
Structure of an Amino Acid-Decorated Exopolysaccharide Secreted by a Vibrio alginolyticus Strain

 ,
,
Abstract
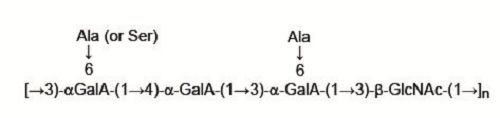
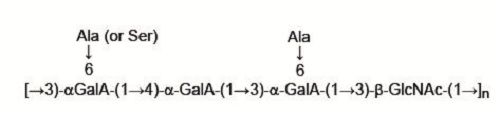
:
1. Introduction
2. Results and Discussion
2.1. Purification and Composition of the EPSs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Sugar | Partially Methylated Alditol Acetate | Deduced Linkage |
|---|---|---|
| Galactose | 1,3,5-Tri-O-acetyl-1-deuterio-2,4,6-tri-O-methyl-d-Galactitol | →3)-Galp-(1→ |
| 1,4,5-Tri-O-acetyl-1-deuterio-2,3,6-tri-O-methyl-d-Galactitol | →4)-Galp-(1→ | |
| 1,3,5,6-Tetra-O-acetyl-1-deuterio-3,4-di-O-methyl-d-Galactitol | →3,6)-Galp-(1→ | |
| N-acetyl Glucosamine | 1,3,5-Tri-O-acetyl-2-(acetylmethylamino)-2-deoxy-1-deuterio-4,6-di-O-methyl-d-Galactitol | →3)-GlcNAc-(1→ |


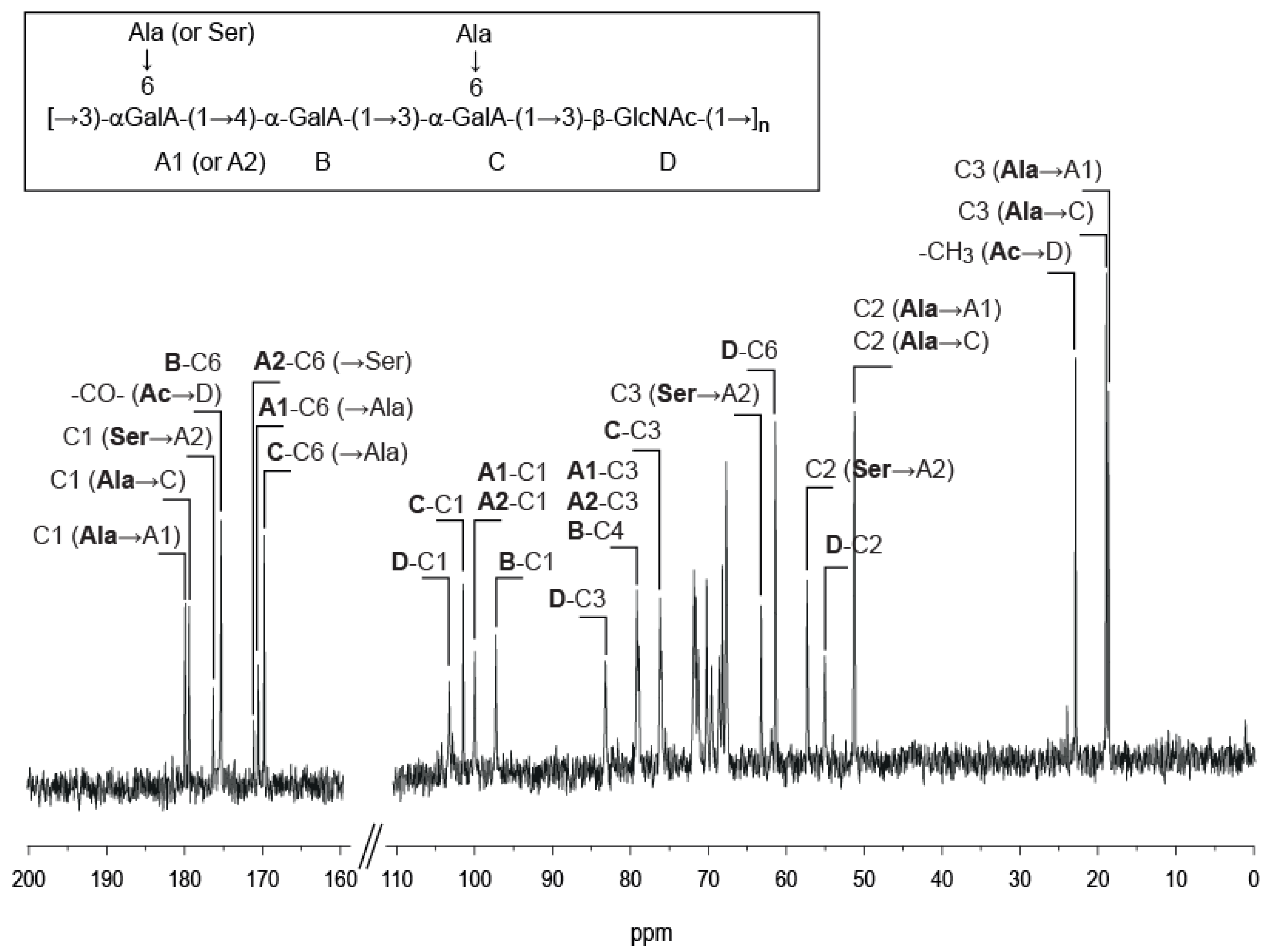
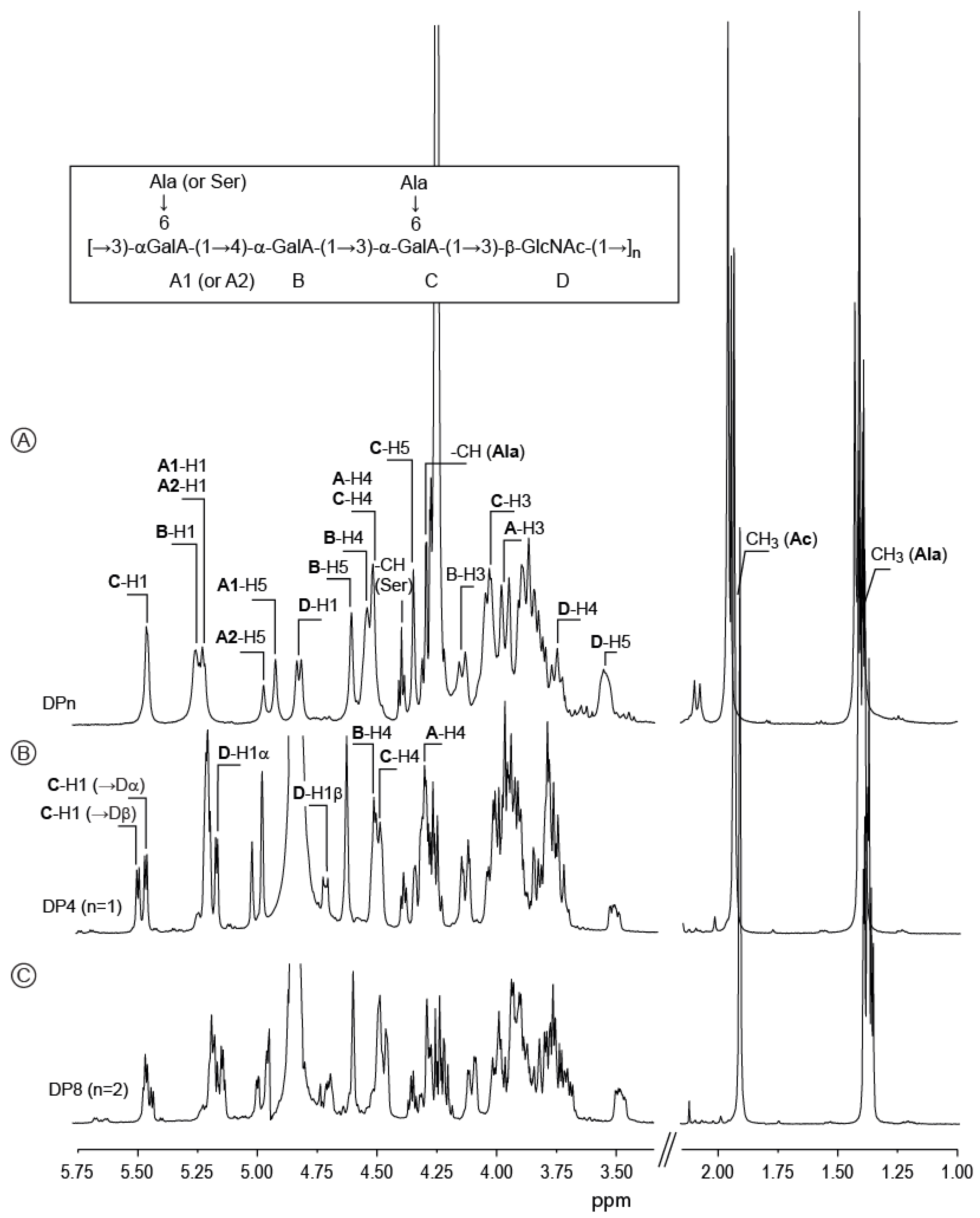
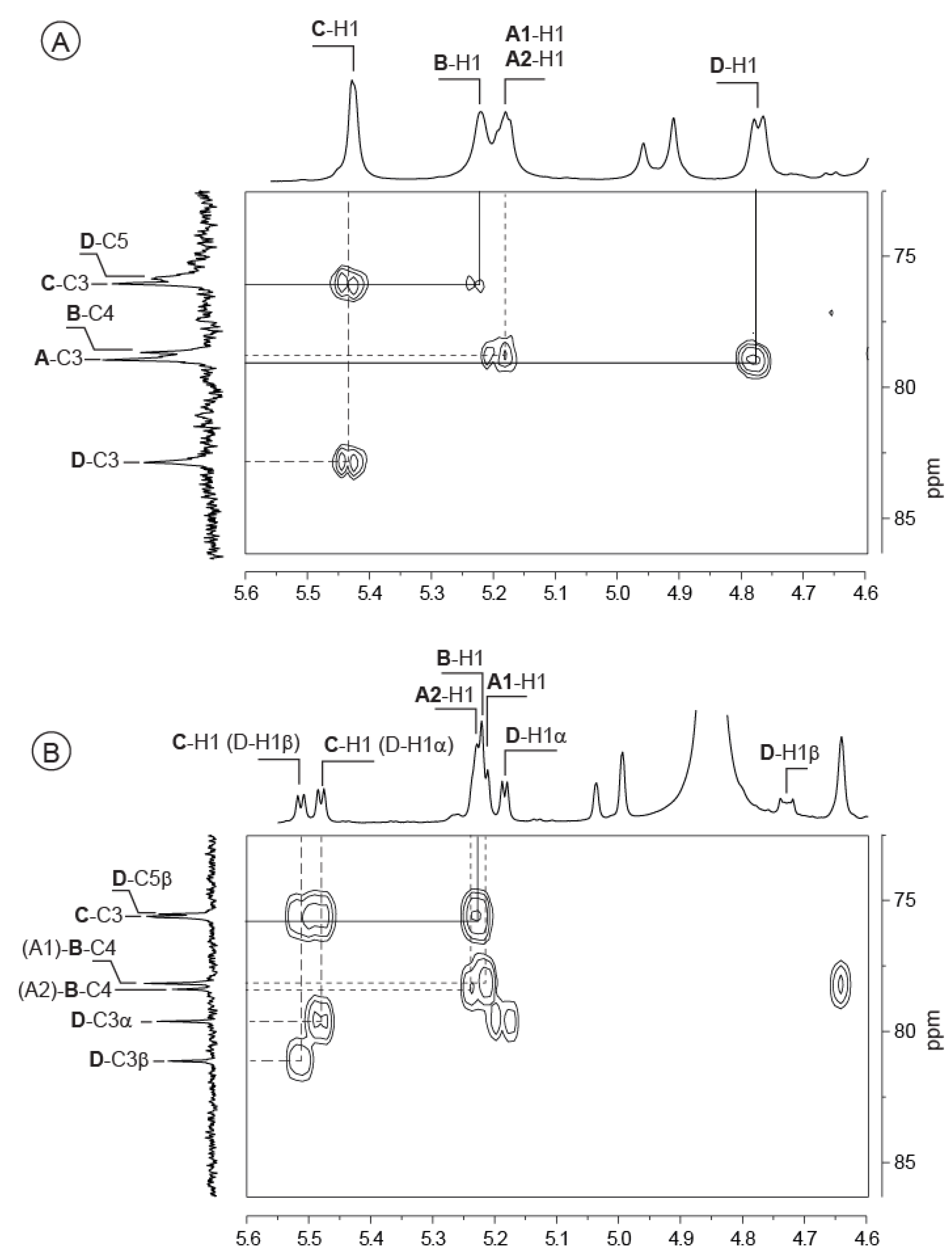
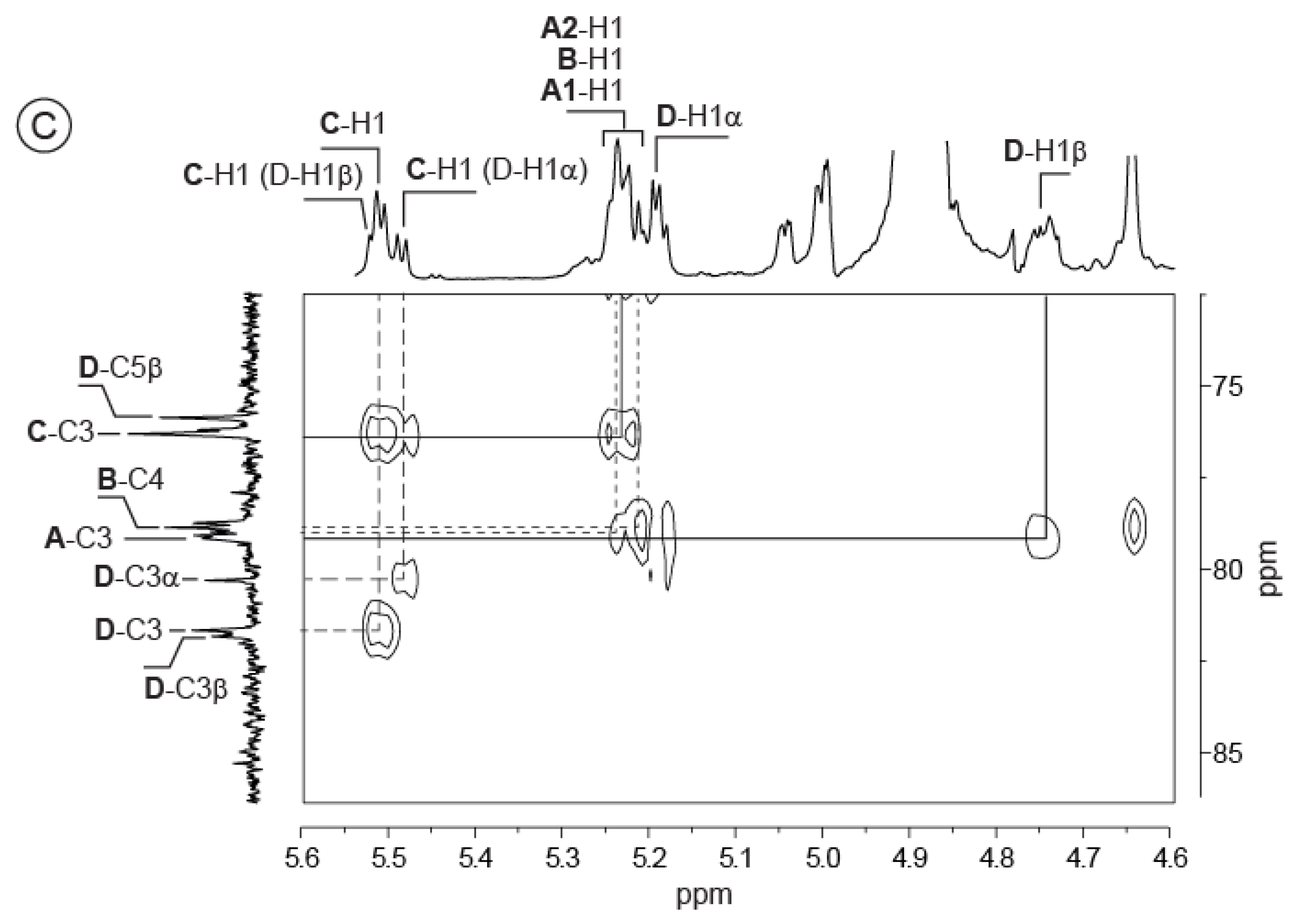
2.2. Detailed Structure Analysis of the EPS Using NMR
| VA-EPS: [ABCD]n | DP4: AnrBCr′Dr | DP8: AnrBCDABCr′Dr | |
|---|---|---|---|
| A1: 6Ala-GalA/A2:6Ser-GalA | |||
| H1 | 5.18/5.19 | 5.20/5.20 | |
| H2 | 3.84/3.84 | 3.80/3.80 | |
| H3 | 4.00/4.00 | 4.04/4.04 | |
| H4 | 4.49/4.51 | 4.54/4.54 | |
| H5 | 4.91/4.96 | 4.99/5.04 | |
| A1nr: 6Ala-GalA/A2nr:6Ser-GalA | |||
| H1 | 5.22/5.23 | 5.20/5.22 | |
| H2 | 3.78/3.78 | 3.80/3.80 | |
| H3 | 3.97/3.99 | 3.99/3.99 | |
| H4 | 4.33/4.36 | 4.34/4.34 | |
| H5 | 4.99/5.04 | 4.99/5.04 | |
| B: GalA | |||
| H1 | 5.22 | 5.22 | 5.23 |
| H2 | 3.93 | 3.93 | 3.95 |
| H3 | 4.10 | 4.14 | 4.15 |
| H4 | 4.51 | 4.52 | 4.53 |
| H5 | 4.58 | 4.64 | 4.65 |
| C: 6Ala-GalA | |||
| H1 | 5.43 | 5.50 | |
| H2 | 3.97 | 3.98 | |
| H3 | 4.03 | 4.02 | |
| H4 | 4.48 | 4.50 | |
| H5 | 4.31 | 4.32 | |
| Cr′: 6Ala-GalA | |||
| H1-Dr α/β | 5.48/5.51 | 5.48/5.51 | |
| H2-Dr α/β | 3.98/3.97 | 3.98/3.97 | |
| H3-Dr α/β | 4.02/4.00 | 4.03/4.00 | |
| H4 | 4.50 | 4.50 | |
| H5 | 4.31 | 4.32 | |
| D: GlcNAc | |||
| H1 | 4.77 | 4.75 | |
| H2 | 3.85 | 3.87 | |
| H3 | 3.81 | 3.87 | |
| H4 | 3.70 | 3.75 | |
| H5 | 3.51 | 3.53 | |
| H6′ | 3.93 | 3.95 | |
| H6′ | 3.77 | 3.82 | |
| NHAc:CH3 | 1.95 | 1.95 | |
| Dr: GlcNAc | |||
| H1 α/β | 5.18/4.73 | 5.18/4.73 | |
| H2 α/β | 4.03/3.80 | 4.03/3.81 | |
| H3 α/β | 3.99/3.81 | 4.00/3.81 | |
| H4 α/β | 3.75/3.73 | 3.75/3.73 | |
| H5 α/β | 3.92/3.52 | 3.93/3.53 | |
| H6 α/β | 3.84/3.91 | 3.95/3.82 | |
| H6′ | 3.78 | 3.82 | |
| NHAc: CH3 | 1.95 | 1.95 | |
| VA-EPS: [ABCD]n | DP4: AnrBCr′Dr | DP8: AnrBCDABCr′Dr | |
|---|---|---|---|
| A1: 6Ala-GalA/A2: 6Ser-GalA | |||
| C1 | 99.82/99.94 | 100.07/100.22 | |
| C2 | 67.99/67.99 | 68.81/68.81 | |
| C3 | 78.97/78.97 | 79.14/79.23 | |
| C4 | 70.12/70.12 | 70.34/70.34 | |
| C5 | 71.43/71.60 | 71.92/72.02 | |
| C6 | 170.50/171.05 | 170.94/171.37 | |
| A1nr: 6Ala-GalA/A2nr: 6Ser-GalA | |||
| C1 | 100.07/100.22 | 100.07/100.22 | |
| C2 | 68.84/68.84 | 68.81/68.81 | |
| C3 | 69.78/69.78 | 69.76/69.76 | |
| C4 | 70.47/70.43 | 70.47/70.43 | |
| C5 | 71.93/72.04 | 71.92/72.04 | |
| C6 | 171.10/171.54 | 171.10/171.54 | |
| B: GalA | |||
| C1 | 97.20 | 97.63 | 97.66 |
| C2 | 68.33 | 68.34 | 68.34 |
| C3 | 69.34 | 69.25 | 69.25 |
| C4-A1/A2 | 78.68/78.85 | 78.93/79.15 | 78.80/78.93 |
| C5 | 71.82 | 71.73 | 71.73 |
| C6 | 175.34 | 175.80 | 175.83 |
| C: 6Ala-GalA | |||
| C1 | 101.32 | 101.06 | |
| C2 | 67.40 | 67.27 | |
| C3 | 76.06 | 76.37 | |
| C4 | 67.61 | 67.92 | |
| C5 | 71.72 | 71.66 | |
| C6 | 169.68 | 170.05 | |
| Cr′: 6Ala-GalA | |||
| C1-Dr α/β | 101.39/101.04 | 101.37/101.06 | |
| C2-Dr α/β | 67.28/67.44 | 67.27/67.44 | |
| C3-Dr α/β | 76.41/76.36 | 76.37/76.26 | |
| C4-Dr α/β | 67.90/67.84 | 67.92/67.86 | |
| C5-Dr α/β | 71.67/71.73 | 71.51/71.66 | |
| C6-Dr α/β | 170.01/170.14 | 170.05/170.15 | |
| D: GlcNAc | |||
| C1 | 103.17 | 103.55 | |
| C2 | 54.84 | 54.85 | |
| C3 | 82.89 | 81.74 | |
| C4 | 71.03 | 70.93 | |
| C5 | 75.87 | 75.93 | |
| C6 | 61.13 | 60.91 | |
| NHAc: CH3/CO | 22.68/175.25 | 22.76/175.64 | |
| Dr: GlcNAc | |||
| C1 α/β | 91.76/95.73 | 91.76/95.73 | |
| C2 α/β | 53.11/55.80 | 53.11/55.80 | |
| C3 α/β | 80.37/81.89 | 80.37/81.90 | |
| C4 α/β | 71.09 | 71.08 | |
| C5 α/β | 72.15/76.28 | 72.15/76.26 | |
| C6 α/β | 61.02/61.17 | 61.01/61.14 | |
| NHAc: CO | 174.90/175.20 | 174.88/175.19 | |
| NHAc: CH3 | 22.44/22.70 | 22.46/22.72 |


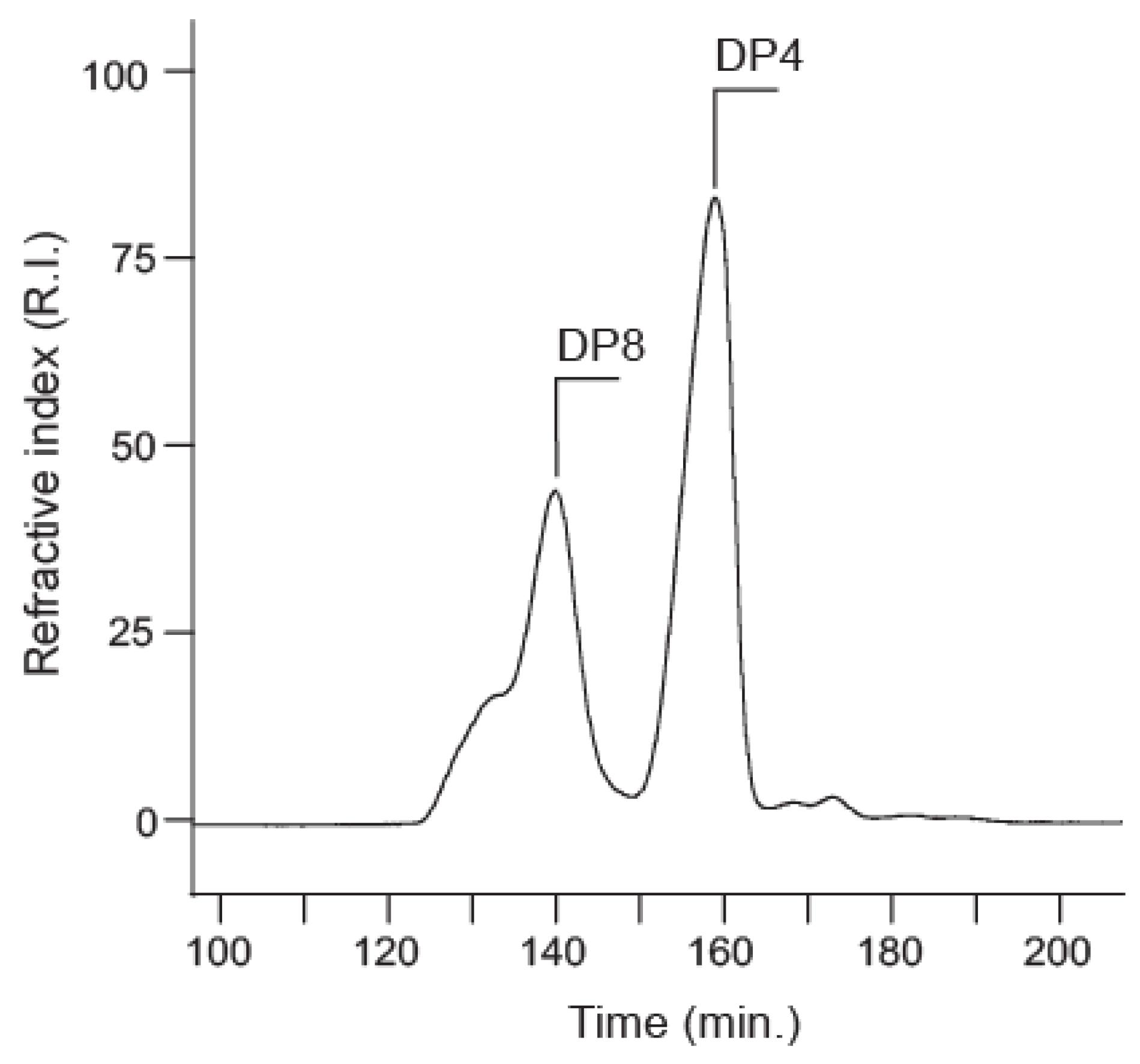
2.3. Preparation and Analysis of Standard Oligosaccharides

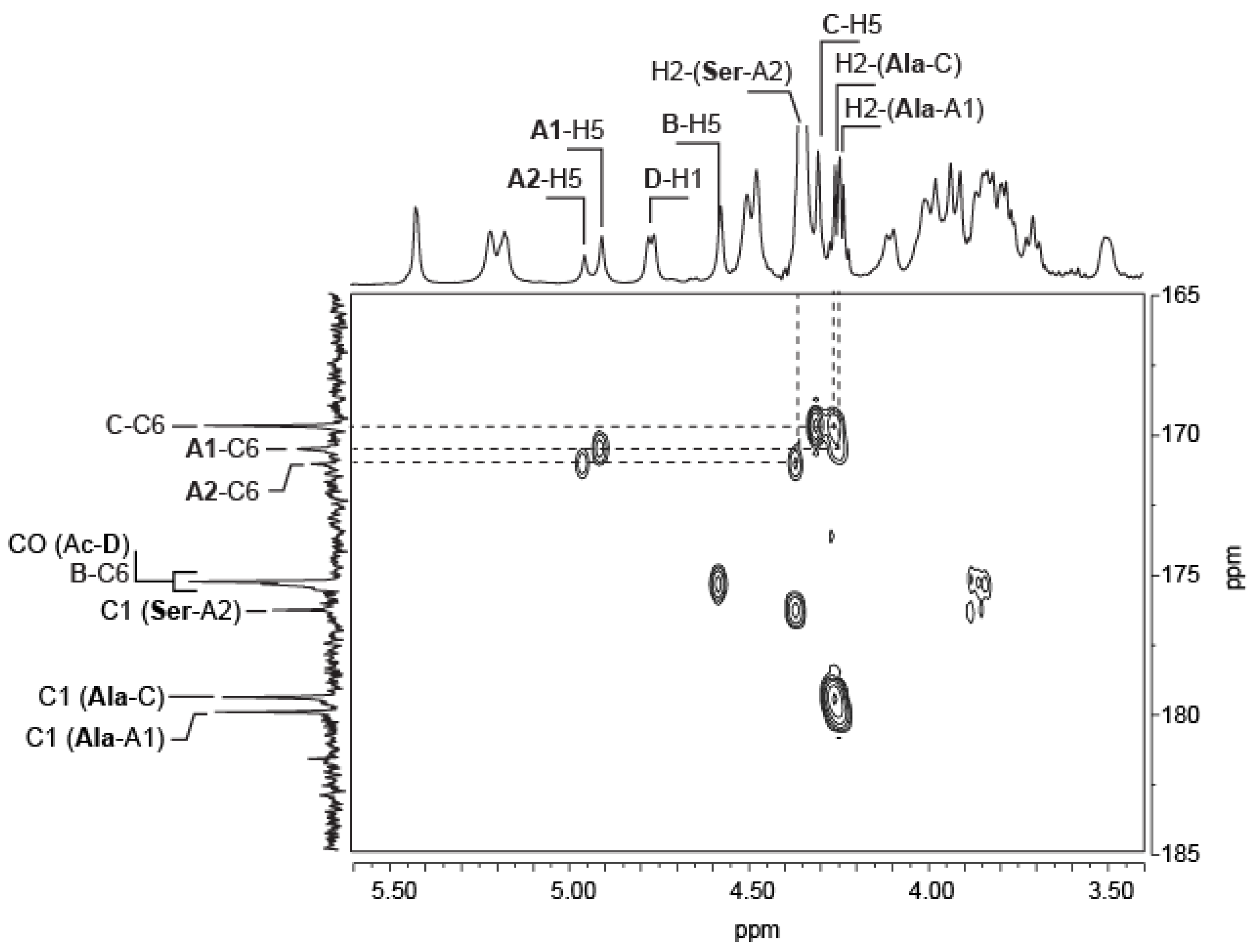
2.4. Position and Distribution of d-Alanine and d-Serine
| VA-EPS: [ABCD]n | DP4: AnrBCr'Dr | DP8: AnrBCDABCr′Dr | |
|---|---|---|---|
| Ala-A1 | |||
| H2 | 4.25 | 4.27 | 4.26 |
| H3 | 1.37 | 1.41 | 1.41 |
| C1 | 179.92 | 180.44 | 180.46 |
| C2 | 51.09 | 51.29 | 51.31 |
| C3 | 18.44 | 18.43 | 18.40/18.35 |
| Ala-C | |||
| H2 | 4.27 | 4.29 | 4.28 |
| H3 | 1.38 | 1.42 | 1.42 |
| C1 | 179.38 | 179.95/179.92 | 179.96/179.93 |
| C2 | 51.09 | 51.29 | 51.31 |
| C3 | 18.91 | 18.71/18.58 | 18.69 |
| Ser-A2 | |||
| H2 | 4.37 | 4.40 | 4.40 |
| H3 | 3.85 | 3.91 | 3.91 |
| C1 | 176.26 | 176.66 | 176.68 |
| C2 | 57.19 | 57.35 | 57.37 |
| C3 | 62.99 | 62.96 | 62.94 |

3. Experimental Section
3.1. Production, Isolation and Purification of VA-EPS Exopolysaccharide
3.2. Monosaccharide Analysis
3.3. Methylation Analysis
3.4. Amino Acid Composition
3.5. Sulfate Content
3.6. Molecular Weight Determination
3.7. Acid Hydrolysis and Oligosaccharides Purification
3.8. NMR
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sutherland, I.W. Microbial polysaccharides from gram-negative bacteria. Int. Dairy J. 2001, 11, 663–674. [Google Scholar] [CrossRef]
- Decho, A.W. Microbial exoplomymer secretion in ocean environments: Their role(s) in food webs and marine processes. Oceanogr. Mar. Biol. Annu. Rev. 1990, 28, 73–153. [Google Scholar]
- Arena, A.; Maugeri, T.L.; Pavone, B.; Iannello, D.; Gugliandolo, C.; Bisignanoc, G. Antiviral and immunoregulatory effect of a novel exopolysaccharide from a marine thermotolerant Bacillus licheniformis. Int. Immunopharmacol. 2006, 6, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Zanchetta, P.; Lagarde, N.; Guezennec, J. Systemic effects on bone healing of a new hyaluronic acid-like bacterial exopolysaccharide. Calcif. Tissue Int. 2003, 73, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Courtois, A.; Berthou, C.; Guézennec, J.; Boisset, C.; Bordron, A. Exopolysaccharides isolated from hydrothermal vent bacteria can modulate the complement system. PLoS ONE 2014, 9, e94965. [Google Scholar] [CrossRef] [PubMed]
- Freitas, F.; Alves, V.D.; Reis, M.A.M. Advances in bacterial exopolysaccharides: From production to biotechnological applications. Trends Biotechnol. 2011, 29, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Gedouin, A.; Vallee, R. Compositions à Visée Cosmétique d’exopolysaccharides Issus de Bactéries Marines. Patent FR_2975906_A1, 2012. [Google Scholar]
- Nishibuchi, M.; Kaper, J.B. Thermostable direct hemolysin gene of Vibrio parahaemolyticus: A virulence gene acquired by a marine bacterium. Infect. Immun. 1995, 63, 2093–2099. [Google Scholar] [PubMed]
- Kamerling, J.P.; Gerwig, G.J.; Vliegenthart, J.F.; Clamp, J.R. Characterization by gas-liquid chromatography-mass spectrometry and proton-magnetic-resonance spectroscopy of pertrimethylsilyl methyl glycosides obtained in the methanolysis of glycoproteins and glycopeptides. Biochem. J. 1975, 151, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Montreuil, J.; Bouquelet, S.; Debray, H.; Fournet, B.; Spik, G.; Strecker, G. Glycoproteines. In Carbohydrates Analysis: A Practical Approach; Chaplin, M.F., Kennedy, J.K., Eds.; IRL Press: Oxford, UK, 1986; pp. 143–204. [Google Scholar]
- Hakomori, S.I. A rapid permethylation of glycolipid, and polysaccharide catalyzed by methylsulfinyl carbanion in dimethyl sulfoxide. J. Biochem. 1964, 55, 205–208. [Google Scholar] [PubMed]
- Taylor, R.L.; Conrad, H.E. Stoichiometric depolymerization of polyuronides and glycosaminoglycans to monosaccharides following reduction of their carbodiimide activated carboxyl groups. Biochemistry 1972, 11, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Kvernheim, A.L. Methylation analysis of polysaccharides with butyllithium in dimethyl sulfoxide. Acta Chem. Scand. 1987, B41, 150–152. [Google Scholar] [CrossRef]
- Linnerborg, M.; Wollin, R.; Widmalm, G. Structural studies of the O-antigen polysaccharide from Escherichia coli O167. Eur. J. Biochem. 1997, 246, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Knirel, Y.A.; Feng, L.; Perepelov, A.V.; Senchenkova, S.N.; Wang, Q.; Reeves, P.R.; Wang, L. Structure and genetics of Shigella O antigens. FEMS Microbiol. Rev. 2008, 32, 627–653. [Google Scholar] [CrossRef] [PubMed]
- Knirel, Y.A.; Perepelov, A.V.; Kondakova, A.N.; Senchenkova, S.N.; Sidorczyk, Z.; Rozalski, A.; Kaca, W. Structure and serology of O-antigens as the basis for classification of Proteus strains. Innate Immun. 2011, 17, 70–96. [Google Scholar] [CrossRef] [PubMed]
- Jann, B.; Kochanowski, H.; Jann, K. Structure of the capsular K96 polysaccharide (K96 antigen) from Escherichia coli O77:K96:H and comparison with the capsular K54 polysaccharide (K54 antigen) from Escherichia coli O6:K54:H10. Carbohydr. Res. 1994, 253, 323–327. [Google Scholar] [CrossRef]
- Gorshkova, R.P.; Isakov, V.V.; Nedashkovskaya, O.I.; Nazarenko, E.L. Structure of carbohydrate antigens from Microbulbifer sp. KMM 6242. Chem. Nat. Compd. 2011, 46, 837–840. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drouillard, S.; Jeacomine, I.; Buon, L.; Boisset, C.; Courtois, A.; Thollas, B.; Morvan, P.-Y.; Vallée, R.; Helbert, W. Structure of an Amino Acid-Decorated Exopolysaccharide Secreted by a Vibrio alginolyticus Strain. Mar. Drugs 2015, 13, 6723-6739. https://doi.org/10.3390/md13116723
Drouillard S, Jeacomine I, Buon L, Boisset C, Courtois A, Thollas B, Morvan P-Y, Vallée R, Helbert W. Structure of an Amino Acid-Decorated Exopolysaccharide Secreted by a Vibrio alginolyticus Strain. Marine Drugs. 2015; 13(11):6723-6739. https://doi.org/10.3390/md13116723
Chicago/Turabian StyleDrouillard, Sophie, Isabelle Jeacomine, Laurine Buon, Claire Boisset, Anthony Courtois, Bertrand Thollas, Pierre-Yves Morvan, Romuald Vallée, and William Helbert. 2015. "Structure of an Amino Acid-Decorated Exopolysaccharide Secreted by a Vibrio alginolyticus Strain" Marine Drugs 13, no. 11: 6723-6739. https://doi.org/10.3390/md13116723
APA StyleDrouillard, S., Jeacomine, I., Buon, L., Boisset, C., Courtois, A., Thollas, B., Morvan, P. -Y., Vallée, R., & Helbert, W. (2015). Structure of an Amino Acid-Decorated Exopolysaccharide Secreted by a Vibrio alginolyticus Strain. Marine Drugs, 13(11), 6723-6739. https://doi.org/10.3390/md13116723