Beta-N-Methylamino-l-Alanine: LC-MS/MS Optimization, Screening of Cyanobacterial Strains and Occurrence in Shellfish from Thau, a French Mediterranean Lagoon


 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of the Analysis of BMAA and Isomers
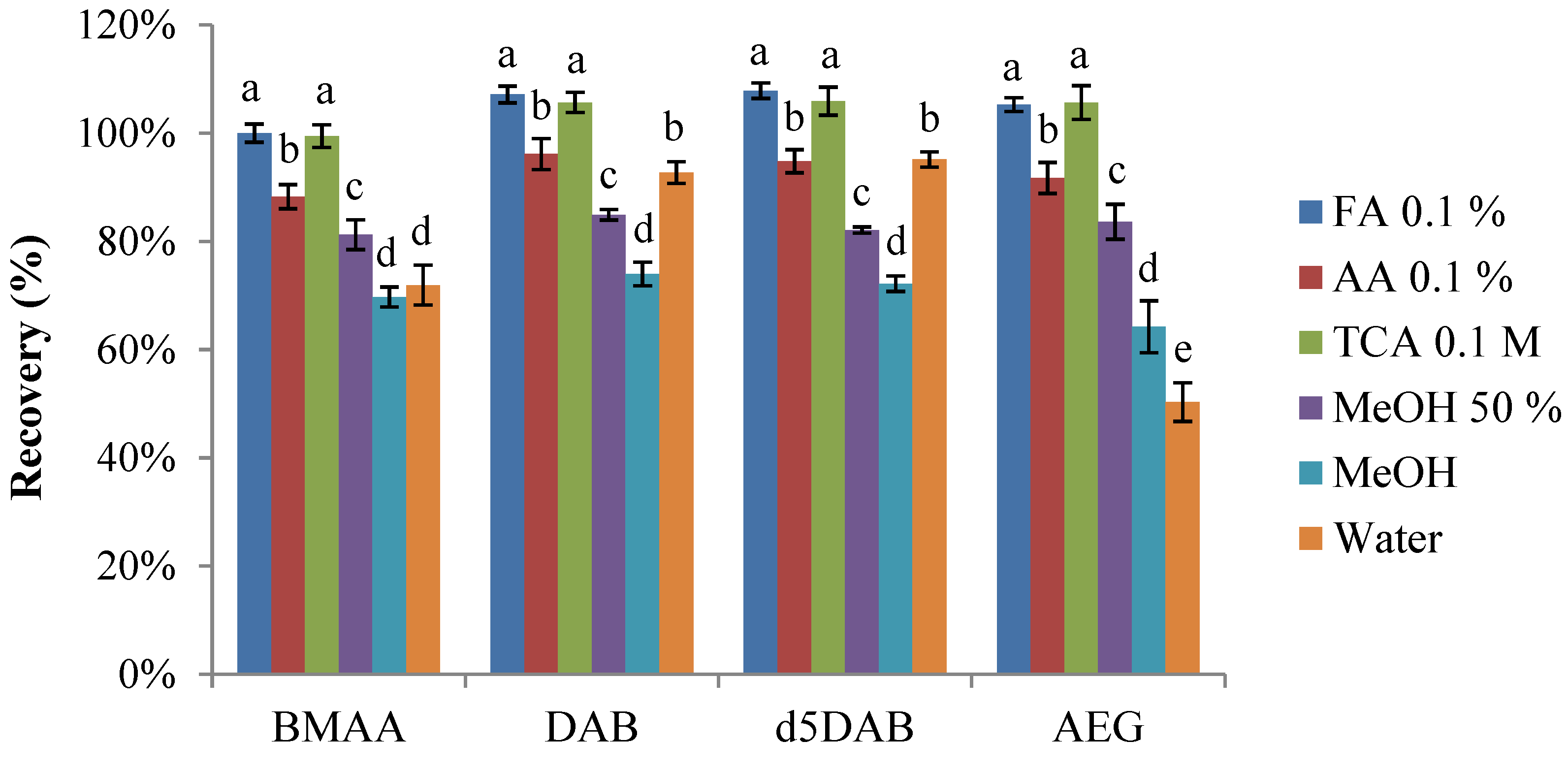
2.1.1. Cell Lysis and Solvent Extraction


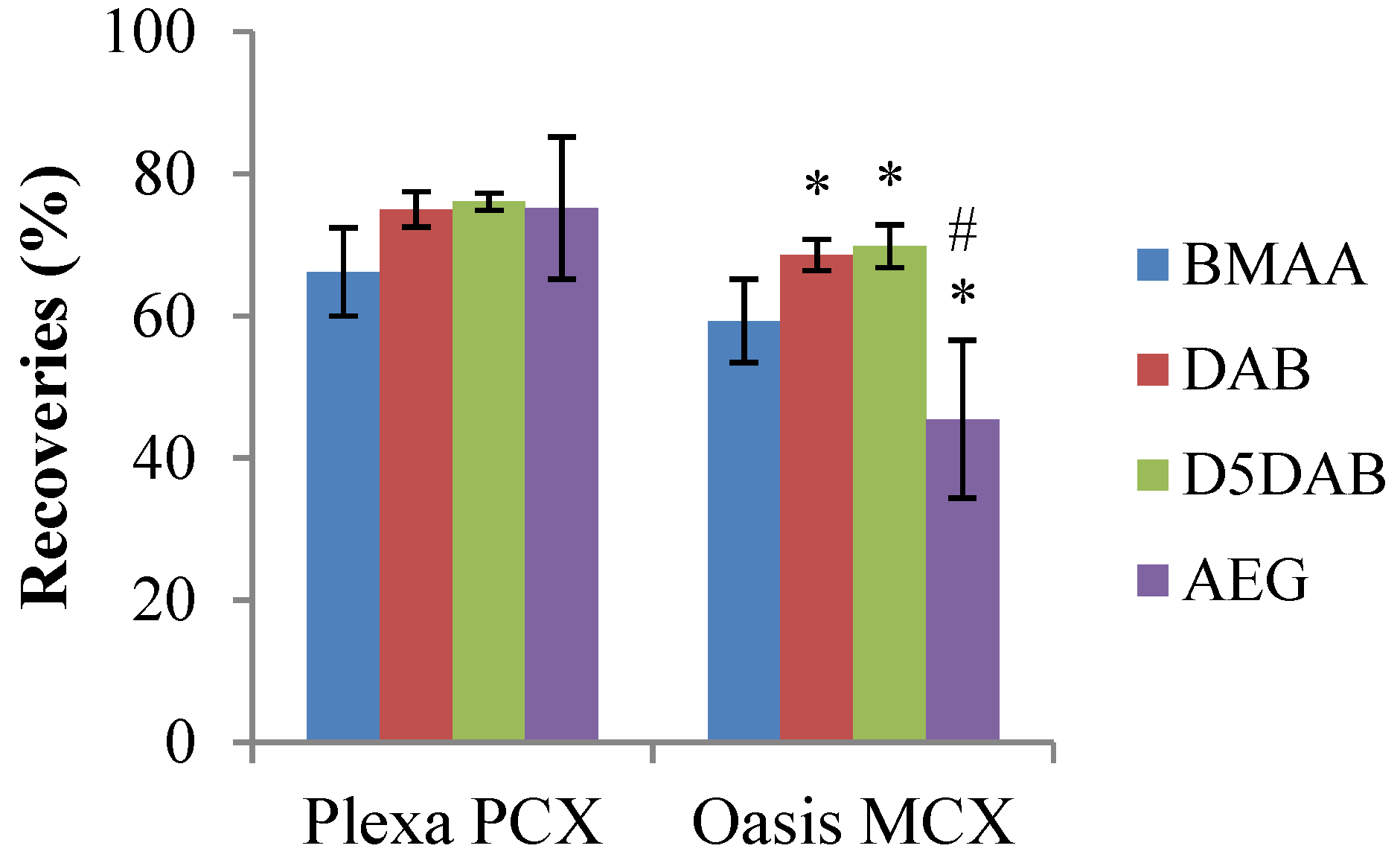
2.1.2. Comparison of Solid Phase Extraction (SPE) Sorbents

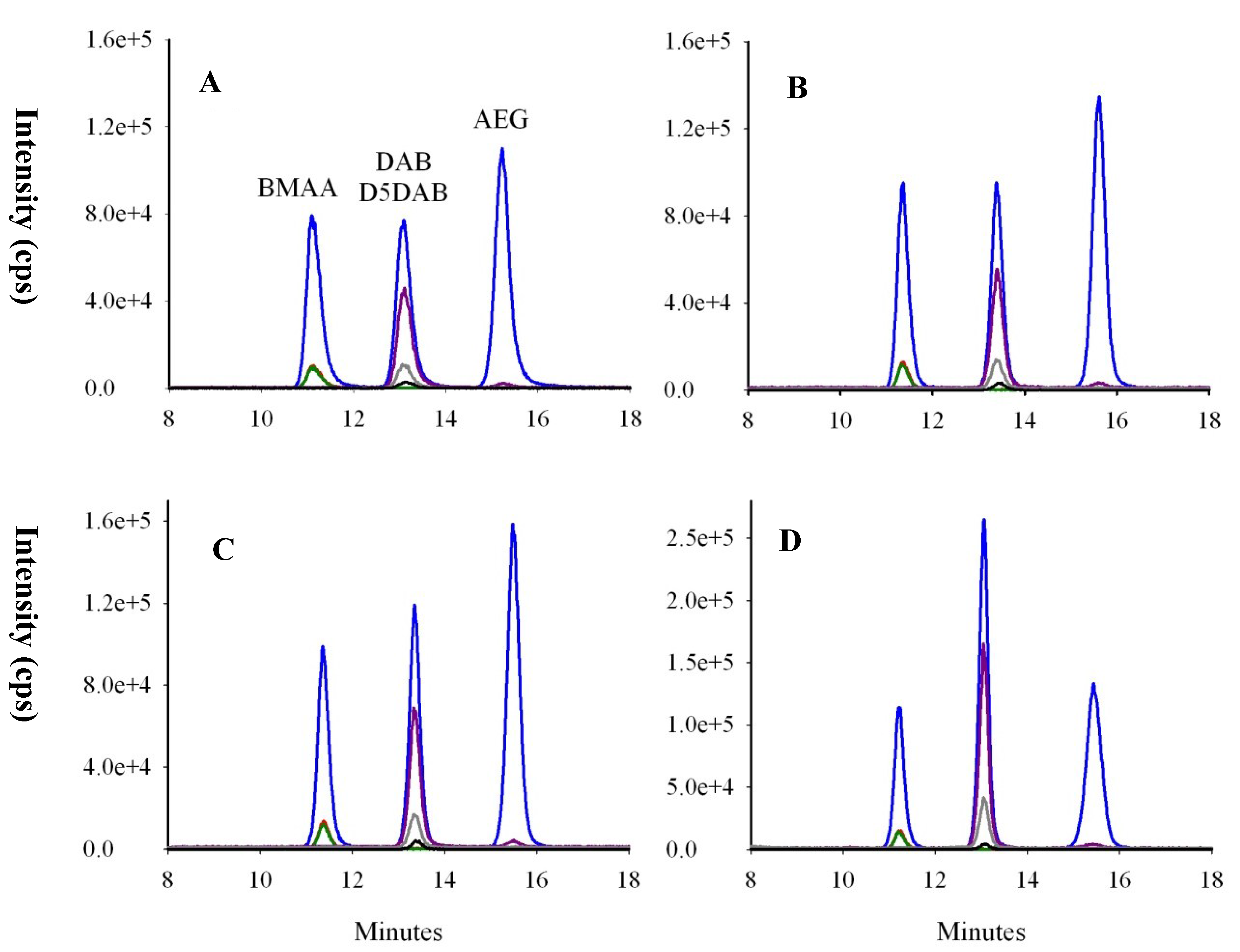
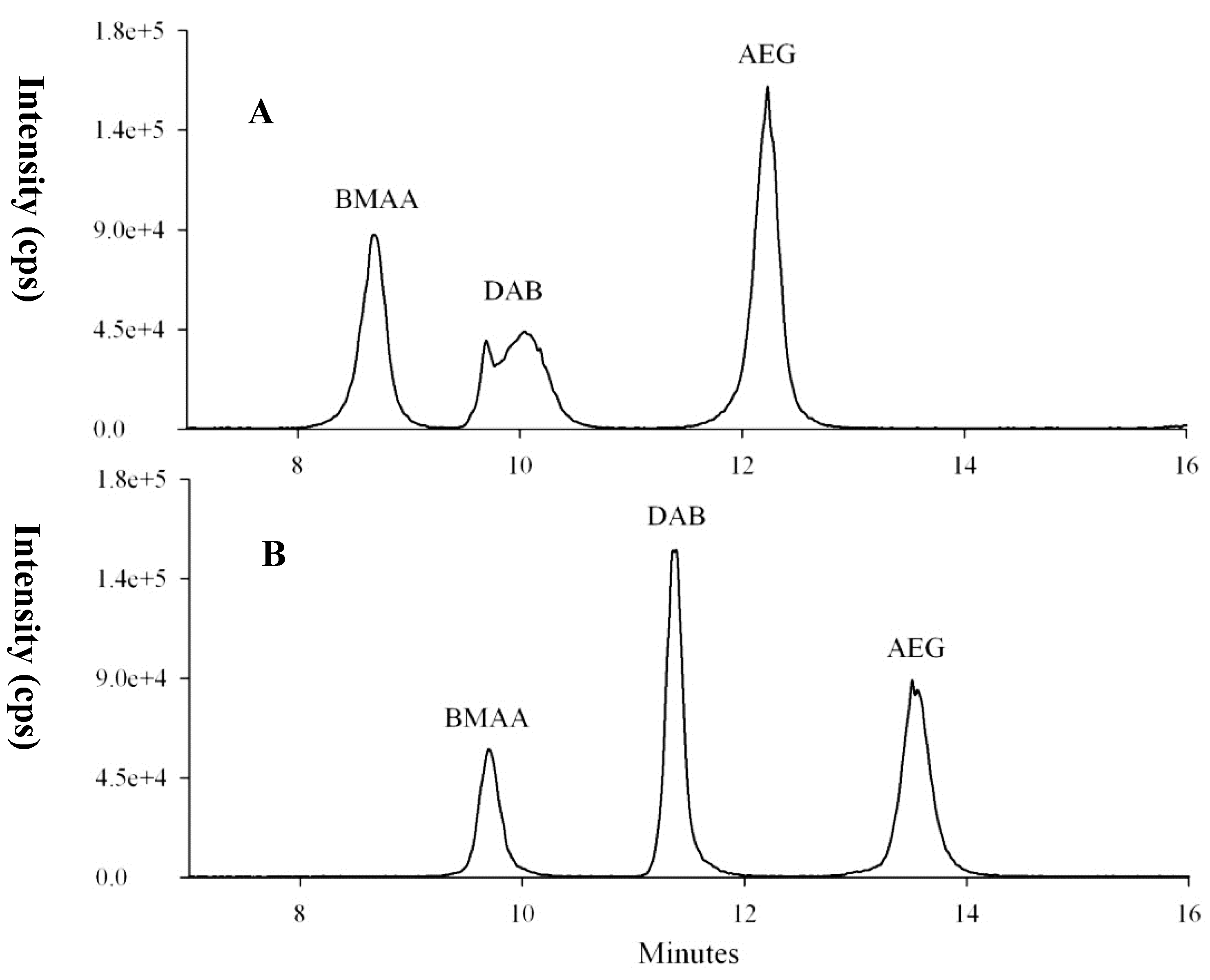
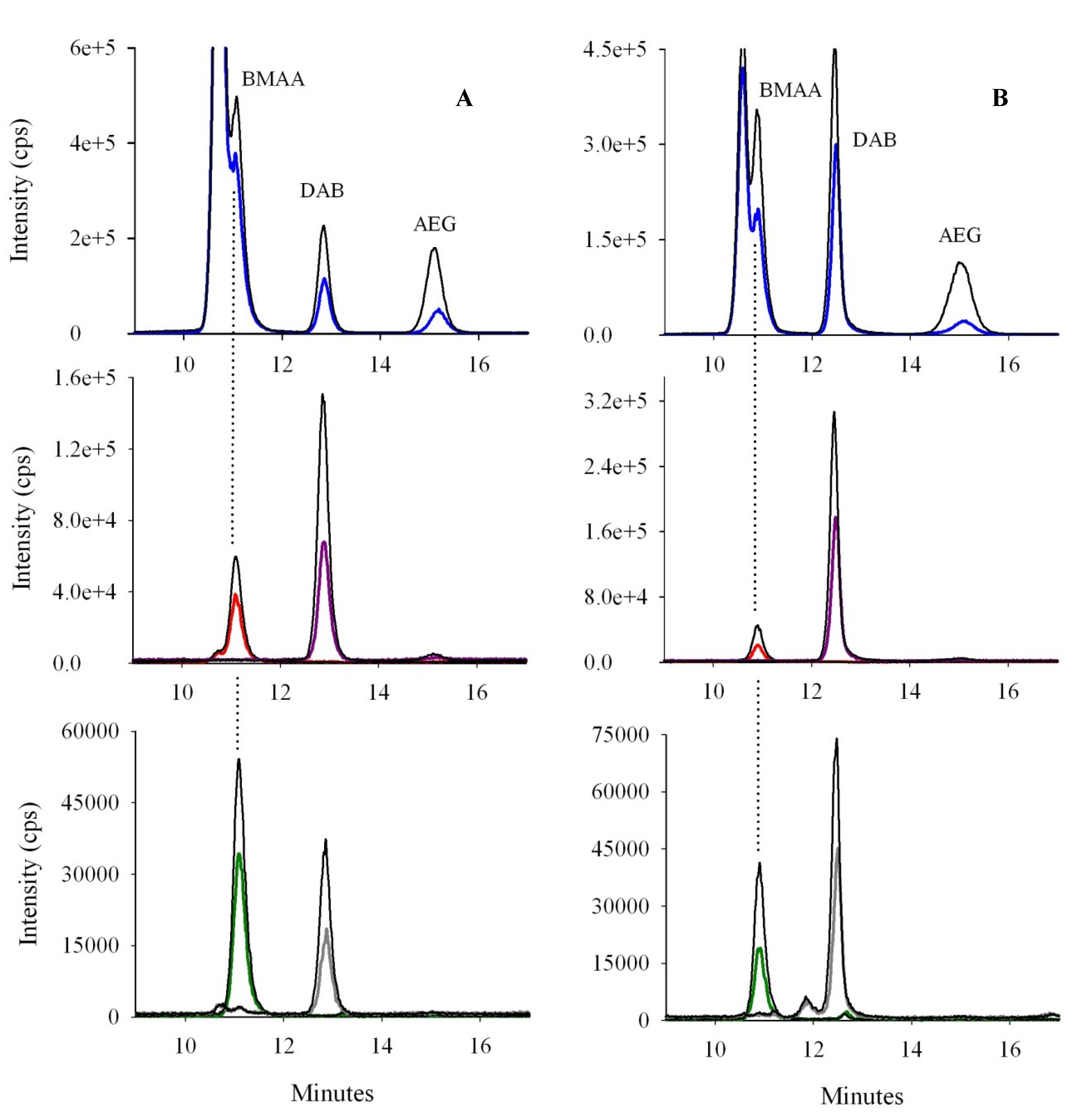
2.1.3. Optimization of Selectivity: Chromatographic Resolution and Mass Spectral Transitions


2.2. Characterization of the Analytical Procedure
2.2.1. Recoveries of SPE Clean-Up and Remaining Matrix Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction | Matrix | Recoveries of SPE Clean-Up (mean% ± SD) | Matrix Effect (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BMAA | DAB | D5DAB | AEG | BMAA | DAB | D5DAB | AEG | ||
| Free analytes | Cyanobacteria | 61.1 ± 4.9 | 51.0 ± 5.6 | 61.0 ± 1.6 | 48.1 ± 5.5 | 8.8 | 5.6 | 4.7 | 9.3 |
| Oyster | 56.1 ± 4.5 | 64.5 ± 4.5 | 65.4 ± 4.5 | 63.1 ± 3.8 | 7.3 | 0.9 | −3.5 | 0.2 | |
| Mussel | 63.8 ± 3.1 | 79.0 ± 6.8 | 75.6 ± 5.7 | 73.3 ± 2.1 | 5.3 | −6.3 | 1.2 | 0.7 | |
| Total analytes | Cyanobacteria | 67.6 ± 3.3 | 63.1 ± 1.9 | 63.4 ± 7.5 | 59.5 ± 1.0 | 3.8 | 1.9 | 4.2 | −9 |
| Oyster | 71.0 ± 2.0 | 76.4 ± 9.5 | 81.1 ± 4.1 | 74.0 ± 6.4 | 7.5 | 3.5 | 3 | 3.6 | |
| Mussel | 64.3 ± 8.0 | 65.0 ± 8.1 | 73.1 ± 6.5 | 64.3 ± 0.9 | 8.7 | 12.1 | 15.7 | −5.5 | |

2.2.2. LC-MS/MS Performance: Linearity, LOD, LOQ and Repeatabilites (RT, R2 and Ion Ratios)
| Samples | RT (min) | R2 | Ion Ratios (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BMAA | DAB | D5DAB | AEG | BMAA | DAB | D5DAB | AEG | 88/102 | 76/102 | 101/102 | 74/102 | |
| Standards | 11.25 ± 0.08 | 13.27 ± 0.10 | 13.34 ± 0.11 | 15.47 ± 0.11 | 0.9999 | 0.9998 | 0.9996 | 0.9996 | 13.1 ± 0.8 | 12 ± 0.8 | 60.3 ± 2 | 14.2 ± 0.8 |
| 73110 a | 11.42 ± 0.05 | 13.39 ± 0.08 | 13.47 ± 0.08 | 15.50 ± 0.13 | 0.9997 | 0.9995 | 0.9995 | 0.9995 | 13.0 ± 0.1 | 12.0 ± 0.4 | 58.8 ± 2 | 14.7 ± 1 |
| 73110 b | 11.12 ± 0.04 | 13.03 ± 0.04 | 13.08 ± 0.04 | 15.17 ± 0.09 | 0.9998 | 0.9973 | 0.9992 | 0.9990 | 12.6 ± 1 | 11.5 ± 1 | 58.9 ± 4 | 14.3 ± 1 |
| Oyster a | 11.42 ± 00.5 | 13.41 ± 0.06 | 13.44 ± 0.07 | 15.55 ± 0.06 | 0.9999 | 0.9991 | 0.9997 | 0.9993 | 12.4 ± 1 | 11.3 ± 1 | 58.6 ± 1 | 14 ± 0.4 |
| Oyster b | 11.06 ± 0.04 | 12.82 ± 0.09 | 12.88 ± 0.10 | 15.08 ± 0.14 | 0.9986 | 0.9807 | 0.9993 | 0.9915 | 12.0 ± 0.5 | 10.5 ± 0.4 | 58.4 ± 2 | 14.4 ± 0.6 |
| Mussel a | 11.30 ± 0.07 | 13.14 ± 0.07 | 13.17 ± 0.07 | 15.46 ± 0.11 | 0.9997 | 0.9983 | 0.9996 | 0.9995 | 12.8 ± 0.9 | 11.5 ± 0.9 | 59.4 ± 0.6 | 14.6 ± 0.3 |
| Mussel b | 10.91 ± 0.03 | 12.53 ± 0.05 | 12.53 ± 0.05 | 15.14 ± 0.11 | 0.9975 | 0.9498 | 0.9990 | 0.9919 | 12.0 ± 0.3 | 10.5 ± 0.4 | 58.5 ± 0.7 | 14.4 ± 0.4 |
2.3. Screening of BMAA and Isomers in Cyanobacteria
2.3.1. Kinetics of Growth and Production of BMAA and Isomers

2.3.2. Screening of Others Lab-Cultured Cyanobacteria
| Cyanobacterial Strain | Free DAB (µg/g DW) | Total DAB (µg/g DW) |
|---|---|---|
| Leptolyngbya PCC 73110 a | <LOD–2.69 | 1.01–1.57 |
| Nostoc CCMP 2511/ CMMED01 a | <LOD–2.27 | 0.82–12.48 |
| Microcystis PCC 7806 a | <LOD | <LOD |
| Nostoc PCC 7120 a | <LOD | * |
| Nostoc PCC 7107 a | 2.12–7.2 | * |
| Symploca PCC 8002 a | 0.3 | 0.43 |
| Synechocystis sp.PCC 6803 | <LOD | <LOD |
| Synechococcus elongatus CCAP1479/1B | 0.23–2.71 | 0.4–3.52 |
| Calothrix crustacea CCAP1410/9 | 0.6–0.92 | 6.95–14.53 |
| Nostoc endophytum CCAP1453/14 | 0.34–0.55 | 6.78–7.52 |
2.3.3. Screening of Mollusks of Thau Lagoon


3. Experimental Section
3.1. Chemicals and Reagents
3.2. Samples
3.2.1. Cultures of Cyanobacteria
3.2.2. Collection of Field Samples
3.3. Sample Preparation
3.3.1. Extraction
3.3.2. SPE Clean-Up
3.4. Analysis by LC-MS/MS
 with Rt the retention time and w the width at the base of the common mass spectral transition m/z 119 > 102. Results were analyzed and interpreted using STATGRAPHICS Centurion XV software.
with Rt the retention time and w the width at the base of the common mass spectral transition m/z 119 > 102. Results were analyzed and interpreted using STATGRAPHICS Centurion XV software.| Transition (m/z) | EP (V) | DP (V) | CE (eV) | CXP (V) |
|---|---|---|---|---|
| 119 > 102 | 10 | 81 | 13 | 12 |
| 124 > 47 | 71 | 21 | 22 | |
| 119 > 88 | 66 | 17 | 10 | |
| 119 > 76 | 66 | 17 | 10 | |
| 119 > 101 | 86 | 11 | 8 | |
| 119 > 74 | 86 | 19 | 8 |
3.5. Recoveries and Matrix Effects
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vega, A.; Bell, E.A. α-Amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Chiu, A.S.; Gehringer, M.M.; Welch, J.H.; Neilan, B.A. Does α-amino-β-methylaminopropionic acid (BMAA) play a role in neurodegeneration? Int. J. Environ. Res. Public Health 2011, 8, 3728–3746. [Google Scholar] [CrossRef]
- Karamyan, V.T.; Speth, R.C. Animal models of BMAA neurotoxicity: A critical review. Life Sci. 2008, 82, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neurodegenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Lindsay, J.; Morrison, L.F.; Cox, P.A.; Codd, G.A. Co-occurrence of β-N-methylamino-l-alanine, a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen, M.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Safety 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Tian, Z.; Li, J.; Yu, R.; Banack, S.A.; Wang, Z. Detection of the neurotoxin BMAA within cyanobacteria isolated from freshwater in China. Toxicon 2010, 55, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Spáčil, Z.; Erkisson, J.; Jonasson, S.; Rasmussen, U.; Ilag, L.L.; Bergman, B. Analytical protocol for identification of BMAA and DAB in biological samples. Analyst 2010, 135, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spáčil, Z.; Ilag, L.L.; Ronnevi, L.-O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Pablo, J.; Compton, A.; Hammerschlag, N.; Mash, D.C. Cyanobacterial blooms and the occurrence of the neurotoxin, beta-N-methylamino-l-alanine (BMAA), in South Florida aquatic food webs. Harmful Algae 2010, 9, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Chen, Q.; Chen, X.; Wang, X.; Liao, X.; Jiang, L.; Wu, J.; Yang, L. Occurrence and transfer of a cyanobacterial neurotoxin beta-methylamino-l-alanine within the aquatic food webs of Gonghu Bay (Lake Taihu, China) to evaluate the potential human health risk. Sci. Total Environ. 2014, 468, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Pablo, J.; Banack, S.A.; Cox, P.A.; Johnson, T.E.; Papapetropoulos, S.; Bradley, W.G.; Buck, A.; Mash, D.C. Cyanobacterial neurotoxin BMAA in ALS and Alzheimer’s disease. Acta Neurol. Scand. 2009, 120, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Kruger, T.; Oelmuller, R.; luckas, B. The origin of β-N-methylamino-l-alanine (BMAA): Cycads and/-or cyanobacteria? J. Endocytobiosis Cell Res. 2012, 22, 29–36. [Google Scholar]
- Faassen, E.J. Presence of the neurotoxin BMAA in aquatic ecosystems: What do we really know? Toxins 2014, 1109–1138. [Google Scholar] [CrossRef]
- Faassen, E.J.; Gillissen, F.; Lürling, M. A Comparative Study on Three Analytical Methods for the Determination of the Neurotoxin BMAA in Cyanobacteria. PLoS One 2012, 7, e36667. [Google Scholar] [CrossRef]
- Krüger, T.; Mönch, B.; Oppenhäuser, S.; Luckas, B. LC–MS/MS determination of the isomeric neurotoxins BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in cyanobacteria and seeds of Cycas revoluta and Lathyrus latifolius. Toxicon 2010, 55, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Kubo, T.; Kato, N.; Hosoya, K.; Kaya, K. Effective determination method for a cyanobacterial neurotoxin, β-N-methylamino-l-alanine. Toxicon 2008, 51, 1264–1268. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.; Hellenas, K.E. Determination of the neurotoxin BMAA (beta-N-methylamino-l-alanine) in cycad seed and cyanobacteria by LC-MS/MS (liquid chromatography tandem mass spectrometry). Analyst 2008, 133, 1785–1789. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Downing, T.G.; Spácil, Z.; Purdie, E.L.; Metcalf, J.S.; Downing, S.; Esterhuizen, M.; Codd, G.A.; Cox, P.A. Distinguishing the cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) from its structural isomer 2,4-diaminobutyric acid (2,4-DAB). Toxicon 2010, 56, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Combes, A.; El Abdellaoui, S.; Sarazin, C.; Vial, J.; Mejean, A.; Ploux, O.; Pichon, V.; Group, B. Validation of the analytical procedure for the determination of the neurotoxin β-N-methylamino-l-alanine in complex environmental samples. Anal. Chim. Acta 2013, 771, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Johnston, E.; Åberg, K.M.; Nilsson, U.; Ilag, L. Strategy for quantifying trace levels of BMAA in cyanobacteria by LC/MS/MS. Anal. Bioanal. Chem. 2013, 405, 1283–1292. [Google Scholar]
- Banack, S.A.; Metcalf, J.S.; Spacil, Z.; Downing, T.G.; Downing, S.; Long, A.; Nunn, P.B.; Cox, P.A. Distinguishing the cyanobacterial neurotoxin beta-N-methylamino-l-alanine (BMAA) from other diamino acids. Toxicon 2011, 57, 730–738. [Google Scholar] [CrossRef]
- Jiang, L.; Aigret, B.; Borggraeve, W.; Spacil, Z.; Ilag, L. Selective LC-MS/MS method for the identification of BMAA from its isomers in biological samples. Anal. Bioanal. Chem. 2012, 403, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Metcalf, J.S.; Jiang, L.; Craighead, D.; Ilag, L.L.; Cox, P.A. Cyanobacteria produce N-(2-aminoethyl) glycine, a backbone for peptide nucleic acids which may have been the first genetic molecules for life on Earth. PLoS One 2012, 7, e49043–e49043. [Google Scholar] [CrossRef]
- Berntzon, L.; Erasmie, S.; Celepli, N.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc sp.PCC 7120. Mar. Drugs 2013, 11, 3091–3108. [Google Scholar]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS One 2014, 9, e84578. [Google Scholar]
- Jiang, L.; Ilag, L.L. Detection of endogenous BMAA in dinoflagellate (Heterocapsa triquetra) hints at evolutionary conservation and environmental concern. PubRaw Sci. 2014, 1, 1–8. [Google Scholar]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The Non-Protein Amino Acid BMAA Is Misincorporated into Human Proteins in Place of l-Serine Causing Protein Misfolding and Aggregation. PLoS One 2013, 8, e75376. [Google Scholar]
- Cohen, S.A. Analytical techniques for the detection of alpha-amino-beta-methylaminopropionic acid. Analyst 2012, 137, 1991–2005. [Google Scholar] [CrossRef] [PubMed]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. The French Network on A.L.S.: Investigation, dietary bmaa exposure in an amyotrophic lateral sclerosis cluster from Southern France. PLoS One 2013, 8, e83406. [Google Scholar]
- Banack, S.; Johnson, H.; Cheng, R.; Cox, P. Production of the Neurotoxin BMAA by a Marine Cyanobacterium. Mar. Drugs 2007, 5, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.; Jonasson, S.; Papaefthimiou, D.; Rasmussen, U.; Bergman, B. Improving derivatization efficiency of BMAA utilizing AccQ-Tag® in a complex cyanobacterial matrix. Amino Acids 2009, 36, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, E.; Fedewa, E.; Richardson, T. Extraction protocols for the quantification of phycobilins in aqueous phytoplankton extracts. J. Appl. Phycol. 2011, 23, 865–871. [Google Scholar] [CrossRef]
- Serive, B.; Kaas, R.; Berard, J.-B.; Pasquet, V.; Picot, L.; Cadoret, J.-P. Selection and optimisation of a method for efficient metabolites extraction from microalgae. Bioresource Technol. 2012, 124, 311–320. [Google Scholar] [CrossRef]
- Ravn, H.; Anthoni, U.; Christophersen, C.; Nielsen, P.H.; Oshima, Y. Standardized extraction method for paralytic shellfish toxins in phytoplankton. J. Appl. Phycol. 1995, 7, 589–594. [Google Scholar] [CrossRef]
- Zendong, Z.; Herrenknecht, C.; Abadie, E.; Brissard, C.; Tixier, C.; Mondeguer, F.; Séchet, V.; Amzil, Z.; Hess, P. Extended evaluation of polymeric and lipophilic sorbents for passive sampling of marine toxins. Toxicon 2014. [Google Scholar] [CrossRef]
- Chambers, E.; Wagrowski-Diehl, D.M.; Lu, Z.; Mazzeo, J.R. Systematic and comprehensive strategy for reducing matrix effects in LC/MS/MS analyses. J. Chromatogr. B 2007, 852, 22–34. [Google Scholar] [CrossRef]
- Gerssen, A.; McElhinney, M.A.; Mulder, P.P.J.; Bire, R.; Hess, P.; de Boer, J. Solid phase extraction for removal of matrix effects in lipophilic marine toxin analysis by liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2009, 394, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; Fux, E. Strategies for the elimination of matrix effects in the liquid chromatography tandem mass spectrometry analysis of the lipophilic toxins okadaic acid and azaspiracid-1 in molluscan shellfish. J. Chromatogr. A 2010, 1217, 7123–7130. [Google Scholar] [CrossRef] [PubMed]
- Li, A.F.; Fan, H.; Ma, F.F.; McCarron, P.; Thomas, K.; Tang, X.H.; Quilliam, M.A. Elucidation of matrix effects and performance of solid-phase extraction for LC-MS/MS analysis of b-N-methylamino-l-alanine (BMAA) and 2,4-diaminobutyric acid (DAB) neurotoxins in cyanobacteria. Analyst 2012, 137, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.M.; Niedzwadek, B.; Rawn, D.F.K.; Lau, B.P.Y. Liquid chromatographic determination of the cyanobacterial toxin b-N-methylamino-l-alanine in algae food supplements, freshwater fish, and bottled water. J. Food Prot. 2009, 78, 1769–1773. [Google Scholar]
- Yoshida, T. Peptide separation by Hydrophilic-Interaction Chromatography: A review. J. Biochem. Biophys. Methods 2004, 60, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Agilent Technologies. Bond Elut Plexa PCX. Available online: (accessed on 13 November 2014).
- Kasprzyk-Hordern, B.; Dinsdale, R.M.; Guwy, A.J. The effect of signal suppression and mobile phase composition on the simultaneous analysis of multiple classes of acidic/neutral pharmaceuticals and personal care products in surface water by solid-phase extraction and ultra performance liquid chromatography-negative electrospray tandem mass spectrometry. Talanta 2008, 74, 1299–1312. [Google Scholar] [CrossRef]
- Jauffrais, T.; Herrenknecht, C.; Séchet, V.; Sibat, M.; Tillmann, U.; Krock, B.; Kilcoyne, J.; Miles, C.; McCarron, P.; Amzil, Z.; et al. Quantitative analysis of azaspiracids in Azadinium spinosum cultures. Anal. Bioanal. Chem. 2012, 403, 833–846. [Google Scholar]
- Glover, W.B.; Liberto, C.M.; McNeil, W.S.; Banack, S.A.; Shipley, P.R.; Murch, S.J. Reactivity of β-methylamino-l-alanine in Complex Sample Matrixes Complicating Detection and Quantification by Mass Spectrometry. Anal. Chem. 2012, 84, 7946–7953. [Google Scholar] [CrossRef] [PubMed]
- Downing, S.; Contardo-Jara, V.; Pflugmacher, S.; Downing, T.G. The fate of the cyanobacterial toxin beta-N-methylamino-l-alanine in freshwater mussels. Ecotoxicol. Environ. Safety 2014, 101, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.J.; Hemscheidt, T.K.; Trapido-Rosenthal, H.; Laws, E.A.; Bidigare, R.R. Detection and quantification of beta-methylamino-l-alanine in aquatic invertebrates. Limnol. Oceanogr. Methods 2012, 10, 891–898. [Google Scholar] [CrossRef]
- Salomonsson, M.L.; Hansson, A.; Bondesson, U. Development and in-house validation of a method for quantification of BMAA in mussels using dansyl chloride derivatization and ultra performance liquid chromatography tandem mass spectrometry. Anal. Methods UK 2013, 5, 4865–4874. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-l-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.L.; Downing, S.; Phelan, R.R.; Downing, T.G. Environmental modulation of microcystin and beta-N-methylamino-l-alanine as a function of nitrogen availability. Toxicon 2014, 87, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Caller, T.A.; Stommel, E.W. The cyanobacteria derived toxin beta-N-methylamino-l-alanine and amyotrophic lateral sclerosis. Toxins 2010, 2, 2837–2850. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; McGeer, P.L. , Weak BMAA toxicity compares with that of the dietary supplement beta-alanine. Neurobiol. Aging 2012, 33, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Field, N.C.; Metcalf, J.S.; Caller, T.A.; Banack, S.A.; Cox, P.A.; Stommel, E.W. Linking beta-methylamino-l-alanine exposure to sporadic amyotrophic lateral sclerosis in Annapolis, MD. Toxicon 2013, 70, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Pernet, F.; Malet, N.; Pastoureaud, A.; Vaquer, A.; Quéré, C.; Dubroca, L. Marine diatoms sustain growth of bivalves in a Mediterranean lagoon. J. Sea Res. 2012, 68, 20–32. [Google Scholar] [CrossRef]
- Mondo, K.; Broc Glover, W.; Murch, S.J.; Liu, G.; Cai, Y.; Davis, D.A.; Mash, D.C. Environmental neurotoxins beta-N-methylamino-l-alanine (BMAA) and mercury in shark cartilage dietary supplements. Food Chem. Toxicol. 2014, 70, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.H.; Christine, C.W.; Choi, D.W. Bicarbonate Dependence of Glutamate Receptor Activation by Beta-N-methylamino-l-alanine—Channel recording and study with related-compounds. Neuron 1989, 3, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Okle, O.; Stemmer, K.; Deschl, U.; Dietrich, D.R. (l)-BMAA induced er stress and enhanced caspase 12 cleavage in human neuroblastoma sh-sy5y cells at low nonexcitotoxic concentrations. Toxicol. Sci. 2013, 131, 217–224. [Google Scholar] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [PubMed]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis-immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar]
- Tompkins, J.; DeVille, M.M.; Day, J.G.; Turner, M.F. Culture collection of algae and protozoa. In Catalogue of Strains; Titus Wilson & Son lmtd: Ambleside, UK, 1995; p. 204. [Google Scholar]
- Guillard, R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W., Chanley, M., Eds.; Springer: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Réveillon, D.; Abadie, E.; Séchet, V.; Brient, L.; Savar, V.; Bardouil, M.; Hess, P.; Amzil, Z. Beta-N-Methylamino-l-Alanine: LC-MS/MS Optimization, Screening of Cyanobacterial Strains and Occurrence in Shellfish from Thau, a French Mediterranean Lagoon. Mar. Drugs 2014, 12, 5441-5467. https://doi.org/10.3390/md12115441
Réveillon D, Abadie E, Séchet V, Brient L, Savar V, Bardouil M, Hess P, Amzil Z. Beta-N-Methylamino-l-Alanine: LC-MS/MS Optimization, Screening of Cyanobacterial Strains and Occurrence in Shellfish from Thau, a French Mediterranean Lagoon. Marine Drugs. 2014; 12(11):5441-5467. https://doi.org/10.3390/md12115441
Chicago/Turabian StyleRéveillon, Damien, Eric Abadie, Véronique Séchet, Luc Brient, Véronique Savar, Michèle Bardouil, Philipp Hess, and Zouher Amzil. 2014. "Beta-N-Methylamino-l-Alanine: LC-MS/MS Optimization, Screening of Cyanobacterial Strains and Occurrence in Shellfish from Thau, a French Mediterranean Lagoon" Marine Drugs 12, no. 11: 5441-5467. https://doi.org/10.3390/md12115441
APA StyleRéveillon, D., Abadie, E., Séchet, V., Brient, L., Savar, V., Bardouil, M., Hess, P., & Amzil, Z. (2014). Beta-N-Methylamino-l-Alanine: LC-MS/MS Optimization, Screening of Cyanobacterial Strains and Occurrence in Shellfish from Thau, a French Mediterranean Lagoon. Marine Drugs, 12(11), 5441-5467. https://doi.org/10.3390/md12115441