
Activation of the Tumor Suppressor PP2A Emerges as a Potential Therapeutic Strategy for Treating Prostate Cancer

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
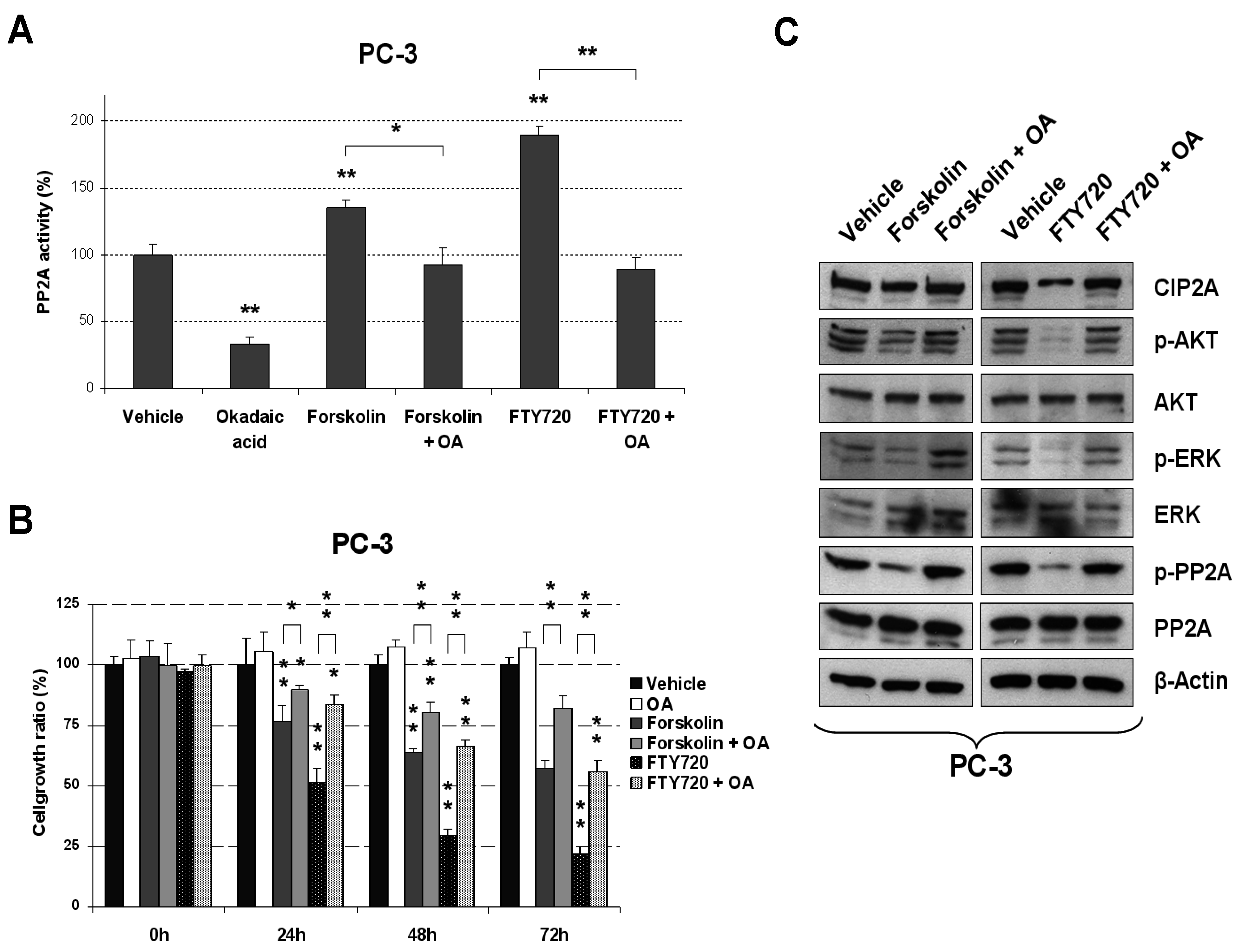
2.1. Forskolin and FTY720 Lead to Reduced Cell Viability in PCa Cells That Is Dependent on PP2A Activation
2.2. Molecular Effects of PP2A Activation in PCa Cells after Forskolin or FTY720 Treatments

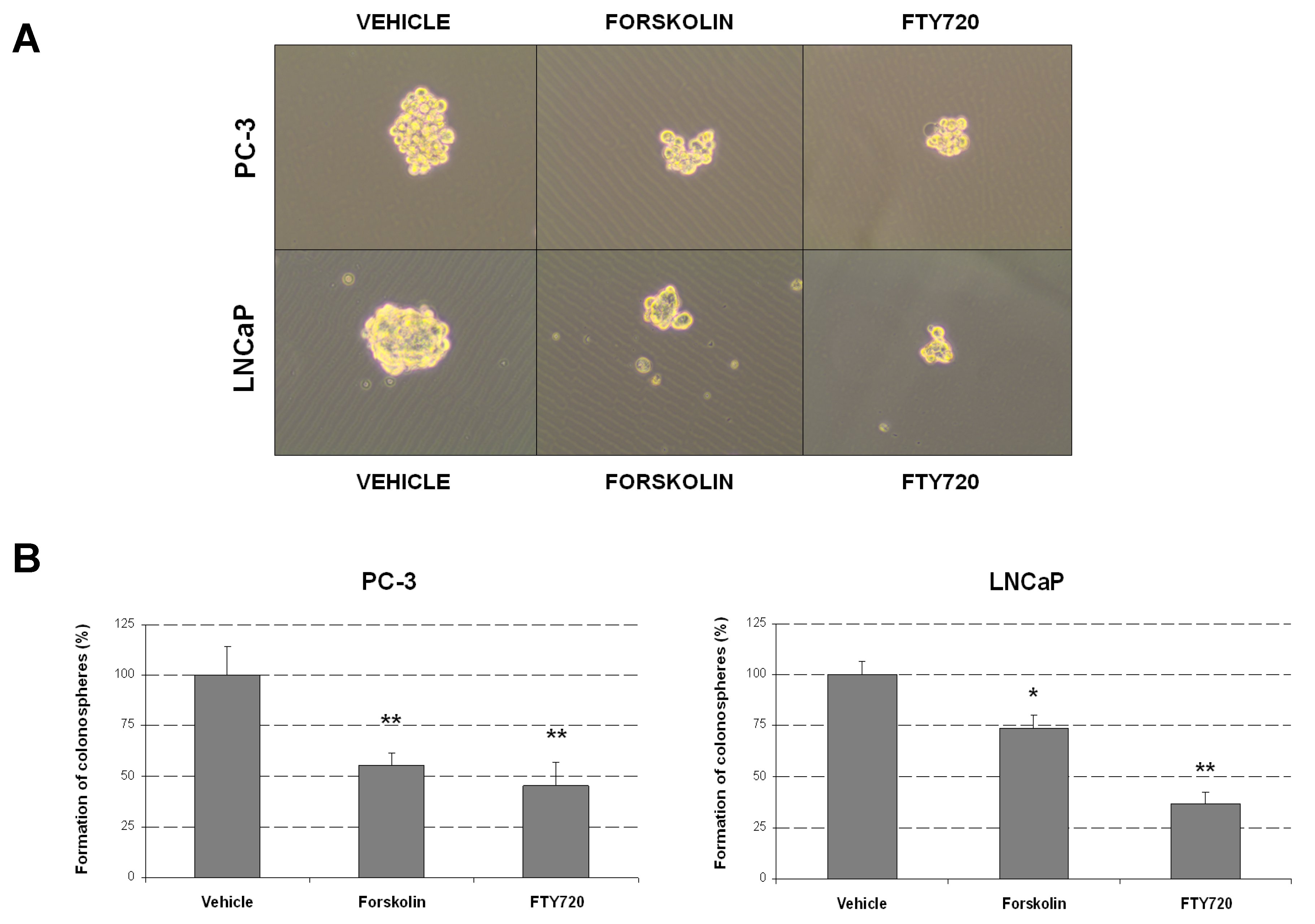
2.3. Forskolin- and FTY720-Induced PP2A Activation Impair Prostasphere Formation

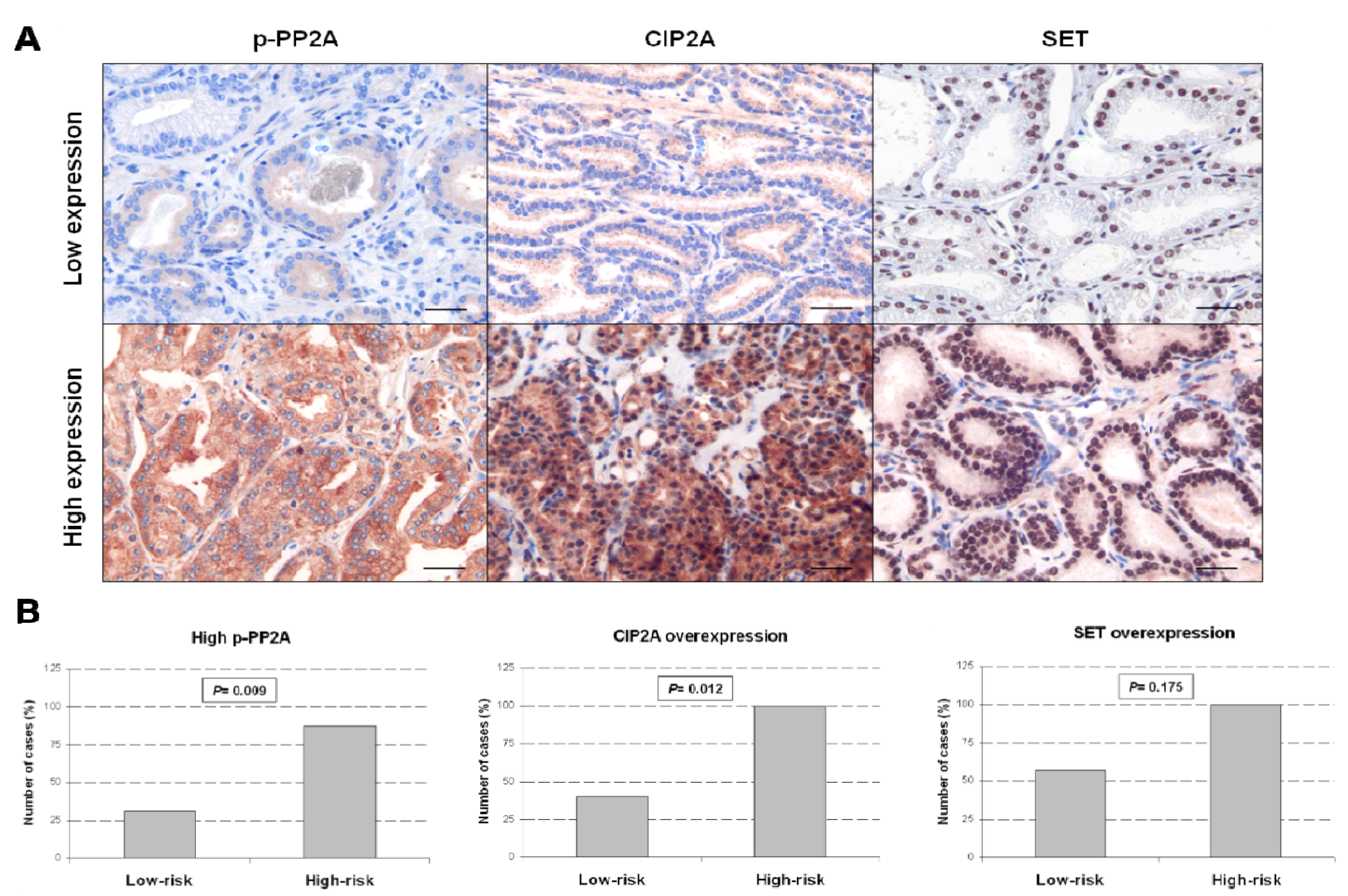
2.4. Deregulation of CIP2A and p-PP2A Are Associated with Aggressive Prostate Tumors

3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Patient Samples
4.3. Western Blot Analysis
4.4. Proliferation Assay and Cell Viability
4.5. PP2A Phosphatase Activity Assays
4.6. Prostaspheres
4.7. Quantitative Real-Time RT-PCR
4.8. Immunohistochemistry
4.9. Statistical Analysis
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Westermarck, J.; Hahn, W.C. Multiple pathways regulated by the tumor suppressor PP2A in transformation. Trends Mol. Med. 2008, 14, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Mumby, M. PP2A: Unveiling a reluctant tumor suppressor. Cell 2007, 130, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Janssens, V.; Goris, J.; van Hoof, C. PP2A: The expected tumor suppressor. Curr. Opin. Genet. Dev. 2005, 15, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn, P.J.; Creyghton, M.P.; Bernards, R. Protein phosphatase 2A regulatory subunits and cancer. Biochim. Biophys. Acta 2009, 1795, 1–15. [Google Scholar] [PubMed]
- Bhardwaj, A.; Singh, S.; Srivastava, S.K.; Arora, S.; Hyde, S.J.; Andrews, J.; Grizzle, W.E.; Singh, A.P. Restoration of PPP2CA expression reverses epithelial-to-mesenchymal transition and suppresses prostate tumour growth and metastasis in an orthotopic mouse model. Br. J. Cancer 2014, 110, 2000–2010. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Singh, S.; Srivastava, S.K.; Honkanen, R.E.; Reed, E.; Singh, A.P. Modulation of protein phosphatase 2A activity alters androgen-independent growth of prostate cancer cells: Therapeutic implications. Mol. Cancer Ther. 2011, 10, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Seshacharyulu, P.; Das, S.; Rachagani, S.; Ponnusamy, M.P.; Yan, Y.; Johansson, S.L.; Datta, K.; Fong Lin, M.; Batra, S.K. Impaired expression of protein phosphatase 2A subunits enhances metastatic potential of human prostate cancer cells through activation of AKT pathway. Br. J. Cancer 2013, 108, 2590–2600. [Google Scholar] [CrossRef] [PubMed]
- Bluemn, E.G.; Spencer, E.S.; Mecham, B.; Gordon, R.R.; Coleman, I.; Lewinshtein, D.; Mostaghel, E.; Zhang, X.; Annis, J.; Grandori, C.; et al. PPP2R2C loss promotes castration-resistance and is associated with increased prostate cancer-specific mortality. Mol. Cancer Res. 2013, 11, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Vaarala, M.H.; Väisänen, M.R.; Ristimäki, A. CIP2A expression is increased in prostate cancer. J. Exp. Clin. Cancer Res. 2010, 29, 136. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Liu, D.; Su, Z. CIP2A mediates prostate cancer progression via the c-MYC signaling pathway. Tumour Biol. 2015, in press. [Google Scholar]
- Huang, J.; Jia, J.; Tong, Q.; Liu, J.; Qiu, J.; Sun, R.; Yao, L.; Yang, C. Knockdown of cancerous inhibitor of protein phosphatase 2A may sensitize metastatic castration-resistant prostate cancer cells to cabazitaxel chemotherapy. Tumour Biol. 2014, 36, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, N.M.; Hovens, C.M.; Michael, M.; Rosenthal, M.A.; Costello, A.J. Open-label, phase I dose-escalation study of sodium selenate, a novel activator of PP2A, in patients with castration-resistant prostate cancer. Br. J. Cancer 2010, 103, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Kim, H.J.; Chun, Y.J.; Kim, M.Y. Ceramide produces apoptosis through induction of p27(KIP1) by protein phosphatase 2A-dependent Akt dephosphorylation in PC-3 prostate cancer cells. J. Toxicol. Environ. Health A 2010, 73, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Palit, S.; Ball, W.B.; Das, P.K. Carnosic acid modulates Akt/IKK/NF-κB signaling by PP2A and induces intrinsic and extrinsic pathway mediated apoptosis in human prostate carcinoma PC-3 cells. Apoptosis 2012, 17, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Cirak, Y.; Varol, U.; Atmaca, H.; Kisim, A.; Sezgin, C.; Karabulut, B.; Uzunoglu, S.; Uslu, R.; Karaca, B. Zoledronic acid in combination with serine/threonine phosphatase inhibitors induces enhanced cytotoxicity and apoptosis in hormone-refractory prostate cancer cell lines by decreasing the activities of PP1 and PP2A. BJU Int. 2012, 110 11 Pt C, E1147–E1154. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.M.; Su, Y.H.; Lin, H.Y.; Lin, J.N.; Liu, L.C.; Ho, C.T.; Way, T.D. Demethoxycurcumin modulates prostate cancer cell proliferation via AMPK-induced down-regulation of HSP70 and EGFR. J. Agric. Food Chem. 2012, 60, 8427–8434. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Garcia-Orti, L.; Cirauqui, C.; Alonso, M.M.; Calasanz, M.J.; Odero, M.D. PP2A impaired activity is a common event in acute myeloid leukemia and its activation by forskolin has a potent anti-leukemic effect. Leukemia 2011, 25, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Manso, R.; Rincón, R.; Caramés, C.; Senin, C.; Borrero, A.; Martínez-Useros, J.; Rodríguez, M.; Zazo, S.; Aguilera, O.; et al. PP2A Inhibition Is a Common Event in Colorectal Cancer and Its Restoration Using FTY720 Shows Promising Therapeutic Potential. Mol. Cancer Ther. 2014, 13, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Rincón, R.; Cristóbal, I.; Zazo, S.; Arpí, O.; Menéndez, S.; Manso, R.; Lluch, A.; Eroles, P.; Rovira, A.; Albanell, J.; et al. PP2A inhibition determines poor outcome and doxorubicin resistance in early breast cancer and its activation shows promising therapeutic effects. Oncotarget 2015, 6, 4299–4314. [Google Scholar] [PubMed]
- Perrotti, D.; Neviani, P. Protein phosphatase 2A: A target for anticancer therapy. Lancet Oncol. 2013, 14, e229–e238. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.S.; Allen-Petersen, B.; Daniel, C.J.; Wang, X.; Wang, Z.; Rodriguez, S.; Impey, S.; Oddo, J.; Vitek, M.P.; Lopez, C.; et al. Targeting Inhibitors of the Tumor Suppressor PP2A for the Treatment of Pancreatic Cancer. Mol. Cancer Res. 2014, 12, 924–939. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, S.; Saliba, C.; Petroni, V.; Fenech, A.G.; Borg, N.; Grech, G. Deregulation of the phosphatase, PP2A is a common event in breast cancer, predicting sensitivity to FTY720. EPMA J. 2014, 5. [Google Scholar] [CrossRef]
- Tonelli, F.; Lim, K.G.; Loveridge, C.; Long, J.; Pitson, S.M.; Tigyi, G.; Bittman, R.; Pyne, S.; Pyne, N.J. FTY720 and (S)-FTY720 vinylphosphonate inhibit sphingosine kinase 1 and promote its proteasomal degradation in human pulmonary artery smooth muscle, breast cancer and androgen-independent prostate cancer cells. Cell Signal. 2010, 22, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, F.; Alossaimi, M.; Williamson, L.; Tate, R.J.; Watson, D.G.; Chan, E.; Bittman, R.; Pyne, N.J.; Pyne, S. The sphingosine kinase inhibitor 2-(p-hyroxyanilino)-4-(p-chlorophenyl)thiazole reduces androgen receptor expression via an oxidative stress-dependent mechanism. Br. J. Pharmacol. 2013, 168, 1497–1505. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristóbal, I.; González-Alonso, P.; Daoud, L.; Solano, E.; Torrejón, B.; Manso, R.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Activation of the Tumor Suppressor PP2A Emerges as a Potential Therapeutic Strategy for Treating Prostate Cancer. Mar. Drugs 2015, 13, 3276-3286. https://doi.org/10.3390/md13063276
Cristóbal I, González-Alonso P, Daoud L, Solano E, Torrejón B, Manso R, Madoz-Gúrpide J, Rojo F, García-Foncillas J. Activation of the Tumor Suppressor PP2A Emerges as a Potential Therapeutic Strategy for Treating Prostate Cancer. Marine Drugs. 2015; 13(6):3276-3286. https://doi.org/10.3390/md13063276
Chicago/Turabian StyleCristóbal, Ion, Paula González-Alonso, Lina Daoud, Esther Solano, Blanca Torrejón, Rebeca Manso, Juan Madoz-Gúrpide, Federico Rojo, and Jesús García-Foncillas. 2015. "Activation of the Tumor Suppressor PP2A Emerges as a Potential Therapeutic Strategy for Treating Prostate Cancer" Marine Drugs 13, no. 6: 3276-3286. https://doi.org/10.3390/md13063276
APA StyleCristóbal, I., González-Alonso, P., Daoud, L., Solano, E., Torrejón, B., Manso, R., Madoz-Gúrpide, J., Rojo, F., & García-Foncillas, J. (2015). Activation of the Tumor Suppressor PP2A Emerges as a Potential Therapeutic Strategy for Treating Prostate Cancer. Marine Drugs, 13(6), 3276-3286. https://doi.org/10.3390/md13063276