
Chitinolytic Bacteria-Assisted Conversion of Squid Pen and Its Effect on Dyes and Pigments Adsorption



Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Identification of a Chitosanase-Producing Strain
2.2. Preparation and Purification of TKU034 Chitosanase


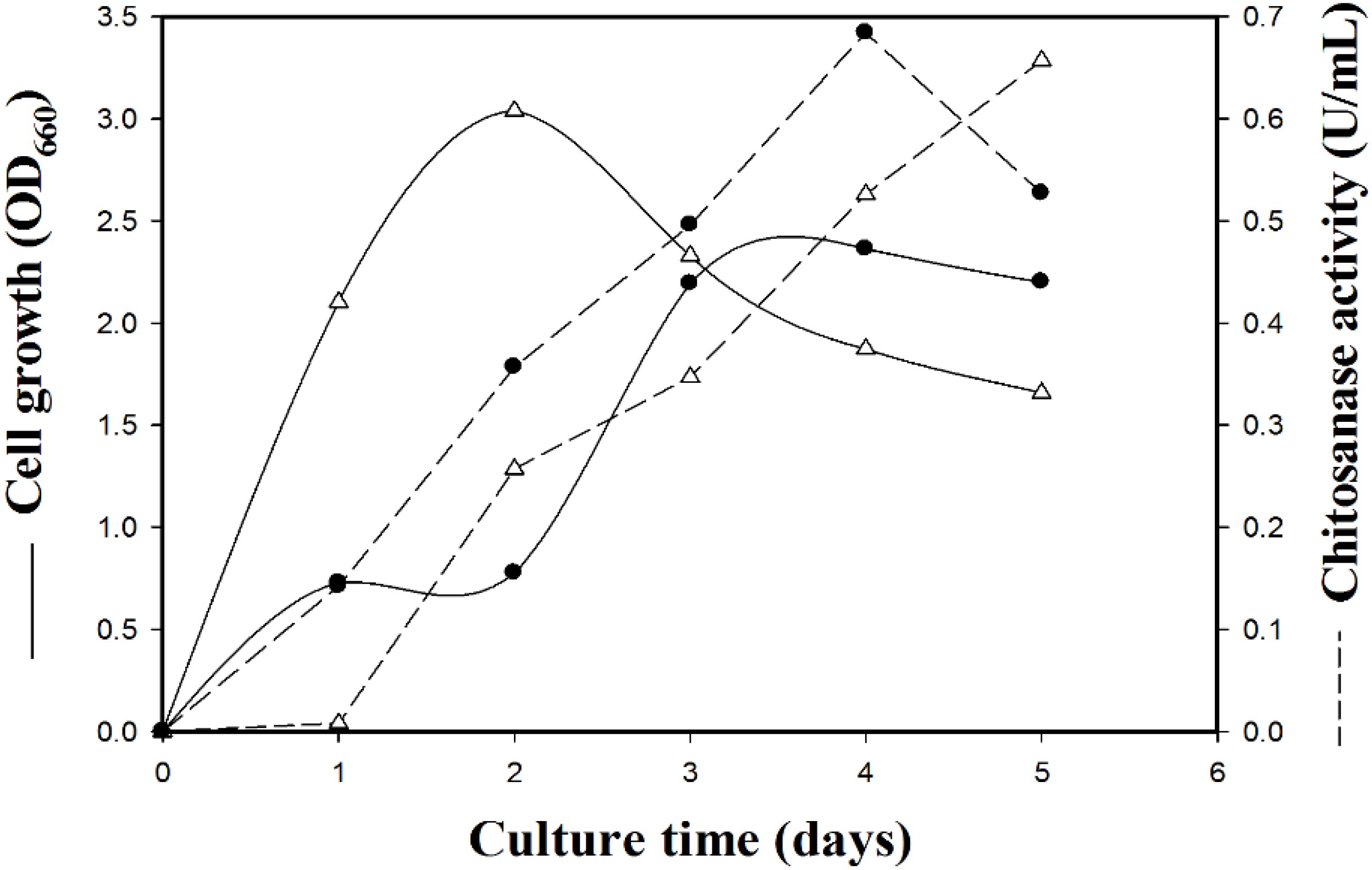{kind=link}
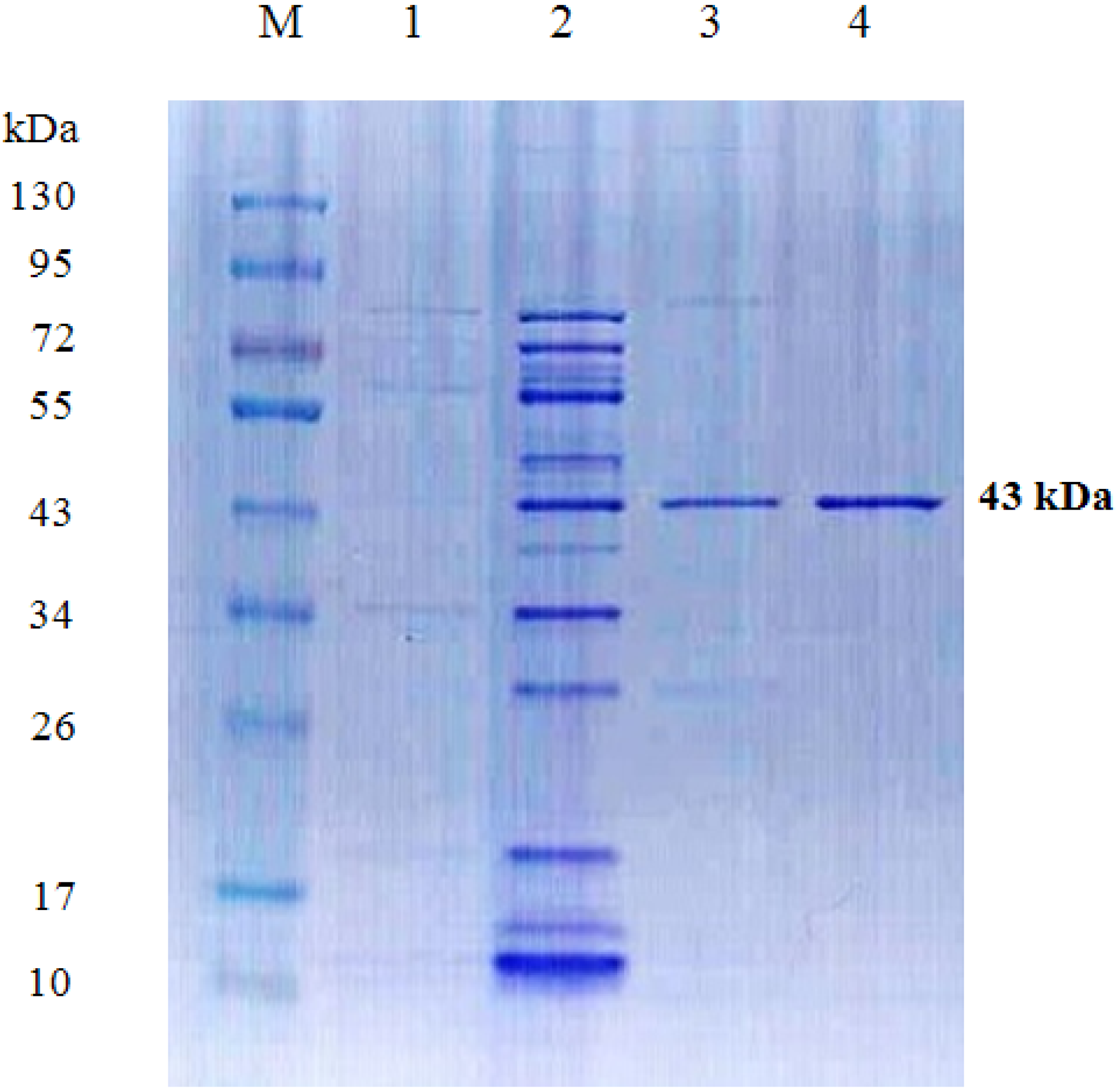{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total | Specific Activity (U/mg) | Purification (fold) | Recovery (%) | ||
|---|---|---|---|---|---|---|
| Volume (mL) | Protein (mg) | Activity (U) | ||||
| Culture supernatant | 730 | 3526 | 445 | 0.13 | 1 | 100 |
| (NH4)2SO4 preciptation | 14.5 | 127.8 | 440 | 3.44 | 26.5 | 98.9 |
| DEAE-sepharose | 38 | 32.87 | 411.3 | 12.51 | 96.2 | 92.4 |
| Marcro-Prep DEAE | 38 | 3.95 | 226.4 | 57.33 | 441.0 | 50.9 |

| Strains | Culture Supernatant | Final Purification | |||||
|---|---|---|---|---|---|---|---|
| Volume (mL) | Protein (mg) | Activity (U) | Activity (U/mL) | Specific Activity (U/mg) | Purification (fold) | Recovery (%) | |
| TKU034 | 730 | 3526 | 445 | 0.610 | 57.33 | 441 | 50.9 |
| TKU033 | 450 | 4032 | 19.4 | 0.043 | 0.052 | 10.4 | 2 |
2.3. Identification of TKU034 Chitosanase by LC-MS/MS Analysis
| Peptide Sequence | Identified Protein and Coverage Rate | Accession Number | |
|---|---|---|---|
| Chitosanase 33% |
|
2.4. Effect of Temperature and pH

2.5. Substrate Specificity
| Substrate | Relative Activity (%) * |
|---|---|
| Water-soluble chitosan | 100 |
| Colloidal chitin from SPP | 0 |
| Suspended chitin α | 1.2 |
| Suspended chitin β | 1.2 |
| Suspended chitin from shrimp shells | 0 |
| Colloidal chitin azure | 0 |
| Suspended chitosan (98% DD, α-type) | 15 |
| Suspended chitosan (98% DD, β-type) | 15 |
| Suspended chitosan (95% DD) | 0 |
| Suspended chitosan (80% DD) | 0 |
| Suspended chitosan (60% DD) | 0 |
| Suspended chitosan from shrimp shells | 7 |
| Suspended chitosan high purity Mw 60,000–120,000 | 0 |
| Suspended chitosan high purity Mw 140,000–220,000 | 8 |
| Suspended chitosan high purity Mw 110,000–150,000 | 34 |
| Soluble CMC | 0 |
2.6. Effects of Surfactants and Metal Ions
| Chemicals | Concentration (mM) | Relative Activity (%) |
|---|---|---|
| None | 0 | 100 |
| Mg2+ | 5 | 91 |
| Fe2+ | 5 | 44 |
| Ca2+ | 5 | 26 |
| Cu2+ | 5 | 0 |
| Ba2+ | 5 | 93 |
| Zn2+ | 5 | 30 |
| Mn2+ | 5 | 22 |
| EDTA | 5 | 59 |
| PMSF | 5 | 111 |
| SDS | 0.5/1/2 | 92/82/84 |
| Tween 20 | 0.5/1/2 (%) | 126/114/112 |
| Tween 40 | 0.5/1/2 (%) | 101/110/109 |
| Triton X-100 | 0.5/1/2 (%) | 115/106/112 |
2.7. Enzymatic Digestion of Chitosan by TKU034 Chitosanase


2.8. Dye Removal by Adsorption Processes Using Fermented SPP

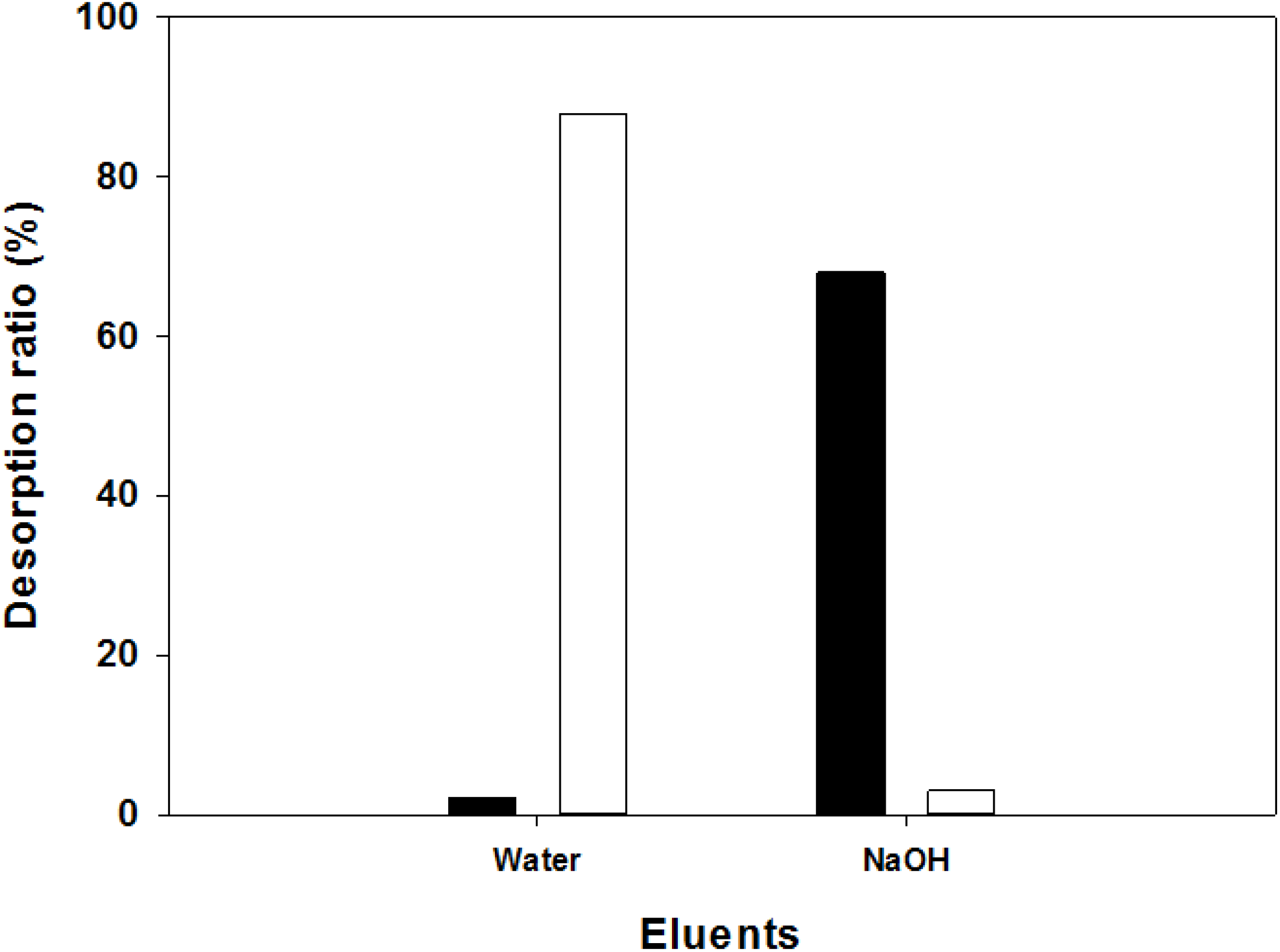
2.9. Adsorption Mechanism

3. Materials and Methods
3.1. Materials
3.2. Screening and Identification of Chitosanase-Producing Strains
3.3. Preparation of Chitosanase
3.4. Chitosanase Activity Assay
3.5. Purification of Chitosanase
3.6. Electrophoresis
3.7. Mass Spectrometry and Protein Identification
3.8. Effects of Temperature and pH on Enzyme Activity and Stability
3.9. Effects of Various Chemicals and Surfactants on Chitosanase Activities
3.10. Adsorption Experiments of Fermented SPP
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Gao, X.A.; Ju, W.T.; Jung, W.J.; Park, R.D. Purification and characterization of chitosanase from Bacillus cereus D-11. Carbohydr. Polym. 2008, 72, 513–520. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, T.R.; Liang, T.W.; Wu, P.C. Conversion and degradation of shellfish wastes by Bacillus cereus TKU018 fermentation for the production of chitosanases and bioactive materials. Biochem. Eng. J. 2009, 48, 111–117. [Google Scholar] [CrossRef]
- Kobayashi, T.; Koide, O.; Deguchi, S.; Horikoshi, K. Characterization of chitosanase of a deep biosphere Bacillus strain. Biosci. Biotechnol. Biochem. 2011, 75, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Pechsrichuang, P.; Yoohat, K.; Yamabhai, M. Production of recombinant Bacillus subtilis chitosanase, suitable for biosynthesis of chitosan-oligosaccharides. Bioresour. Technol. 2013, 127, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Pen, J.H.; Liang, T.W.; Liu, K.C. Purification and characterization of a chitosanase from Serratia marcescens TKU011. Carbohydr. Res. 2008, 343, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, M.G.; Hansen, O.C.; Stougaard, P. Isolation, characterization and heterologous expression of a novel chitosanase from Janthinobacterium sp. strain 4239. Microb. Cell Fact. 2010, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Zitouni, M.; Fortin, M.; Scheerle, R.K.; Letzel, T.; Matteau, D.; Rodrigue, S.; Brzezinski, R. Biochemical and molecular characterization of a thermostable chitosanase produced by the strain Paenibacillus sp. 1794 newly isolated from compost. Appl. Microbiol. Biotechnol. 2013, 97, 5801–5813. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Chen, S.J.; Wang, C.L. Purification and characterization of chitinases and chitosanases from a new species strain Pseudomonas sp.TKU015 using shrimp shells as a substrate. Carbohydr. Res. 2008, 343, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Tseng, W.N.; Liang, T.W. Biodegradation of shellfish wastes and production of chitosanases by a squid pen-assimilating bacterium, Acinetobacter calcoaceticus TKU024. Biodegradation 2011, 22, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Chen, D.; Chen, L.; Yang, G.; Zou, S. Purification, characterization, and action mode of a chitosanase from Streptomyces roseolus induced by chitin. Carbohydr. Res. 2012, 355, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Muzzarelli, R.A.A.; Boudrant, J.; Meyer, D.; Manno, N.; DeMarchis, M.; Paoletti, M.G. Current views on fungal chitin/chitosan, human chitinases, food preservation, glucans, pectins and inulin: A tribute to Henri Braconnot, precursor of the carbohydrate polymers science, on the chitin bicentennial. Carbohydr. Polym. 2012, 87, 995–1012. [Google Scholar] [CrossRef]
- La Thi, K.N.; Wang, S.L.; Dinh, M.H.; Phung, M.L.; Nguyen, T.V.; Tran, M.D.; Nguyen, A.D. Preparation of chitosan nanoparticles by spray dryer and their antibacterial activity. Res. Chem. Intermed. 2014, 40, 2165–2175. [Google Scholar]
- Thadathil, N.; Velappan, S.P. Recent developments in chitosanase research and its biotechnological applications: A review. Food Chem. 2014, 150, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Liu, K.C.; Liang, T.W.; Kuo, Y.H.; Wang, C.Y. In vitro antioxidant activity of liquor and semi-purified fractions from squid pen biowaste by Serratia ureilytica TKU013. Food Chem. 2010, 119, 1380–1385. [Google Scholar] [CrossRef]
- Busilacchi, A.; Gigante, A.; Mattioli-Belmonte, M.; Manzotti, S.; Muzzarelli, R.A.A. Chitosan stabilizes platelet growth factors and modulates stem cell differentiation toward tissue regeneration. Carbohydr. Polym. 2013, 98, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Chen, Y.J.; Yen, Y.H.; Wang, S.L. The antitumor activity of the hydrolysates of chitinous materials hydrolyzed by crude enzyme from Bacillus amyloliquefaciens V656. Process Biochem. 2007, 42, 527–534. [Google Scholar] [CrossRef]
- Wang, S.L.; Lin, T.Y.; Yen, Y.H.; Liao, H.F.; Chen, Y.J. Bioconversion of shellfish chitin wastes for the production of Bacillus subtilis W-118 chitinase. Carbohydr. Res. 2006, 341, 2507–2515. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Lin, H.T.; Liang, T.W.; Chen, Y.J.; Yen, Y.H.; Guo, S.P. Reclamation of chitinous materials by bromelain for the preparation of antitumor and antifungal materials. Bioresour. Technol. 2008, 99, 4386–4393. [Google Scholar] [CrossRef] [PubMed]
- Goo, B.G.; Park, J.K. Characterization of an alkalophilic extracellular chitosanase from Bacillus cereus GU-02. J. Biosci. Bioeng. 2014, 117, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Azlan, K.; Wan Saime, W.N.; Lai Ken, L. Chitosan and chemically modified chitosan beads for acid dyes sorption. J. Environ. Sci. 2009, 21, 296–302. [Google Scholar] [CrossRef]
- Wang, C.L.; Chen, C.J.; Nguyen, A.D.; Liang, T.W.; Twu, Y.K.; Huang, S.Y.; Wang, S.L. Environmental chitinous materials as adsorbents for the one-step purification of protease and chitosanase. Res. Chem. Intermed. 2014, 40, 2363–2370. [Google Scholar] [CrossRef]
- Dotto, G.L.; Pinto, L.A.A. Adsorption of food dyes onto chitosan: Optimization process and kinetic. Carbohydr. Polym. 2011, 84, 231–238. [Google Scholar] [CrossRef]
- Wan Ngah, W.S.; Ariff, N.F.M.; Hanafiah, M.A.K.M. Preparation, characterization, and environmental application of crosslinked chitosan-coated bentonite for tartrazine adsorption from aqueous solutions. Water Air Soil Pollut. 2010, 206, 225–236. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, S.Y.; Yen, Y.H.; Liang, T.W. Utilization of chitinous materials in pigment adsorption. Food Chem. 2012, 135, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.J.; Jo, G.H.; Kuk, J.H.; Kim, Y.J.; Oh, K.T.; Park, R.D. Production of chitin from red crab shell waste by successive fermentation with Lactobacillus paracasei KCTC-3037 and Serratia marcescens FS-3. Carbohydr. Polym. 2007, 68, 746–750. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, Y.Y.; Pan, P.S.; Wang, S.L. Purification of chitinase/chitosanase from Bacillus cereus and discovery of an enzyme inhibitor. Int. J. Biol. Macromol. 2014, 63, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Hsieh, J.L.; Wang, S.L. Production and purification of a protease, a chitosanase, and chitinoligosaccharides by Bacillus cereus TKU022 fermentation. Carbohydr. Res. 2012, 362, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Su, J.W.; Liang, T.W.; Nguyen, A.D.; Wang, S.L. Production, purification and characterization of a chitosanase from Bacillus cereus. Res. Chem. Intermed. 2014, 40, 2237–2248. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, E.J.; Piao, Z.; Yun, Y.C.; Shin, Y.C. Purification and characterization of chitosanase from Bacillus sp. strain KCTC 0377BP and its application for the production of chitosan oligosaccharides. Appl. Environ. Microbiol. 2004, 70, 4522–4531. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Wang, D.; Yao, L.; Yu, Z. Purification, characterization, and gene cloning of a chitosanase from Bacillus species strain S65. J. Agric. Food Chem. 2006, 54, 4208–4214. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Huang, C.T.; Nguyen, A.D.; Wang, S.L. Squid pen chitin chitooligomers as food colorants absorbers. Mar. Drugs 2015, 13, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Katsumi, T.; Lacombe-Harvey, M.; Tremblay, H.; Brzezinski, R.; Fukamizo, T. Role of acidic amino acid residues in chitooligosaccharide-binding to Streptomyces sp. N174 chitosanase. Biochem. Biophys. Res. Commun. 2005, 338, 1839–1844. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, W.; Yuan, H.; Wang, Y. Characterization of a novel fungal chitosanase Csn2 from Gongronella sp. JG. Carbohydr. Res. 2008, 343, 2583–2588. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.-W.; Lo, B.-C.; Wang, S.-L. Chitinolytic Bacteria-Assisted Conversion of Squid Pen and Its Effect on Dyes and Pigments Adsorption. Mar. Drugs 2015, 13, 4576-4593. https://doi.org/10.3390/md13084576
Liang T-W, Lo B-C, Wang S-L. Chitinolytic Bacteria-Assisted Conversion of Squid Pen and Its Effect on Dyes and Pigments Adsorption. Marine Drugs. 2015; 13(8):4576-4593. https://doi.org/10.3390/md13084576
Chicago/Turabian StyleLiang, Tzu-Wen, Bo-Chang Lo, and San-Lang Wang. 2015. "Chitinolytic Bacteria-Assisted Conversion of Squid Pen and Its Effect on Dyes and Pigments Adsorption" Marine Drugs 13, no. 8: 4576-4593. https://doi.org/10.3390/md13084576
APA StyleLiang, T.-W., Lo, B.-C., & Wang, S.-L. (2015). Chitinolytic Bacteria-Assisted Conversion of Squid Pen and Its Effect on Dyes and Pigments Adsorption. Marine Drugs, 13(8), 4576-4593. https://doi.org/10.3390/md13084576