Polyketide Synthases in the Microbiome of the Marine Sponge Plakortis halichondrioides: A Metagenomic Update



Abstract
:
1. Introduction
2. Results
2.1. Diversity of Polyketide Synthase Genes from P. halichondrioides
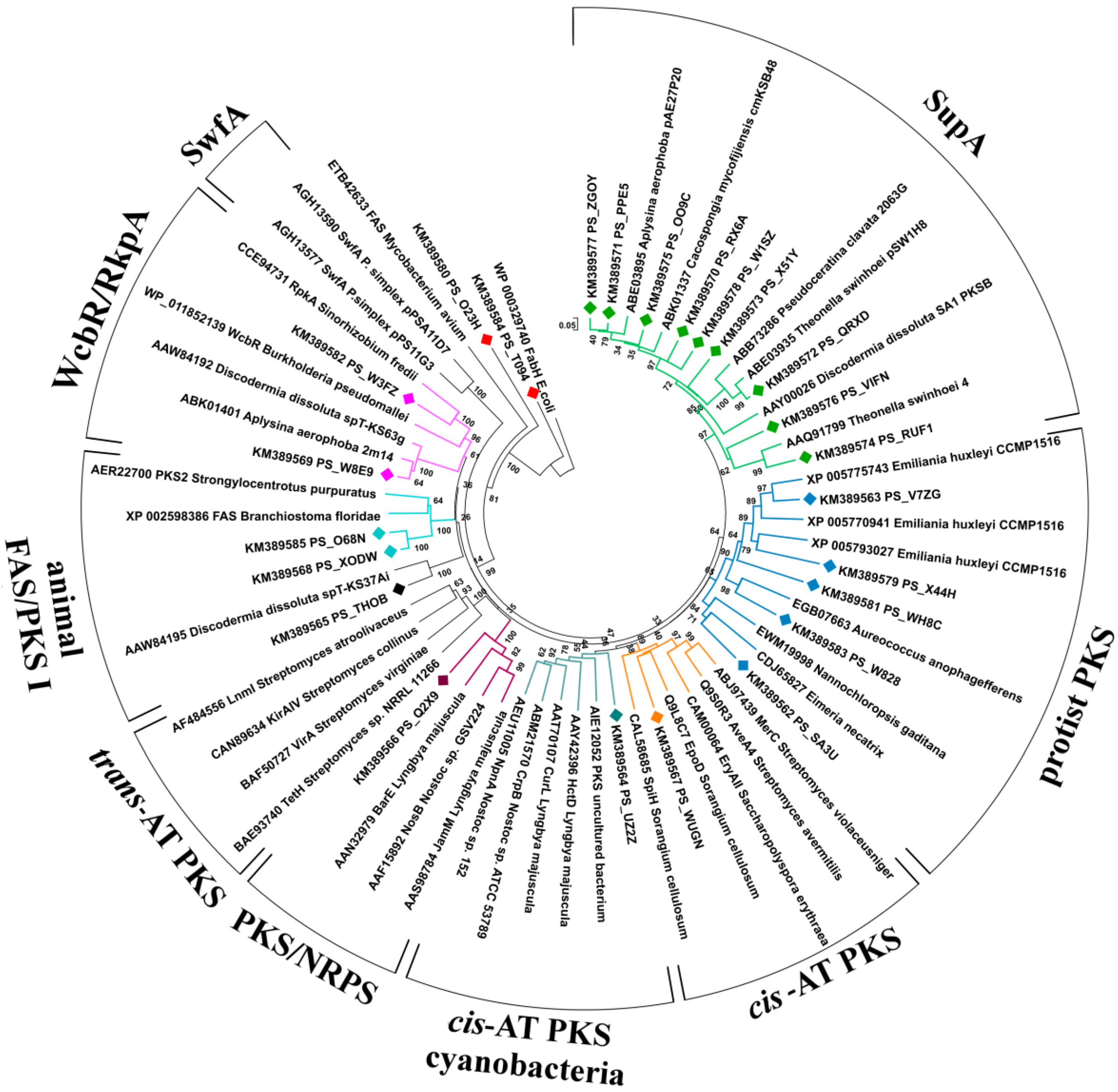
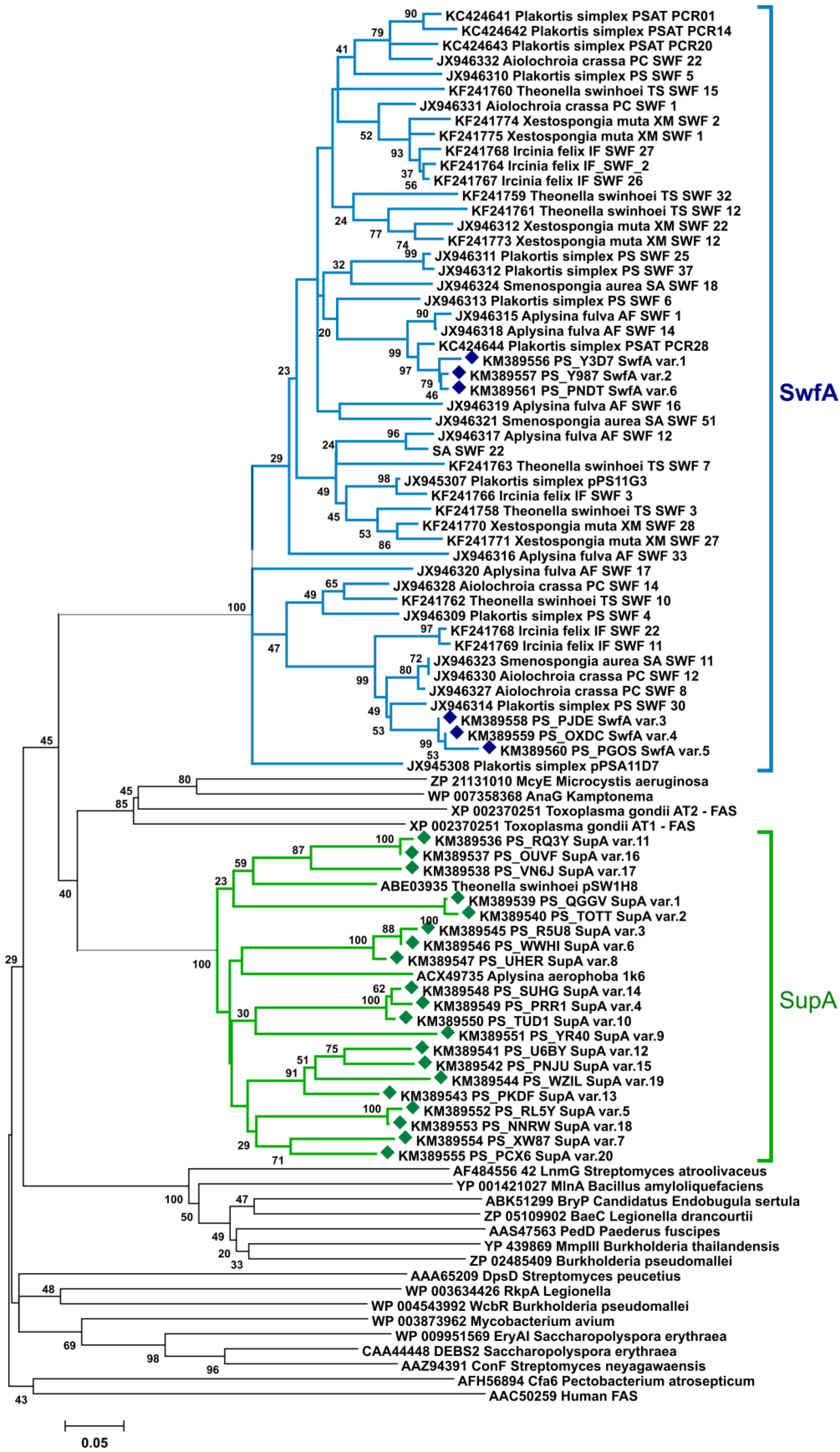
2.1.1. Phylogenetic Analysis of KS Amplicons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | bp | G + C Content (%) | Putative KS Domain Class | BLASTx Closest Homolog (Accession#) Organism | Expect Value | Identity/Positives (% aa) | NaPDos Match |
|---|---|---|---|---|---|---|---|
| PS_Q2X9 | 453 | 64.9 | PKS/NRPS | WP_004354935, Thauera phenylacetica | 2e−76 | 97/98 | JamM (AAS98784), L. majuscula |
| PS_WUGN | 456 | 60.5 | modular cis-AT | AGK63339, uncultured symbiont from A. brasiliensis | 1e−60 | 70/79 | EpoD (Q9L8C7), S. cellulosum |
| PS_UZ2Z | 471 | 55.8 | modular cis-AT | AIE12052, uncultured bacterium from mangrove soil | 1e−56 | 60/75 | CurL (AAT70107), L. majuscula |
| PS_SA3U | 464 | 60.1 | iterative PKS I | EWM19998, Nannochloropsis gaditana | 1e−45 | 54/70 | CALO5 (AAM70355), M. echinospora |
| PS_WH8C | 465 | 59.4 | modular cis-AT (starter KS) | XP_005793027, Emiliania huxleyi CCMP1516 | 1e−42 | 54/69 | CurA (AAT70096), L. majuscula |
| PS_V7ZG | 353 | 65.4 | modular cis-AT | XP_005775743, Emiliania huxleyi CCMP1516 | 8e−48 | 74/84 | MxaD (Q93TW8), S. aurantiaca |
| PS_X44H | 281 | 52.7 | modular cis-AT | XP_005770941, Emiliania huxleyi CCMP1516 | 5e−28 | 58/79 | AveA4 (Q9S0R3), S. avermitilis |
| PS_W828 | 416 | 58.2 | modular cis-AT | EGB07663, Aureococcus anophagefferens | 1e−36 | 56/70 | MxaB (Q93TX0), S. aurantiaca |
2.1.2. Phylogenetic Analysis of AT Amplicons
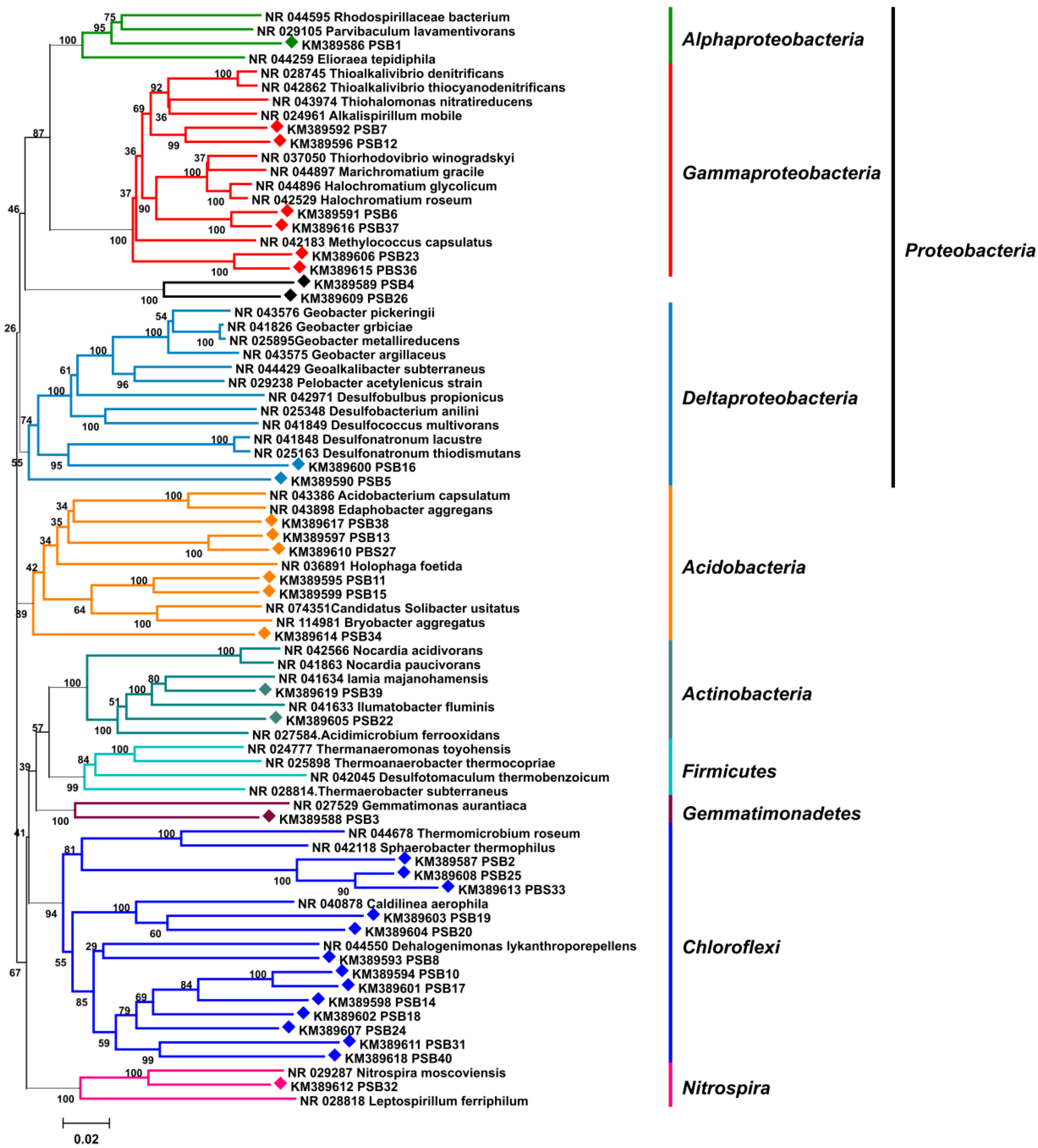
2.2. Sponge-Microbe Associations


3. Discussion
4. Experimental Section
4.1. Sponge Collection
4.2. Metagenomic DNA Isolation
4.3. Amplification of 16S rRNA Gene from P. halichondrioides
4.4. 454 Sequencing of KS and AT Amplicons
4.5. Bioinformatics
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vacelet, J. Étude en microscopie électronique de l’association entre bactéries et spongiaires du genre Verongia (Dictyoceratida). J. Microsc. Biol. Cell. 1975, 23, 271–288. [Google Scholar]
- Laroche, M.; Imperatore, C.; Grozdanov, L.; Costantino, V.; Mangoni, A.; Hentschel, U.; Fattorusso, E. Cellular localisation of secondary metabolites isolated from the Caribbean sponge Plakortis simplex. Mar. Biol. 2007, 151, 1365–1373. [Google Scholar] [CrossRef]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Hopke, J.; Horn, M.; Friedrich, A.B.; Wagner, M.; Hacker, J.; Moore, B.S. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 2002, 68, 4431–4440. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Negri, A.P.; Munro, M.M.H.G.; Battershill, C.N. Diverse microbial communities inhabit Antarctic sponges. Environ. Microbiol. 2004, 6, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Schupp, P.J.; Dahllof, I.; Kjelleberg, S.; Steinberg, P.D. Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity. Environ. Microbiol. 2004, 6, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2009, 26, 338–362. [Google Scholar] [CrossRef] [PubMed]
- Piel, J.; Hui, D.; Wen, G.; Butzke, D.; Platzer, M.; Fusetani, N.; Matsunaga, S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl. Acad. Sci. USA 2004, 101, 16222–16227. [Google Scholar] [CrossRef] [PubMed]
- Fisch, K.M.; Gurgui, C.; Heycke, N.; van der Sar, S.A.; Anderson, S.A.; Webb, V.L.; Taudien, S.; Platzer, M.; Rubio, B.K.; Robinson, S.J.; et al. Polyketide assembly lines of uncultivated sponge symbionts from structure-based gene targeting. Nat. Chem. Biol. 2009, 5, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Holland, N.D.; Faulkner, D.J. Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Cell. Mol. Life Sci. 1996, 52, 716–722. [Google Scholar] [CrossRef]
- Andrianasolo, E.H.; Gross, H.; Goeger, D.; Musafija-Girt, M.; McPhail, K.; Leal, R.M.; Mooberry, S.L.; Gerwick, W.H. Isolation of swinholide A and related glycosylated derivatives from two field collections of marine cyanobacteria. Org. Lett. 2005, 7, 1375–1378. [Google Scholar] [CrossRef] [PubMed]
- Uria, A.; Piel, J. Cultivation-independent approaches to investigate the chemistry of marine symbiotic bacteria. Phytochem. Rev. 2009, 8, 401–411. [Google Scholar] [CrossRef]
- Higgs, M.D.; Faulkner, D.J. Plakortin, an antibiotic from Plakortis halichondrioides. J. Org. Chem. 1978, 43, 3454–3457. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; di Rosa, M.; Ianaro, A. Plakoside A and B, two unique prenylated glycosphingolipids with immunosuppressive activity from the marine sponge Plakortis simplex. J. Am. Chem. Soc. 1997, 119, 12465–12470. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; di Rosa, M.; Ianaro, A. Simplexides, novel immunosuppressive glycolipids from the Caribbean sponge Plakortis simplex. Bioorg. Med. Chem. Lett. 1999, 9, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. Plaxyloside from the marine sponge Plakortis simplex: An improved strategy for NMR structural studies of carbohydrate chains. Eur. J. Org. Chem. 2001, 4457–4462. [Google Scholar]
- Gloeckner, V.; Wehrl, M.; Moitinho-Silva, L.; Gernert, C.; Schupp, P.; Pawlik, J.R.; Lindquist, N.L.; Erpenbeck, D.; Wörheide, G.; Hentschel, U. The HMA-LMA dichotomy revisited: An electron microscopical survey of 56 sponge species. Biol. Bull. 2014, 227, 78–88. [Google Scholar] [PubMed]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. A biosynthetically significant new bacteriohopanoid present in large amounts in the Caribbean sponge Plakortis simplex. Tetrahedron 2001, 57, 4045–4048. [Google Scholar] [CrossRef]
- Della Sala, G.; Hochmuth, T.; Costantino, V.; Teta, R.; Gerwick, W.; Gerwick, L.; Piel, J.; Mangoni, A. Polyketide genes in the marine sponge Plakortis simplex: A new group of mono-modular type I polyketide synthases from sponge symbionts. Environ. Microbiol. Rep. 2013, 5, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Piel, J. A Polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles. Proc. Natl. Acad. Sci. USA 2002, 99, 14002–14007. [Google Scholar]
- Piel, J.; Hui, D.; Fusetani, N.; Matsunaga, S. Targeting modular polyketide synthases with iteratively acting acyltransferases from metagenomes of uncultured bacterial consortia. Environ. Microbiol. 2004, 6, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Fieseler, L.; Hentschel, U.; Grozdanov, L.; Schirmer, A.; Wen, G.; Platzer, M.; Hrvatin, S.; Butzke, D.; Zimmermann, K.; Piel, J. Widespread occurrence and genomic context of unusually small polyketide synthase genes in microbial consortia associated with marine sponges. Appl. Environ. Microbiol. 2007, 73, 2144–2155. [Google Scholar]
- Hochmuth, T.; Piel, J. Polyketide synthases of bacterial symbionts in sponges—Evolution-based applications in natural products research. Phytochemistry 2009, 70, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Kiss, E.; Reuhs, B.L.; Kim, J.S.; Kereszt, A.; Petrovics, G.; Putnoky, P.; Dusha, I.; Carlson, R.W.; Kondorosi, A. The rkpGHI and -J genes are involved in capsular polysaccharide production by Rhizobium meliloti. J. Bacteriol. 1997, 179, 2132–2140. [Google Scholar] [PubMed]
- Parada, M.; Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; Gutierrez, R.; Buendia-Claveria, A.M.; Lei, W.; Margaret, I.; López-Baena, F.J.; Gil-Serrano, A.M.; et al. Sinorhizobium fredii HH103 mutants affected in capsular polysaccharide (KPS) are impaired for nodulation with soybean and Cajanus cajan. Mol. Plant Microbe Interact. 2006, 19, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Donadio, S.; Monciardini, P.; Sosio, M. Polyketide synthases and nonribosomal peptide synthetases: the emerging view from bacterial genomics. Nat. Prod. Rep. 2007, 24, 1073–1109. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, A.; Gadkari, R.; Reeves, C.D.; Ibrahim, F.; DeLong, E.F.; Hutchinson, C.R. Metagenomic analysis reveals diverse polyketide synthase gene clusters in microorganisms associated with the marine sponge Discodermia dissoluta. Appl. Environ. Microbiol. 2005, 71, 4840–4849. [Google Scholar] [CrossRef] [PubMed]
- Ziemert, N.; Podell, S.; Penn, K.; Badger, J.H.; Allen, E.; Jensen, P.R. The natural product domain seeker NaPDoS: A phylogeny based bioinformatic tool to classify secondary metabolite gene diversity. PLoS One 2012, 7, e34064. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Stüken, A.; Orr, R.J.S.; Svendsen, H.M.; Jakobsen, K.S. Biosynthesis and molecular genetics of polyketides in marine dinoflagellates. Mar. Drugs 2010, 8, 1011–1048. [Google Scholar] [CrossRef] [PubMed]
- John, U.; Beszteri, B.; Derelle, E.; van de Peer, Y.; Read, B.; Moreau, H.; Cembella, A. Novel insights into evolution of protistan polyketide synthases through phylogenomic analysis. Protist 2008, 159, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The ribosomal database project: Improved alignments and new tools for rRNA analysis. Nucl. Acids Res. 2009, 37, (Database issue). D141–D145. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microb. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef]
- Hochmuth, T.; Niederkruger, H.; Gernert, C.; Siegl, A.; Taudien, S.; Platzer, M.; Crews, P.; Hentschel, U.; Piel, J. Linking chemical and microbial diversity in marine sponges: Possible role for poribacteria as producers of methyl-branched fatty acids. Chembiochem 2010, 11, 2572–2578. [Google Scholar] [CrossRef] [PubMed]
- Siegl, A.; Kamke, J.; Hochmuth, T.; Piel, J.; Richter, M.; Liang, C.; Dandekar, T.; Hentschel, U. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J. 2011, 5, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Kamke, J.; Sczyrba, A.; Ivanova, N.; Schwientek, P.; Rinke, C.; Mavromatis, K.; Woyke, T.; Hentschel, U. Single-cell genomics reveals complex carbohydrate degradation patterns in poribacterial symbionts of marine sponges. ISME J. 2013, 7, 2287–2300. [Google Scholar] [CrossRef] [PubMed]
- Kamke, J.; Rinke, C.; Schwientek, P.; Mavromatis, K.; Ivanova, N.; Sczyrba, A.; Woyke, T.; Hentschel, U. The candidate phylum Poribacteria by single-cell genomics: New insights into phylogeny, cell-compartmentation, eukaryote-like repeat proteins, and other genomic features. PLoS One 2014, 9, e87353. [Google Scholar] [CrossRef] [PubMed]
- Jenke-Kodama, H.; Sandmann, A.; Müller, R.; Dittmann, E. Evolutionary implications of bacterial polyketide synthases. Mol. Biol. Evol. 2005, 22, 2027–2039. [Google Scholar] [CrossRef] [PubMed]
- The Sponge Guide. Available online: http://www.spongeguide.org (accessed on 7 July 2010).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Sala, G.; Hochmuth, T.; Teta, R.; Costantino, V.; Mangoni, A. Polyketide Synthases in the Microbiome of the Marine Sponge Plakortis halichondrioides: A Metagenomic Update. Mar. Drugs 2014, 12, 5425-5440. https://doi.org/10.3390/md12115425
Della Sala G, Hochmuth T, Teta R, Costantino V, Mangoni A. Polyketide Synthases in the Microbiome of the Marine Sponge Plakortis halichondrioides: A Metagenomic Update. Marine Drugs. 2014; 12(11):5425-5440. https://doi.org/10.3390/md12115425
Chicago/Turabian StyleDella Sala, Gerardo, Thomas Hochmuth, Roberta Teta, Valeria Costantino, and Alfonso Mangoni. 2014. "Polyketide Synthases in the Microbiome of the Marine Sponge Plakortis halichondrioides: A Metagenomic Update" Marine Drugs 12, no. 11: 5425-5440. https://doi.org/10.3390/md12115425
APA StyleDella Sala, G., Hochmuth, T., Teta, R., Costantino, V., & Mangoni, A. (2014). Polyketide Synthases in the Microbiome of the Marine Sponge Plakortis halichondrioides: A Metagenomic Update. Marine Drugs, 12(11), 5425-5440. https://doi.org/10.3390/md12115425