
Mechanistic Insight into the Elastin Degradation Process by the Metalloprotease Myroilysin from the Deep-Sea Bacterium Myroides profundi D25

 ,
, 

Abstract
:1. Introduction
2. Results
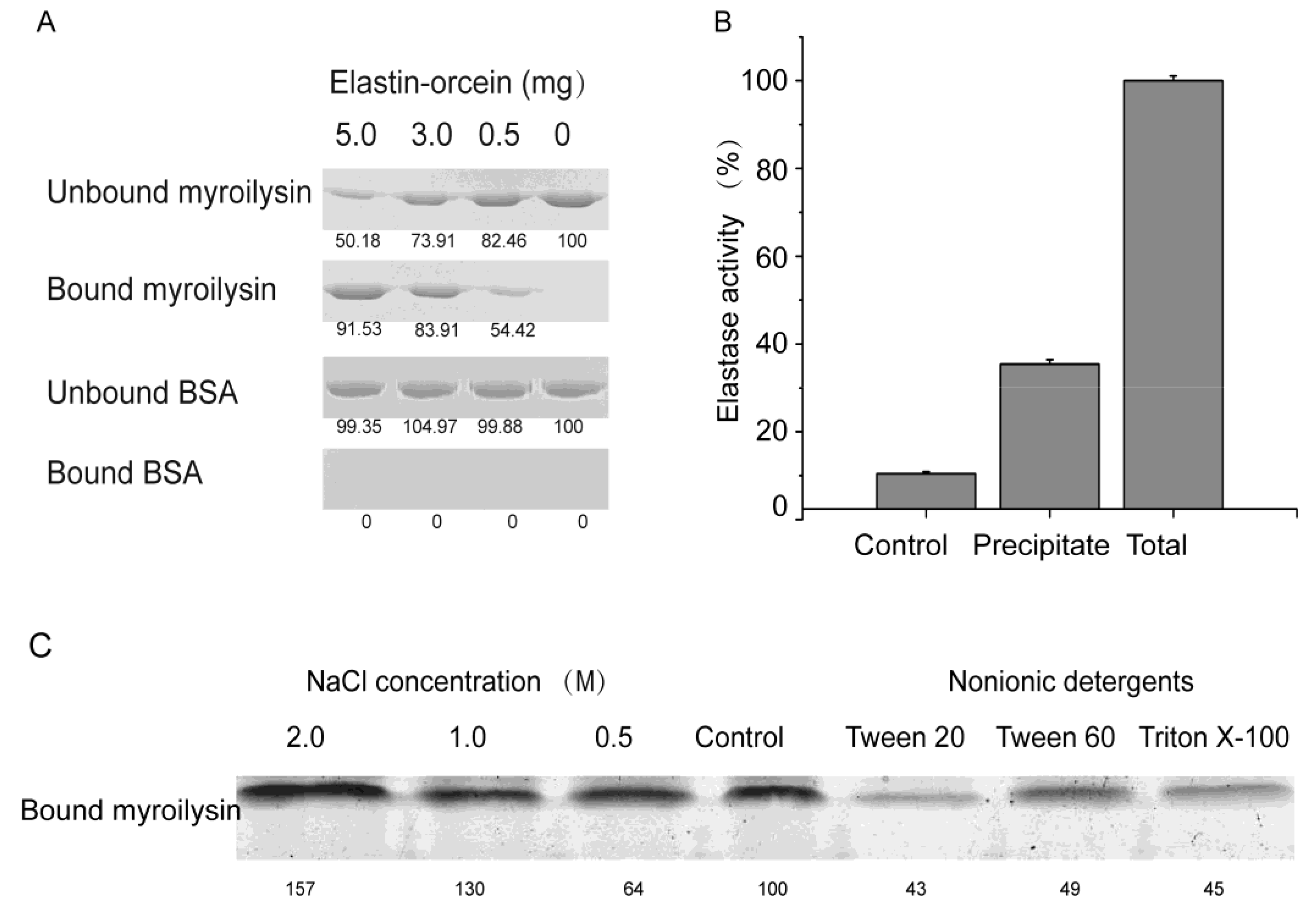
2.1. Hydrophobic Binding of Myroilysin to Insoluble Elastin
2.2. Cleavage Sites of Myroilysin in Bovine Elastin



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P4 | P3 | P2 | P1 | P1′ | P2′ | P3′ | P4′ | Occurrence Number | |
|---|---|---|---|---|---|---|---|---|---|
| G | 30.86 | 37.04 | 23.46 | 64.20 | 25.93 | 32.10 | 46.91 | 22.22 | 229 |
| V | 17.28 | 20.99 | 23.46 | 3.70 | 43.21 | 17.28 | 19.75 | 27.16 | 140 |
| P | 20.99 | 13.58 | 17.28 | 4.94 | 1.23 | 20.99 | 12.35 | 22.22 | 92 |
| A | 14.81 | 11.11 | 13.58 | 14.81 | 9.88 | 7.41 | 4.94 | 9.88 | 70 |
| L | 4.94 | 3.70 | 8.64 | 4.94 | 7.41 | 9.88 | 6.17 | 8.64 | 44 |
| Y | 1.23 | 1.23 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2 |
| F | 1.23 | 1.23 | 2.47 | 2.47 | 3.70 | 4.94 | 3.70 | 2.47 | 18 |
| I | 2.47 | 1.23 | 2.47 | 0.00 | 3.70 | 2.47 | 0.00 | 2.47 | 12 |
| K | 3.70 | 3.70 | 1.23 | 0.00 | 0.00 | 0.00 | 1.23 | 1.23 | 9 |
| Q | 0.00 | 2.47 | 3.70 | 1.23 | 2.47 | 1.23 | 0.00 | 1.23 | 10 |
| T | 0.00 | 2.47 | 2.47 | 1.23 | 1.23 | 1.23 | 1.23 | 2.47 | 10 |
| R | 1.23 | 0.00 | 0.00 | 1.23 | 0.00 | 0.00 | 1.23 | 0.00 | 3 |
| S | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 | 2.47 | 2.47 | 0.00 | 9 |

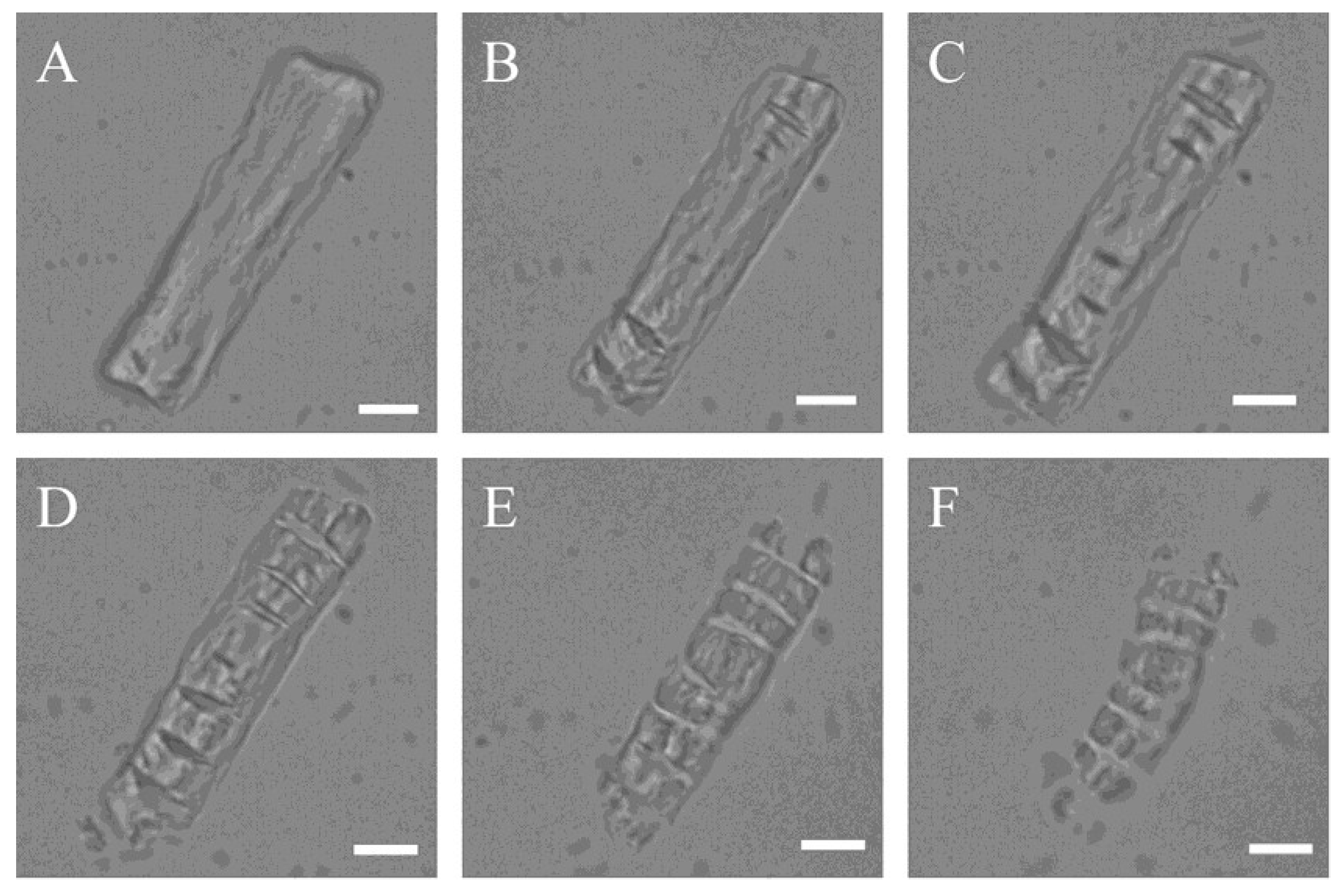
2.3. LM and SEM Observations of Bovine Elastin Fibre Degradation by Myroilysin

2.4. SEM Observation of the Degradation of Cross-Linked Recombinant Tropoelastin Spherules by Myroilysin

2.5. Cleavage Sites of Myroilysin in Recombinant Tropoelastin

| P4 | P3 | P2 | P1 | P1′ | P2′ | P3′ | P4′ | Occurrence Number | |
|---|---|---|---|---|---|---|---|---|---|
| G | 28.89 | 22.22 | 35.56 | 36.67 | 36.67 | 16.67 | 31.11 | 22.22 | 207 |
| V | 14.44 | 13.33 | 3.33 | 7.78 | 12.22 | 16.67 | 8.89 | 22.22 | 89 |
| P | 14.44 | 31.11 | 8.89 | 11.11 | 3.33 | 27.78 | 15.56 | 20.00 | 119 |
| A | 20.00 | 14.44 | 17.78 | 21.11 | 22.22 | 17.78 | 17.78 | 14.44 | 131 |
| L | 5.56 | 6.67 | 10.00 | 6.67 | 11.11 | 8.89 | 6.67 | 6.67 | 56 |
| Y | 3.33 | 0.00 | 7.78 | 2.22 | 1.11 | 1.11 | 3.33 | 0.00 | 17 |
| F | 3.33 | 2.22 | 5.56 | 4.44 | 1.11 | 0.00 | 1.11 | 1.11 | 17 |
| I | 1.11 | 1.11 | 3.33 | 0.00 | 5.56 | 3.33 | 1.11 | 2.22 | 16 |
| C | 0.00 | 0.00 | 0.00 | 0.00 | 1.11 | 1.11 | 1.11 | 1.11 | 4 |
| K | 3.33 | 5.56 | 3.33 | 7.78 | 3.33 | 3.33 | 7.78 | 5.56 | 36 |
| Q | 1.11 | 1.11 | 2.22 | 0.00 | 0.00 | 0.00 | 1.11 | 1.11 | 6 |
| T | 1.11 | 1.11 | 0.00 | 1.11 | 2.22 | 0.00 | 0.00 | 1.11 | 6 |
| R | 1.11 | 0.00 | 0.00 | 1.11 | 0.00 | 1.11 | 2.22 | 1.11 | 6 |
| S | 2.22 | 0.00 | 1.11 | 0.00 | 0.00 | 2.22 | 2.22 | 1.11 | 8 |
| E | 0.00 | 1.11 | 1.11 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2 |
3. Discussion
4. Experimental Section
4.1. Reagents and Materials
4.2. Assay of the Ability of Myroilysin to Bind to Insoluble Elastin
4.3. Effects of NaCl and Nonionic Detergents on the Binding of Myroilysin to Insoluble Elastin
4.4. LM In Situ Observation of Bovine Elastin Fibre Degradation by Myroilysin
4.5. SEM Observation of Bovine Elastin Fibre Degradation by Myroilysin
4.6. SEM Observation of the Degradation of Cross-Linked Recombinant Tropoelastin by Myroilysin
4.7. Analysis of Myroilysin Cleavage Sites in Bovine Elastin and Recombinant Human Tropoelastin
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rosenbloom, J.; Abrams, W.R.; Mecham, R. Extracellular matrix 4: The elastic fiber. FEBS J. 1993, 7, 1208–1218. [Google Scholar]
- Wise, S.G.; Weiss, A.S. Tropoelastin. Int. J. Biochem. Cell Biol. 2009, 41, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Xie, B.B.; Bian, F.; Zhao, G.Y.; Zhao, H.L.; He, H.L.; Zhou, B.C.; Zhang, Y.Z. Ecological function of myroilysin, a novel bacterial M12 metalloprotease with elastinolytic activity and a synergisticrole in collagen hydrolysis, in biodegradation of deep-sea high-molecular-weight organic nitrogen. Appl. Environ. Microbiol. 2009, 75, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.M.; Indik, Z.; Yeh, H.; Ornstein-Goldstein, N.; Rosenbloom, J.C.; Abrams, W.; Fazio, M.; Uitto, J.; Rosenbloom, J. Characterization of the complete human elastin gene. Delineation of unusual features in the 5′-flanking region. J. Biol. Chem. 1989, 264, 8887–8891. [Google Scholar] [PubMed]
- Indik, Z.; Yeh, H.; Ornstein-Goldstein, N.; Sheppard, P.; Anderson, N.; Rosenbloom, J.C.; Peltonen, L.; Rosenbloom, J. Alternative splicing of human elastin mRNA indicated by sequence analysis of cloned genomic and complementary DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 5680–5684. [Google Scholar] [CrossRef] [PubMed]
- Indik, Z.; Yeh, H.; Ornstein-Goldstein, N.; Kucich, U.; Abrams, W.; Rosenbloom, J.C.; Rosenbloom, J. Structure of the elastin gene and alternative splicing of elastin mRNA: Implications for human disease. Am. J. Med. Genet. 1989, 34, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.C.; Keeley, F.W.; Weiss, A.S. Coacervation of tropoelastin. Adv. Colloid Interface Sci. 2011, 167, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Mithieux, S.M.; Weiss, A.S. Elastin. Adv. Protein Chem. 2005, 70, 437–461. [Google Scholar] [PubMed]
- Vrhovski, B.; Weiss, A.S. Biochemistry of tropoelastin. Eur. J. Biochem. 1998, 258, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Jenkins, N.P.; Huang, W.C.; di Gregoli, K.; Sala-Newby, G.B.; Scholtes, V.P.; Moll, F.L.; Pasterkamp, G.; Newby, A.C. Relationship of MMP-14 and TIMP-3 expression with macrophage activation and human atherosclerotic plaque vulnerability. Mediat. Inflamm. 2014, 276457. [Google Scholar] [CrossRef]
- Johnson, J.L.; Devel, L.; Czarny, B.; George, S.J.; Jackson, C.L.; Rogakos, V.; Beau, F.; Yiotakis, A.; Newby, A.C.; Dive, V. A selective matrix metalloproteinase-12 inhibitor retards atherosclerotic plaque development in apolipoprotein E-knockout mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.A.; Rocco, P.R. Elastase-induced pulmonary emphysema: Insights from experimental models. An. Acad. Bras. Cienc. 2011, 83, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; He, G.; Jiao, Y.; Ni, H. Effects of elastase from a Bacillus strain on the tenderization of beef meat. Food Chem. 2006, 98, 624–629. [Google Scholar] [CrossRef]
- Zhao, H.L.; Chen, X.L.; Xie, B.B.; Zhou, M.Y.; Gao, X.; Zhang, X.Y.; Zhou, B.C.; Weiss, A.S.; Zhang, Y.Z. Elastolytic mechanism of a novel M23 metalloprotease pseudoalterin from deep-sea Pseudoalteromonas sp. CF6–2: Cleaving not only glycyl bonds in the hydrophobic regions but also peptide bonds in the hydrophilic regions involved in cross-linking. J. Biol. Chem. 2012, 287, 39710–39720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Zhang, Y.J.; Chen, X.L.; Qin, Q.L.; Zhao, D.L.; Li, T.G.; Dang, H.Y.; Zhang, Y.Z. Myroides profundi sp. nov., isolated from deep-sea sediment of the southern Okinawa Trough. FEMS Microbiol. Lett. 2008, 287, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Fujita, J.; Sköld, C.M.; Daughton, D.M.; Ertl, R.F.; Takahara, J.; Rennard, S.I. Modulation of elastase binding to elastin by human alveolar macrophage-derived lipids. Am. J. Respir. Crit. Care Med. 1999, 160, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K. Collagenolytic proteases from bacteria. Appl. Microbiol. Biotechnol. 2004, 63, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Y.; Chen, X.L.; Zhao, H.L.; Xie, B.B.; Zhou, B.C.; Zhang, Y.Z. Hydrolysis of insoluble collagen by deseasin MCP-01 from deep-sea Pseudoalteromonas sp. SM9913: Collagenolytic characters, collagen-binding ability of novel role in deep-sea sedimentary particulate organic nitrogen degradation. J. Biol. Chem. 2008, 283, 36100–36107. [Google Scholar] [CrossRef] [PubMed]
- Brown-Augsburger, P.; Tisdale, C.; Broekelmann, T.; Sloan, C.; Mecham, R.P. Identification of an elastin cross-linking domain that joins three peptide chains: Possible role in nucleated assembly. J. Biol. Chem. 1995, 270, 17778–17783. [Google Scholar] [CrossRef] [PubMed]
- Baldock, C.; Oberhauser, A.F.; Ma, L.; Lammie, D.; Siegler, V.; Mithieux, S.M.; Tu, Y.; Chow, J.Y.; Suleman, F.; Malfois, M.; et al. Shape of tropoelastin, the highly extensible protein that controls human tissue elasticity. Proc. Natl. Acad. Sci. USA 2011, 108, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.C.; Baldock, C.; Tuukkanen, A.; Roessle, M.; Dyksterhuis, L.B.; Wise, S.G.; Matthews, J.; Mithieux, S.M.; Weiss, A.S. Tropoelastin bridge region positions the cell-interactive C terminus and contributes to elastic fiber assembly. Proc. Natl. Acad. Sci. USA 2012, 109, 2878–2883. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.; Jung, M.C.; Duca, L.; Sippl, W.; Taddese, S.; Ihling, C.; Rusciani, A.; Jahreis, G.; Weiss, A.S.; Reinhard, H.H.; et al. Schmelzer1 Degradation of tropoelastin by matrix metalloproteinases—Cleavage site specificities and release of matrikines. FEBS J. 2010, 277, 1939–1956. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Visse, R.; Murohy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.L.; Vrhovski, B.; Weiss, A.S. Total synthesis andexpression in Escherichia coli of a gene encoding human tropoelastin. Gene 1995, 154, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Vrhovski, B.; Weiss, A.S. Glycosaminoglycans mediate the coacervation of human tropoelastin through dominant charge interactions involving lysine side chains. J. Biol. Chem. 1999, 274, 21719–21724. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, S.V.; Diaz, P.; Pastor, F.I. Modular glucuronoxylan-specific xylanase with a family CBM35 carbohydrate-binding module. Appl. Environ. Microbiol. 2012, 78, 3923–3931. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Zhao, H.-L.; Tang, B.-L.; Chen, X.-L.; Su, H.-N.; Zhang, X.-Y.; Song, X.-Y.; Zhou, B.-C.; Xie, B.-B.; Weiss, A.S.; et al. Mechanistic Insight into the Elastin Degradation Process by the Metalloprotease Myroilysin from the Deep-Sea Bacterium Myroides profundi D25. Mar. Drugs 2015, 13, 1481-1496. https://doi.org/10.3390/md13031481
Yang J, Zhao H-L, Tang B-L, Chen X-L, Su H-N, Zhang X-Y, Song X-Y, Zhou B-C, Xie B-B, Weiss AS, et al. Mechanistic Insight into the Elastin Degradation Process by the Metalloprotease Myroilysin from the Deep-Sea Bacterium Myroides profundi D25. Marine Drugs. 2015; 13(3):1481-1496. https://doi.org/10.3390/md13031481
Chicago/Turabian StyleYang, Jie, Hui-Lin Zhao, Bai-Lu Tang, Xiu-Lan Chen, Hai-Nan Su, Xi-Ying Zhang, Xiao-Yan Song, Bai-Cheng Zhou, Bin-Bin Xie, Anthony S. Weiss, and et al. 2015. "Mechanistic Insight into the Elastin Degradation Process by the Metalloprotease Myroilysin from the Deep-Sea Bacterium Myroides profundi D25" Marine Drugs 13, no. 3: 1481-1496. https://doi.org/10.3390/md13031481
APA StyleYang, J., Zhao, H. -L., Tang, B. -L., Chen, X. -L., Su, H. -N., Zhang, X. -Y., Song, X. -Y., Zhou, B. -C., Xie, B. -B., Weiss, A. S., & Zhang, Y. -Z. (2015). Mechanistic Insight into the Elastin Degradation Process by the Metalloprotease Myroilysin from the Deep-Sea Bacterium Myroides profundi D25. Marine Drugs, 13(3), 1481-1496. https://doi.org/10.3390/md13031481