabH selectivity for anteiso branched-chain fatty acid precursors in low-temperature adaptation in Listeria monocytogenes. FEMS Microbiol. Lett. 2009, 301, 188–192. [Google Scholar] [CrossRef]
- Carty, S.M.; Sreekumar, K.R.; Raetz, C.R. Effect of cold shock on lipid A biosynthesis in Escherichia coli. Induction At 12 degrees C of an acyltransferase specific for palmitoleoyl-acyl carrier protein. J. Biol. Chem. 1999, 274, 9677–9685. [Google Scholar] [CrossRef]
- Kawasaki, K.; Nogi, Y.; Hishinuma, M.; Nodasaka, Y.; Matsuyama, H.; Yumoto, I. Psychromonas marina sp. nov., a novel halophilic, facultatively psychrophilic bacterium isolated from the coast of the Okhotsk Sea. Int. J. Syst. Evol. Microbiol. 2002, 52, 1455–1459. [Google Scholar] [CrossRef]
- Bakermans, C.; Ayala-del-Rio, H.L.; Ponder, M.A.; Vishnivetskaya, T.; Gilichinsky, D.; Thomashow, M.F.; Tiedje, J.M. Psychrobacter cryohalolentis sp. nov. and Psychrobacter arcticus sp. nov., isolated from Siberian permafrost. Int. J. Syst. Evol. Microbiol. 2006, 56, 1285–1291. [Google Scholar] [CrossRef]
- Williams, K.P.; Gillespie, J.J.; Sobral, B.W.; Nordberg, E.K.; Snyder, E.E.; Shallom, J.M.; Dickerman, A.W. Phylogeny of gammaproteobacteria. J. Bacteriol. 2010, 192, 2305–2314. [Google Scholar] [CrossRef]
- Sweet, C.R.; Williams, A.H.; Karbarz, M.J.; Werts, C.; Kalb, S.R.; Cotter, R.J.; Raetz, C.R. Enzymatic synthesis of lipid A molecules with four amide-linked acyl chains. LpxA acyltransferases selective for an analog of UDP-N-acetylglucosamine in which an amine replaces the 3″-hydroxyl group. J. Biol. Chem. 2004, 279, 25411–25419. [Google Scholar]
- Reynolds, C.M.; Ribeiro, A.A.; McGrath, S.C.; Cotter, R.J.; Raetz, C.R.; Trent, M.S. An outer membrane enzyme encoded by Salmonella typhimurium lpxR that removes the 3′-acyloxyacyl moiety of lipid A. J. Biol. Chem. 2006, 281, 21974–21987. [Google Scholar] [CrossRef]
- Trent, M.S.; Pabich, W.; Raetz, C.R.; Miller, S.I. A PhoP/PhoQ-induced Lipase (PagL) that catalyzes 3-O-deacylation of lipid A precursors in membranes of Salmonella typhimurium. J. Biol. Chem. 2001, 276, 9083–9092. [Google Scholar]
- Gibbons, H.S.; Reynolds, C.M.; Guan, Z.; Raetz, C.R. An inner membrane dioxygenase that generates the 2-hydroxymyristate moiety of Salmonella lipid A. Biochemistry 2008, 47, 2814–2825. [Google Scholar] [CrossRef]
- Fregolino, E.; Fugazza, G.; Galano, E.; Gargiulo, V.; Landini, P.; Lanzetta, R.; Lindner, B.; Pagani, L.; Parrilli, M.; Holst, O.; et al. Complete Lipooligosaccharide Structure of the Clinical Isolate Acinetobacter baumannii, Strain SMAL. Eur. J. Org. Chem. 2010, 2010, 1345–1352. [Google Scholar] [CrossRef]
- Yano, Y.; Nakayama, A.; Yoshida, K. Distribution of polyunsaturated Fatty acids in bacteria present in intestines of deep-sea fish and shallow-sea poikilothermic animals. Appl. Environ. Microbiol. 1997, 63, 2572–2577. [Google Scholar]
- Russell, N.J.; Nichols, D.S. Polyunsaturated fatty acids in marine bacteria—A dogma rewritten. Microbiology 1999, 145, 767–779. [Google Scholar] [CrossRef]
- Nichols, D.S.; Nichols, P.D.; Russell, N.J.; Davies, N.W.; McMeekin, T.A. Polyunsaturated fatty acids in the psychrophilic bacterium Shewanella gelidimarina ACAM 456T: Molecular species analysis of major phospholipids and biosynthesis of eicosapentaenoic acid. Biochim. Biophys. Acta 1997, 1347, 164–176. [Google Scholar] [CrossRef]
- Sato, S.; Kurihara, T.; Kawamoto, J.; Hosokawa, M.; Sato, S.B.; Esaki, N. Cold adaptation of eicosapentaenoic acid-less mutant of Shewanella livingstonensis Ac10 involving uptake and remodeling of synthetic phospholipids containing various polyunsaturated fatty acids. Extremophiles 2008, 12, 753–761. [Google Scholar] [CrossRef]
- Gao, J.; Ajjawi, I.; Manoli, A.; Sawin, A.; Xu, C.; Froehlich, J.E.; Last, R.L.; Benning, C. FATTY ACID DESATURASE4 of Arabidopsis encodes a protein distinct from characterized fatty acid desaturases. Plant J. 2009, 60, 832–839. [Google Scholar] [CrossRef]
- Attygalle, A.B.; Jham, G.N.; Svatos, A.; Frighetto, R.T.; Ferrara, F.A.; Vilela, E.F.; Uchoa-Fernandes, M.A.; Meinwald, J. (3E,8Z,11Z)-3,8,11-tetradecatrienyl acetate, major sex pheromone component of the tomato pest Scrobipalpuloides absoluta (Lepidoptera: Gelechiidae). Bioorg. Med. Chem. 1996, 4, 305–314. [Google Scholar] [CrossRef]
- Corsaro, M.M.; Piaz, F.D.; Lanzetta, R.; Parrilli, M. Lipid A structure of Pseudoalteromonas haloplanktis TAC 125: Use of electrospray ionization tandem mass spectrometry for the determination of fatty acid distribution. J. Mass Spectrom. 2002, 37, 481–488. [Google Scholar] [CrossRef]
- Sweet, C.R.; Preston, A.; Toland, E.; Ramirez, S.M.; Cotter, R.J.; Maskell, D.J.; Raetz, C.R. Relaxed acyl chain specificity of Bordetella UDP-N-acetylglucosamine acyltransferases. J. Biol. Chem. 2002, 277, 18281–18290. [Google Scholar]
- Kempf, M.; Rolain, J.M. Emergence of resistance to carbapenems in Acinetobacter baumannii in Europe: Clinical impact and therapeutic options. Int. J. Antimicrob. Agents 2012, 39, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Sebeny, P.J.; Riddle, M.S.; Petersen, K. Acinetobacter baumannii skin and soft-tissue infection associated with war trauma. Clin. Infect. Dis. 2008, 47, 444–449. [Google Scholar] [CrossRef]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; St Michael, F.; Cox, A.D.; et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef]
- Leone, S.; Sturiale, L.; Pessione, E.; Mazzoli, R.; Giunta, C.; Lanzetta, R.; Garozzo, D.; Molinaro, A.; Parrilli, M. Detailed characterization of the lipid A fraction from the nonpathogen Acinetobacter radioresistens strain S13. J. Lipid Res. 2007, 48, 1045–1051. [Google Scholar] [CrossRef]
- Corsaro, M.M.; Lanzetta, R.; Parrilli, E.; Parrilli, M.; Tutino, M.L.; Ummarino, S. Influence of growth temperature on lipid and phosphate contents of surface polysaccharides from the antarctic bacterium Pseudoalteromonas haloplanktis TAC 125. J. Bacteriol. 2004, 186, 29–34. [Google Scholar] [CrossRef]
- Shaffer, S.A.; Harvey, M.D.; Goodlett, D.R.; Ernst, R.K. Structural heterogeneity and environmentally regulated remodeling of Francisella tularensis subspecies novicida lipid A characterized by tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2007, 18, 1080–1092. [Google Scholar] [CrossRef]
- American Type Culture Collection (ATCC). Available online: http://www.atcc.org (accessed on 4 May 2014).
- National Center for Biotechnology Information (NCBI USA). Available online: http://www.ncbi.nlm.nih.gov (accessed on 4 May 2014).
- Sweet, C.R.; Lin, S.; Cotter, R.J.; Raetz, C.R. A Chlamydia trachomatis UDP-N-acetylglucosamine acyltransferase selective for myristoyl-acyl carrier protein. Expression in Escherichia coli and formation of hybrid lipid A species. J. Biol. Chem. 2001, 276, 19565–19574. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sweet, C.R.; Alpuche, G.M.; Landis, C.A.; Sandman, B.C. Endotoxin Structures in the Psychrophiles Psychromonas marina and Psychrobacter cryohalolentis Contain Distinctive Acyl Features. Mar. Drugs 2014, 12, 4126-4147. https://doi.org/10.3390/md12074126
Sweet CR, Alpuche GM, Landis CA, Sandman BC. Endotoxin Structures in the Psychrophiles Psychromonas marina and Psychrobacter cryohalolentis Contain Distinctive Acyl Features. Marine Drugs. 2014; 12(7):4126-4147. https://doi.org/10.3390/md12074126
Chicago/Turabian StyleSweet, Charles R., Giancarlo M. Alpuche, Corinne A. Landis, and Benjamin C. Sandman. 2014. "Endotoxin Structures in the Psychrophiles Psychromonas marina and Psychrobacter cryohalolentis Contain Distinctive Acyl Features" Marine Drugs 12, no. 7: 4126-4147. https://doi.org/10.3390/md12074126
APA StyleSweet, C. R., Alpuche, G. M., Landis, C. A., & Sandman, B. C. (2014). Endotoxin Structures in the Psychrophiles Psychromonas marina and Psychrobacter cryohalolentis Contain Distinctive Acyl Features. Marine Drugs, 12(7), 4126-4147. https://doi.org/10.3390/md12074126