1. Introduction
Routinely performed sperm tests, such as semen volume, colour, pH, liquefaction time, viscosity, sperm count and motility, sperm morphology, concentration of round cells and polymorphonucleocytes, sperm agglutination and sperm viability, often do not explain male inability to achieve a pregnancy after 12 months or more of regular, unprotected and well-timed intercourse. The term idiopathic infertility is due to the fact that results of these tests typically fall within the normal range without clarifying the exact reason why patients result infertile/subfertile [
1].
In order to acquire the correct features for potential oocyte fertilization, human sperm has to undergo a series of transformations, known as capacitation, which prepares the cell to undergo the acrosome reaction (AR). During the capacitation process, the sperm outer membrane undergoes cholesterol depletion [
2,
3], to augment membrane fusibility and enhance the acrosomal exocytosis, followed by downstream signaling processes such as protein serine and tyrosine phosphorylation (Tyr-P) [
4,
5,
6] and reactive oxygen species (ROS) generation [
7].
Capacitation-related efflux of cholesterol, a major component of lipid rafts [
8,
9], might also change membrane fluidity and/or induce rearrangement of membrane lipid rafts [
10,
11].
Generally, lipid rafts are small microdomains ranging from 10 to 200 nm in size characterized by the ganglioside GM1, a raft constituent and marker, which, by binding to the fluorescence-conjugated cholera toxin B subunit (CTB) [
12], may be identified and monitored during capacitation transformations [
13].
Caveolin 1 (CAV 1) is one of rafts’ constituents [
14] and provides the scaffolding of these microdomains, also called caveolae [
15], which can embed and inactivate many proteins and enzymes [
14].
High ROS level and oxidative stress have been implicated in the pathophysiology of male infertility [
16] being correlated with sperm DNA damage, reduced sperm motility, and lipid and protein denaturation [
16,
17]. However, growing evidence has been recently proposed [
18] about the essential role of a controlled ROS generation on the correct sperm functioning, thus giving ROS a key role in the maturation process, other than the detrimental factor previously assessed [
19].
Moreover, the time-dependent ROS generation has been demonstrated to represent an important tool to predict the potential ability of the cell to accomplish acrosome reaction [
13,
18,
19,
20], discriminating sperms that, either for lower or higher ROS content, were prevented from being capacitated [
18]. In addition, the close relationship between the Tyr-P of the sperm head and AR has been demonstrated [
18,
19], although the molecular mechanisms involved in all these capacitation-related processes remain to be characterized.
Phosphorylation is among the most common regulatory mechanisms for protein function, regulating cell functions by inducing conformational changes in proteins via allosteric modification [
21]. In human sperm, several members of the Src kinase family have recently been described [
22,
23,
24], but, despite the importance of the events triggered by Tyr-P such as hyperactivation [
24], capacitation progress and acrosome reaction regulation [
25], protein tyrosine kinases (PTKs) identification remains to be completely elucidated. At present, the most well characterized mechanisms of Tyr-P are regulated by members of the Src family kinases (SFK), which is the largest family of non-receptor tyrosine kinases widely expressed in many cell types and in different subcellular compartments [
26]. SFKs induce cellular responses associated with proliferation, growth control, survival, differentiation and cytoskeletal arrangements [
26] and many of their own functions have been clarified with the use of inhibitors of their activities, such as 4-Amino-5-(4-chlorophenyl)-7-(
t-butyl)pyrazolo[3,4-d]pyrimidine (PP2) [
27].
While the role of hormone therapy for men with an identified abnormality is well defined [
28], the literature remains inconclusive and controversial regarding the use of food supplements for men with idiopathic infertility, mainly due to the incomplete knowledge of all the steps of the capacitation process.
Many medical therapies have been historically used for male infertility, including herbs, vitamins, and nutritional supplements, and many of them rely on antioxidant properties. Astaxanthin (Asta), a carotenoid nutrient widely distributed in algae, crustaceans, shellfish and various plants [
29], inserts within cell membranes protecting them with potent antioxidant and anti-inflammatory actions [
30,
31]. In a recent study [
20], we described that Asta ameliorates viability and AR percentages of human sperm. To test the hypothesis of whether Asta could induce protein-protein or protein-lipid dislocation at the plasma membrane level, in the present study we evaluated the effect of Asta on sperm membrane raft reorganization, in particular at the level of head/acrosome region. In addition, the presence of Lyn, a Src family kinase found in lipid rafts [
32,
33], was investigated as potential Tyr-kinase involved in the capacitation-related Tyr-P.
2. Results and Discussion
We have previously described that Asta can improve human sperm viability in a dose-dependent manner with 2 μM dose representing optimal concentration for capacitation [
20].
After gradient separation, sperm samples of each group were collected three by three in a single pool to obtain a sufficient number of cells to perform all tests. Pools were divided in samples which were immediately analysed (T
0), or incubated in the absence (C), or presence of Asta, PP2, an inhibitor of the Src family kinase, or both (Asta + PP2). Before any further analysis, aliquots were then assessed for sperm parameters by computer-assisted sperm analysis (CASA) (
Supplementary Table S1). As expected, samples C showed a net increase in the different motility parameters compared with T
0, thus confirming that capacitation-related hyper-activation was occurred. No significant alteration was evident by comparing capacitated sperm (C) with those with Asta, or Asta + PP2, except for a minor increase in patient group (PG) sperm treated with Asta, thus suggesting that hyperactivity was almost not affected by these different conditions.
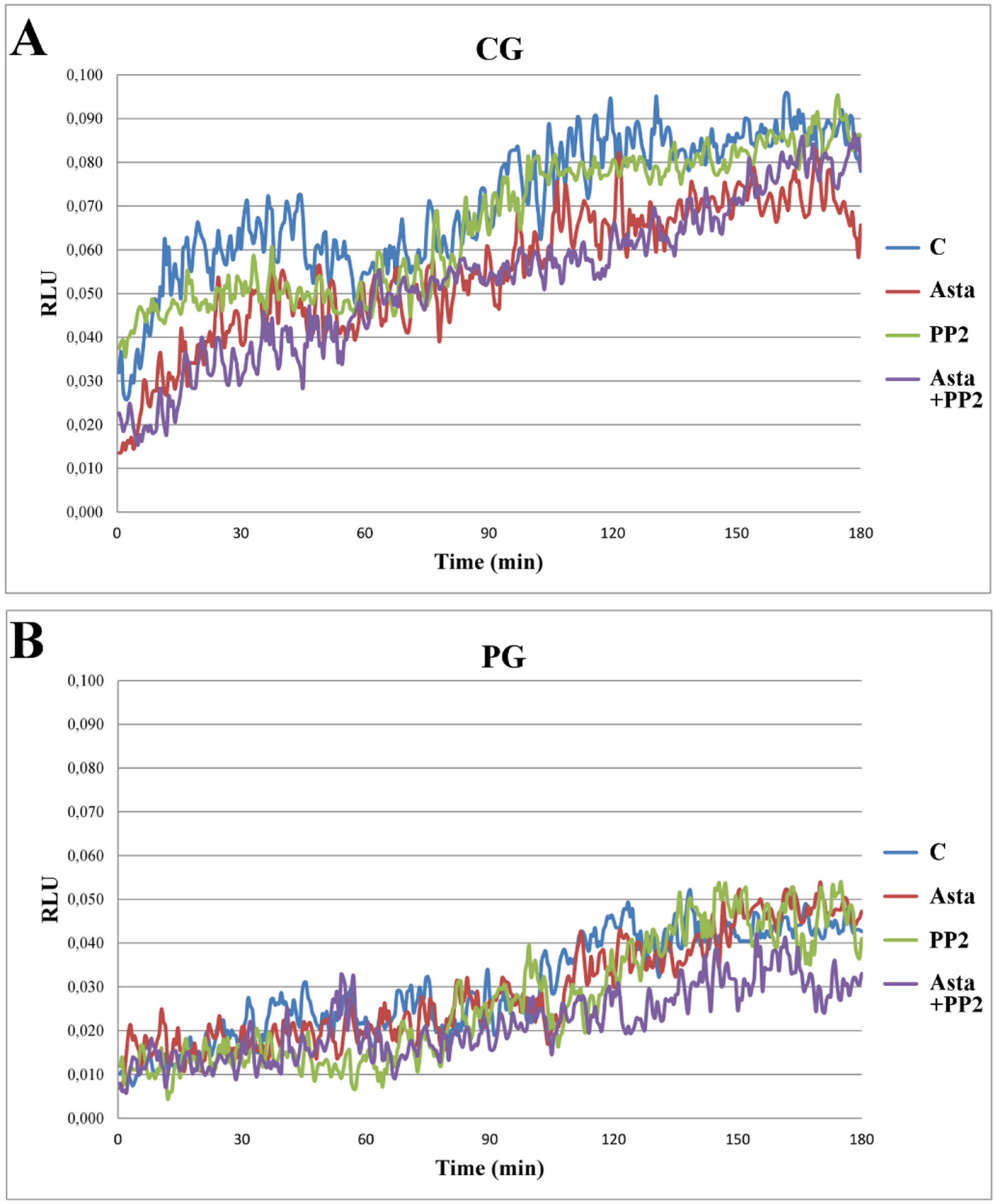
Another aliquot of each sample was analysed for the ROS generation in a luminometer for 180 min with luminol as a luminescent source. Samples C generating ROS in a concentration sufficient to induced luminescence between the values 0.05–0.08 Relative Luminescence Units (RLU) were considered normal [
18] and, therefore, belonging to the control group (CG,
n = 15), whereas samples expressing ROS below these values were inserted in the patient group (PG,
n = 18). On the basis of this classification, samples which did not meet the above criteria were discarded.
When analysed in the presence of Asta, no relevant alteration in the curves of ROS generation (
Figure 1) was observed in either CG (panel A) or PG (panel B) groups, although evaluating the integrated values (as area under the curve, AUC) a modest, but significant (
p < 0.01), reduction was found in RLU of CG group, also in the presence of both Asta + PP2 (panel C). This fact suggests that Asta has a modest role in preventing H
2O
2 formation in sperm in physiological condition but no effect in patients.
Figure 1.
ROS generation curves of sperm samples and effects of Asta, PP2, or both. Sperm from three volunteers for each experiment was collected to form a pool with a sufficient number of cells. Sperm was incubated for up to 180 min in capacitating conditions in the absence (C) or presence of Asta (2 μM), PP2 (6 μM), or both (Asta + PP2, 2 μM + 6 μM). Luminol chemiluminescence was monitored during sperm capacitation. Results are expressed as moving averages of Relative Luminescence Units (RLU)/30 s for 2 × 10
6 cells.
Figure 1 is representative of 11 separate experiments, conducted on samples belonging to the control group (CG, panel
A) or the patient group (PG, panel
B). Detection was carried out in triplicate. Graphs in panel
C show AUC of the RLU data in both groups. **
p < 0.01
vs. C; Student’s
t test.
Figure 1.
ROS generation curves of sperm samples and effects of Asta, PP2, or both. Sperm from three volunteers for each experiment was collected to form a pool with a sufficient number of cells. Sperm was incubated for up to 180 min in capacitating conditions in the absence (C) or presence of Asta (2 μM), PP2 (6 μM), or both (Asta + PP2, 2 μM + 6 μM). Luminol chemiluminescence was monitored during sperm capacitation. Results are expressed as moving averages of Relative Luminescence Units (RLU)/30 s for 2 × 10
6 cells.
Figure 1 is representative of 11 separate experiments, conducted on samples belonging to the control group (CG, panel
A) or the patient group (PG, panel
B). Detection was carried out in triplicate. Graphs in panel
C show AUC of the RLU data in both groups. **
p < 0.01
vs. C; Student’s
t test.
The close relation between the endogenous ROS content (
Supplementary Figure S1) and sperm viability (
Supplementary Table S2) were further investigated by incubating CG sperm in the presence of ascorbic acid (AA), and PG sperm with H
2O
2 (0.25 μM). As previously reported [
18], AA greatly reduced ROS content (panel A) and prevented capacitation as shown by Tyr-P and AR (
Supplementary Table S2). On the other hand, when PG sperm was treated in the presence of H
2O
2 to ameliorate ROS content (panel B) and trigger capacitation, although a peak within the region of correct values of ROS was formed, no improvement either in Tyr-P level and distribution or acrosome reacted cells (ARC) was shown (
Supplementary Table S2). In this case, it is hypothesized that the impairment, present in PG sperm, was not accessible by exogenous ROS addition, probably due to its inner insertion within the membrane. Higher ROS doses were not consistent with cell survival [
18] as suggested by the increase of non-viable cells (NVC) percentage (
Supplementary Table S2).
At the end of incubation, aliquots of cells were withdrawn from all samples and analyzed for their ability to undergo Tyr-P and AR.
The Tyr-P pattern was evaluated (
Supplementary Figure S2 and Table S1). According to previous reports [
19] (
Supplementary Figure S2, panel A), capacitation induced a net increase in the total cell lysate Tyr-P level of two main bands at about 90 and 110 kDa, respectively, in the CG, with Asta showing only a slight further enhancement and PP2 completely preventing any increase either alone or in co-addition with Asta (lanes PP2 and Asta + PP2, respectively). On the other hand, in the PG, capacitation-related increase of the Tyr-P was clearly lower (only 20% compared with T
0,
p < 0.001) with Asta inducing a net increase (40% compared with T
0,
p < 0.001). PP2 completely inhibited Tyr-P increase in any conditions (lanes PP2 and Asta + PP2).
When cells were analysed for Tyr-P distribution with immunocytochemistry (
Supplementary Figure S2), as previously reported [
19], in sample T
0, Tyr-P was mainly located in the mid-piece region and tails of spermatozoa [
19] and involved about 95% of the cells (
Table 1, Tyr-P cells) in CG but only 48% of cells in PG. Cells presenting Tyr-P also in the region of head (Tyr-P head) were about 10% or less in both CG and PG. After 180 min in capacitating conditions (C), only a few cells of PG presented Tyr-P in the head region (19% ± 3%), as expected on the basis of ROS generation values expressed by cells (
Figure 1B), compared to C sample of CG (63% ± 6%). Interestingly, the addition of Asta clearly increased the percentage of PG sperm presenting Tyr-P in head (27% ± 3% compared to 19% ± 3% in C,
p < 0.0001) although ROS generation content was not affected at all (
Figure 1B). On the contrary, in CG samples, Asta did not induce any significant increase compared with C condition. When added, PP2, either alone or in co-addition with Asta, drastically diminished the percentage of cells presenting Tyr-P in both groups (
Table 1), thus confirming the involvement of Src family kinase in the capacitation-related Tyr-P process.
Table 1.
Sperm cells from control group (CG) or patient group (PG) at T
0, or incubated for 180 min in capacitating conditions in the absence (C) or presence of Asta 2 μM, PP2 6 μM or both (Asta + PP2), were analyzed for Tyr-P pattern (
Supplementary Figure S2), acrosome-reacted cells (ARC) and viability (non-viable cells, NVC) by immunofluorescence cytochemistry (see Methods). Number of cells expressed as % of total number of cells showing Tyr-P in any part of cell body or in head, were detected and reported as Tyr-P cells and Tyr-P head, respectively. Percentages of cells undergoing acrosome reaction (ARC) or NVC were also reported. Values are expressed as means ± SD.
† p < 0.01;
†† p < 0.001;
‡ p < 0.0001 comparison between various samples
vs. C as reference; Student’s
t-test for paired data. *
p < 0.05; **
p < 0.001; ***
p < 0.0001 comparison C
vs. T
0; Student’s
t-test for paired data.
Table 1.
Sperm cells from control group (CG) or patient group (PG) at T0, or incubated for 180 min in capacitating conditions in the absence (C) or presence of Asta 2 μM, PP2 6 μM or both (Asta + PP2), were analyzed for Tyr-P pattern (Supplementary Figure S2), acrosome-reacted cells (ARC) and viability (non-viable cells, NVC) by immunofluorescence cytochemistry (see Methods). Number of cells expressed as % of total number of cells showing Tyr-P in any part of cell body or in head, were detected and reported as Tyr-P cells and Tyr-P head, respectively. Percentages of cells undergoing acrosome reaction (ARC) or NVC were also reported. Values are expressed as means ± SD. † p < 0.01; †† p < 0.001; ‡ p < 0.0001 comparison between various samples vs. C as reference; Student’s t-test for paired data. * p < 0.05; ** p < 0.001; *** p < 0.0001 comparison C vs. T0; Student’s t-test for paired data.
| Groups | Parameters | T0 | C | Asta | PP2 | Asta + PP2 |
|---|
| CG | Tyr-P cells | 95 ± 3 | 93 ± 4 | 94 ± 5 | 30 ± 6 †† | 36 ± 7 ‡ |
| Tyr-P head | 10 ± 2 | 63 ± 6 *** | 67 ± 5 | 4 ± 2 ‡ | 5 ± 2 ‡ |
| ARC | 16 ± 5 | 59 ± 7 *** | 63 ± 6 | 5 ± 2 ‡ | 6 ± 1 ‡ |
| NVC | 7 ± 5 | 9 ± 3 | 5 ± 2 † | 69 ± 5 ‡ | 57 ± 7 †† |
| PG | Tyr-P cells | 48 ± 6 | 57 ± 5 | 64 ± 5 | 24 ± 6 ‡ | 29 ± 5 ‡ |
| Tyr-P head | 8 ± 2 | 19 ± 3 ** | 27 ± 3 ‡ | 2 ± 1 ‡ | 3 ± 1 †† |
| ARC | 10 ± 4 | 15 ± 4 ** | 31 ± 4 ‡ | 4 ± 2 †† | 4 ± 1 †† |
| NVC | 13 ± 4 | 19 ± 2 * | 12 ± 2 †† | 65 ± 8 ‡ | 58 ± 4 ‡ |
Aliquots of sperm suspensions were analyzed for percentages of ARC and NVC. The values of cell counts are reported in
Table 1. As previously reported [
19], the percentages of cells which underwent acrosome reaction (ARC) had a trend similar to the corresponding values of Tyr-P in acrosome/head region, once more confirming the correlation existing between these two parameters. Asta, ameliorating the percentage of Tyr-P cells in PG, increased ARC values in this group (31% ± 4%
vs. 15% ± 4%,
p < 0.0001) but in CG, Asta did not induce any increase, compared to C (63% ± 6%
vs. 59% ± 7%, respectively). PP2, both alone or in co-addition with Asta, prevented cells from AR, as indicated by the low percentages of ARC (5% ± 2% or 6% ± 1% compared to 59% ± 7% of the C sample in CG,
p < 0.0001; 4% ± 2% or 4% ± 1% compared to 15% ± 4% of the C sample in PG,
p < 0.001) and seriously compromised cell viability, as shown by the high number of NVC (69% ± 5% compared to 9% ± 3% of the C sample in CG,
p < 0.0001; 65% ± 8% compared to 19% ± 2% of the C sample in PG,
p < 0.001) also in the presence of Asta. Results showed that Asta was an optimal support for PG sperm to achieve capacitation status even in the absence of the correct ROS amount. Addition of Asta increased both Tyr-P and ARC percentages with significant increase of PG-C values (Tyr-P head 27% ± 3% in the presence of Asta, compared to 19% ± 3% of the corresponding sample C, and ARC 31% ± 4% compared to 15% ± 4%, in the presence or absence of Asta, respectively). When added, PP2 greatly augmented the percentages of NVC in both groups, which also showed a net decrease in all the other parameters. Asta did not succeed in reversing PP2 negative effects on sperm functioning and viability.
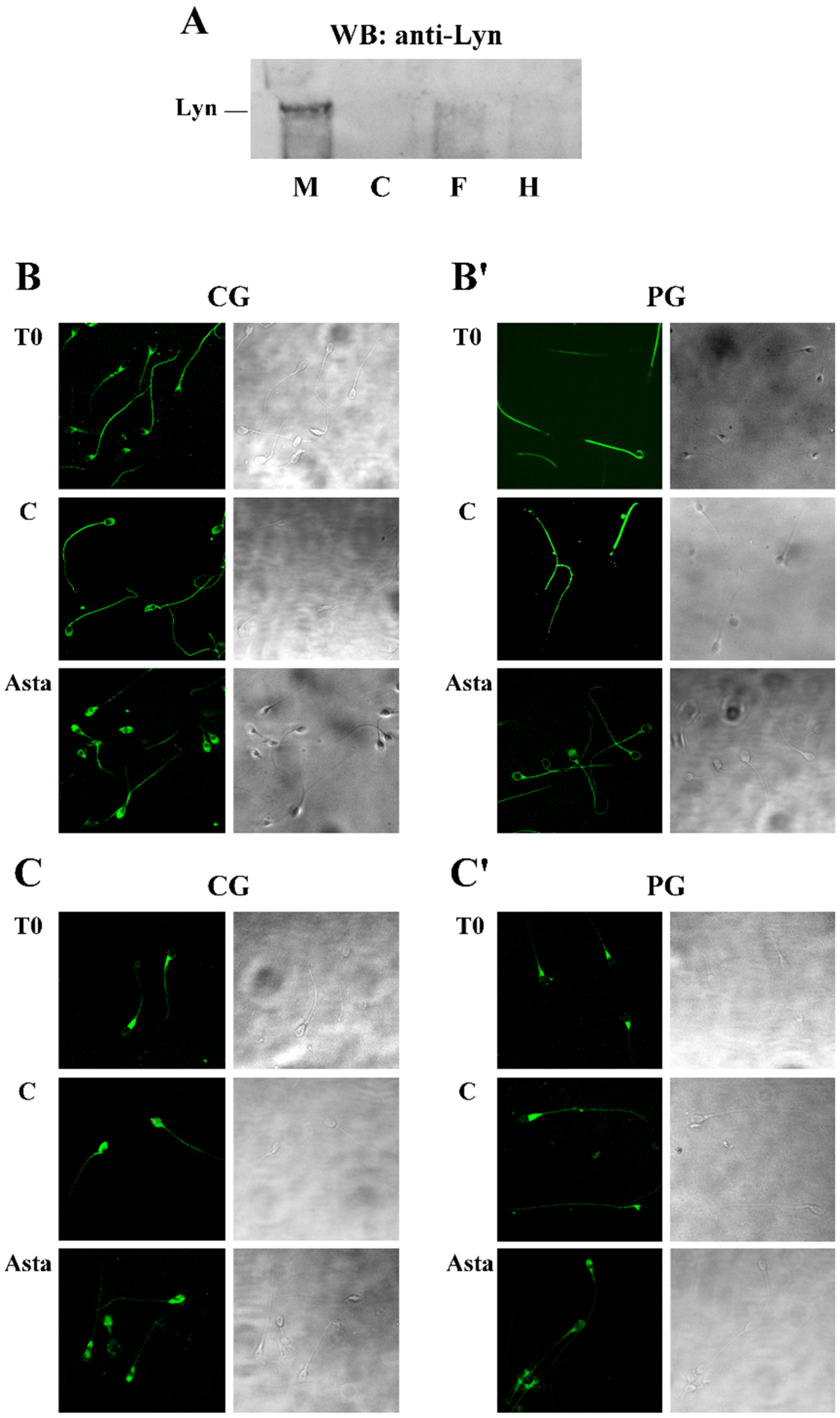
Taking into account that, in other cells, rafts contain Lyn, another member of the Src family kinase, we investigated the presence and the potential involvement of this enzyme in sperm functioning. In order to determine if the enzyme was expressed all over sperm surface or in precise cellular compartments, sperm was separated in membranes (M), cytosol (C), head (H) and flagellum (F), and successively analysed by anti-Lyn antibody in a Western blotting assay. As indicated by
Figure 2 panel A, Lyn was present in human sperm with a subcellular localization in the plasma M, enveloping the cell. For this reason, to further inquire about any potential alteration of Lyn position within the cells following treatments, differently treated sperm samples were analysed with anti-Lyn and visualized by confocal microscopy, to localize the enzyme (
Figure 2). Sperm was also inquired with CTB, which, by binding to membrane-inserted protein G1, allowed researchers to localize rafts and capacitation-related raft translocation [
13].
At T
0, Lyn was predominantly located at the neck of the sperm in both CG and PG, but after 180 min of capacitation, Lyn spread over sperm heads, locating predominantly to the acrosome region in CG, whereas, in PG, it remained in the neck or, at most, in the pre-acrosomal region, only rarely reaching the acrosome region (
Figure 2 and
Table 2 for quantification). Asta increased the percentage of cells from PG presenting both CTB and Lyn in the acrosome region, from 12% ± 2% to 28% ± 7% (CTB) and from 15% ± 2% to 22% ± 1% (Lyn) of samples C and Asta, respectively. These percentages accounted for what observed in PG Tyr-P and ARC (
Table 1). Interestingly, PP2, the inhibitor of Src kinase activity, did not cause any Lyn displacement alteration either in the absence or presence of Asta in PG and CG, thus confirming that Lyn activity was not a prerequisite for its translocation. However, even if Lyn spread over the acrosome region, a net inhibition of acrosome reaction was observed, as indicated by the percentage of ARC (5% ± 2% in PP2 and 6% ± 1% in Asta + PP2), compared to capacitated (C) samples (59% ± 7%).
The relative tyrosine kinase activities in samples from differently treated cells were also evaluated (
Table 2). Cells were lysed, and lysates were assayed as Protein Tyrosine Kinase (PTK) activity in the total cell lysate, or, after immuno-precipitation (Ip) with anti-Lyn antibodies (Lyn-Ip). Results showed that capacitating conditions (C) induced a net increase of both total lysate and Lyn activities in CG (+73% and +81%, respectively, compared to T
0 conditions), by far higher than those evidenced in PG (+24% and +21%, compared to T
0) (
Table 2). Asta addition induced further increase of both total lysate and Lyn activities in PG (+16% in both samples, compared to C,
p < 0.01), but not in CG.
Figure 2.
Subcellular localization of Lyn (panel A) and membrane rafts (panels B and B’) and Lyn (panels C and C’) translocation in human sperm during capacitation in absence or presence of Asta. (Panel A): Western blot analysis of subcellular localization of human sperm Lyn. The presence of the Src family kinase Lyn was assessed in the non-capacitated sperm plasma membrane (M), cytosol (C), flagellum (F) or head (H), obtained as described in Methods. Aliquots from each fraction corresponding to 3 × 106 cells were loaded and analysed by SDS-PAGE (10%), transferred to nitrocellulose and immuno-revealed with anti-Lyn antibody. (Panels B, B’, C, and C’): Sperm cells from control group (CG, panels B and C) and patient group (PG, panels B’ and C’), at T0 or incubated in capacitating conditions for 180 min in absence (C) or presence of Asta (2 μM), were analysed for CTB labelling (panels B and B’) and Lyn localization (panels C and C’) by immunofluorescence cytochemistry as described in Methods. Corresponding phase-contrast images for each condition are shown. The figure is representative of 11 separate experiments conducted in triplicate.
Figure 2.
Subcellular localization of Lyn (panel A) and membrane rafts (panels B and B’) and Lyn (panels C and C’) translocation in human sperm during capacitation in absence or presence of Asta. (Panel A): Western blot analysis of subcellular localization of human sperm Lyn. The presence of the Src family kinase Lyn was assessed in the non-capacitated sperm plasma membrane (M), cytosol (C), flagellum (F) or head (H), obtained as described in Methods. Aliquots from each fraction corresponding to 3 × 106 cells were loaded and analysed by SDS-PAGE (10%), transferred to nitrocellulose and immuno-revealed with anti-Lyn antibody. (Panels B, B’, C, and C’): Sperm cells from control group (CG, panels B and C) and patient group (PG, panels B’ and C’), at T0 or incubated in capacitating conditions for 180 min in absence (C) or presence of Asta (2 μM), were analysed for CTB labelling (panels B and B’) and Lyn localization (panels C and C’) by immunofluorescence cytochemistry as described in Methods. Corresponding phase-contrast images for each condition are shown. The figure is representative of 11 separate experiments conducted in triplicate.
![Marinedrugs 13 05533 g002]()
Table 2.
Membrane rafts localization, Lyn localization and activity in human sperm during capacitation and effects of Asta, PP2, or both. Sperm cells from control group (CG, panel A) and patient group (PG, panel B), at T0 or incubated in capacitating conditions for 180 min in absence (C) or presence of Asta 2 μM (Asta), PP2 6 μM (PP2) or both (Asta + PP2), were analysed for cholera toxin subunit B (CTB) labelling and Lyn localization by immunofluorescence cytochemistry as described in Methods. Number of cells, expressed as % of the total amount of cells showing labelling in acrosome, sub-acrosomal region (Sub-acro), neck and tail, were detected and reported. Protein Tyrosine Kinase (PTK) activity: aliquots of total sperm lysate or Lyn-Ip from each sample were analyzed for PTK activity as described in Methods. The values were obtained as the ratio percentage of PTK activity of samples to T0 (chosen as arbitrary comparison units). The figure is representative of 11 separate experiments. Values are expressed as means ± SD. † p < 0.05; †† p < 0.01; ‡ p < 0.001 comparison between various samples vs. C as reference; Student’s t-test for paired data. * p < 0.05; ** p < 0.01; *** p < 0.001 comparison C vs. T0; Student’s t-test for paired data.
Table 2.
Membrane rafts localization, Lyn localization and activity in human sperm during capacitation and effects of Asta, PP2, or both. Sperm cells from control group (CG, panel A) and patient group (PG, panel B), at T0 or incubated in capacitating conditions for 180 min in absence (C) or presence of Asta 2 μM (Asta), PP2 6 μM (PP2) or both (Asta + PP2), were analysed for cholera toxin subunit B (CTB) labelling and Lyn localization by immunofluorescence cytochemistry as described in Methods. Number of cells, expressed as % of the total amount of cells showing labelling in acrosome, sub-acrosomal region (Sub-acro), neck and tail, were detected and reported. Protein Tyrosine Kinase (PTK) activity: aliquots of total sperm lysate or Lyn-Ip from each sample were analyzed for PTK activity as described in Methods. The values were obtained as the ratio percentage of PTK activity of samples to T0 (chosen as arbitrary comparison units). The figure is representative of 11 separate experiments. Values are expressed as means ± SD. † p < 0.05; †† p < 0.01; ‡ p < 0.001 comparison between various samples vs. C as reference; Student’s t-test for paired data. * p < 0.05; ** p < 0.01; *** p < 0.001 comparison C vs. T0; Student’s t-test for paired data.
| A | | Localization | PTK Activity |
| | Treatment | Compartment | CTB (%) | Lyn (%) | Total Lysate | Lyn-Ip |
| CG | T0 | Acrosome | 0 ± 0 | 0 ± 0 | 100 ± 1 | 101 ± 2 |
| Sub-acro | 1 ± 1 | 4 ± 2 |
| Neck | 89 ± 8 | 96 ± 4 |
| Tail | 92 ± 4 | 11 ± 6 |
| C | Acrosome | 91 ± 8 *** | 93 ± 6 *** | 173 ± 9 *** | 181 ± 7 *** |
| Sub-acro | 91 ± 5 *** | 90 ± 8 *** |
| Neck | 92 ± 4 | 77 ± 6 *** |
| Tail | 78 ± 10 | 9 ± 4 |
| Asta | Acrosome | 95 ± 3 | 91 ± 5 | 178 ± 11 | 183 ± 8 |
| Sub-acro | 92 ± 2 | 93 ± 6 |
| Neck | 90 ± 6 | 72 ± 7 |
| Tail | 76 ± 7 | 10 ± 5 |
| PP2 | Acrosome | 31 ± 8 ‡ | 26 ± 8 ‡ | 29 ± 6 ‡ | 2 ± 1 ‡ |
| Sub-acro | 37 ± 11 †† | 29 ± 3 ‡ |
| Neck | 79 ± 10 † | 89 ± 7 |
| Tail | 78 ± 9 | 7 ± 3 |
| Asta + PP2 | Acrosome | 52 ± 7 ‡ | 34 ± 5 ‡ | 30 ± 5 ‡ | 1 ± 1 ‡ |
| Sub-acro | 54 ± 6 ‡ | 31 ± 6 ‡ |
| Neck | 85 ± 9 | 79 ± 5 |
| Tail | 84 ± 5 | 8 ± 4 |
| B | | Localization | PTK Activity |
| | Treatment | Compartment | CTB (%) | Lyn (%) | Total Lysate | Lyn-Ip |
| PG | T0 | Acrosome | 0 ± 0 | 0 ± 0 | 101 ± 1 | 102 ± 1 |
| Sub-acro | 0 ± 1 | 2 ± 1 |
| Neck | 75 ± 9 | 95 ± 4 |
| Tail | 93 ± 6 | 16 ± 5 |
| C | Acrosome | 12 ± 2 *** | 15 ± 2 *** | 125 ± 7 *** | 123 ± 5 *** |
| Sub-acro | 7 ± 2 *** | 6 ± 2 * |
| Neck | 89 ± 10 ** | 90 ± 6 |
| Tail | 93 ± 4 | 12 ± 5 |
| Asta | Acrosome | 28 ± 7 †† | 22 ± 1 †† | 141 ± 4 †† | 139 ± 6 †† |
| Sub-acro | 32 ± 9 †† | 21 ± 4 †† |
| Neck | 92 ± 7 | 96 ± 3 |
| Tail | 90 ± 6 | 13 ± 4 |
| PP2 | Acrosome | 9 ± 1 † | 8 ± 2 †† | 2 ± 1 ‡ | 1 ± 2 ‡ |
| Sub-acro | 5 ± 2 | 1 ± 1 † |
| Neck | 71 ± 9 † | 68 ± 11 †† |
| Tail | 84 ± 8 † | 9 ± 4 |
| Asta + PP2 | Acrosome | 6 ± 4 † | 9 ± 1 †† | 3 ± 1 ‡ | 1 ± 1 ‡ |
| Sub-acro | 7 ± 3 | 3 ± 2 |
| Neck | 82 ± 8 | 73 ± 7 † |
| Tail | 83 ± 6 † | 9 ± 3 |
These data strengthen the efficacy of Asta in ameliorating sperm unable to undergo correct capacitation leading to AR. On the other hand, normal sperm did not benefit from Asta positive effects for AR, probably due to the fact that sperm cells were working to the best of their capacity so that any further improvement was quite impossible. In any case, in both groups the presence of Asta in the incubation medium decreased the number of NVC (
Table 1), thus confirming the efficacy of this compound to prevent sperm apoptosis.
PP2, an inhibitor of the Src family kinase, induced cell denaturation leading to apoptosis. This confirmed the involvement of one or more members of this family, not only in the capacitating process but also in spermatozoa survival. Previous investigations [
32,
33] indicated p60src as responsible for capacitation progress in human sperm, though this enzyme was found predominantly in the neck, tail and pre-acrosome region. In the present study, Lyn, rather than p60src, better represents the key enzyme to the Tyr-P process involving sperm head. In fact, Lyn migrates from neck to the acrosome region, probably enveloped in raft caveolae/microdomains. Only when located in acrosome, Lyn activity highly increased, thus allowing the Tyr-P of the head.
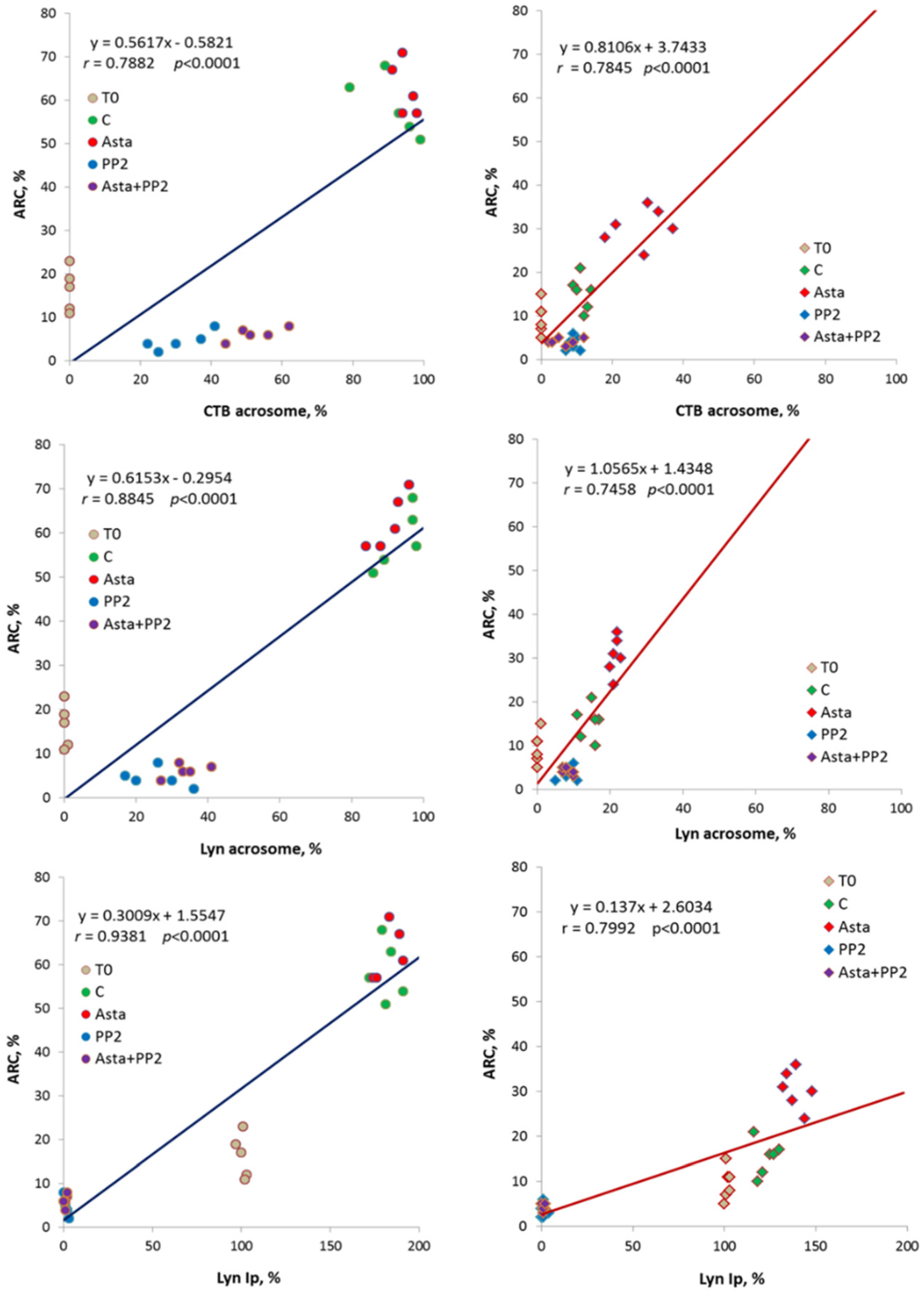
Statistical analysis indicated close relationships between acrosomal CTB, acrosomal Lyn and Lyn-Ip activity values
vs. ARC, as showed by the statistically significant linear regressions obtained through data points (
p < 0.0001) in both groups (
Figure 3). Moreover, the trend lines show clearly the phenomenon of increase in all the considered parameters following Asta treatment in PG.
Previous studies have demonstrated that AR closely depends on a controlled range of ROS generation leading to the Tyr-P process of the sperm head [
18,
19]. In sperm unable to produce them or in the presence of strong anti-oxidants, such as ascorbic acid, spermatozoa undergo a kind of general impairment, which prevents not only their ability to undergo AR but also cell survival [
18]. This close relation between the endogenous ROS content and sperm viability were further confirmed in the present study. In fact, when CG sperm was incubated in the presence of AA capacitation was prevented as shown by the inhibition of ROS production (
Supplementary Figure S1, panel A), Tyr-P and AR (
Supplementary Table S2). On the other hand, when PG sperm was treated in the presence of H
2O
2 (0.25 μM) to ameliorate ROS content (
Supplementary Figure S1, panel B) and trigger capacitation, no improvement either in Tyr-P level and distribution or ARC was shown (
Supplementary Table S2). In this case, it is hypothesized that the impairment present in PG sperm, being inserted in the membrane, was not achievable by exogenously added ROS.
Figure 3.
Correlations between acrosomal CTB, acrosomal Lyn and Lyn-Ip activity (Lyn-Ip %) values vs. ARC in both groups (CG on left, PG on right). Linear regression parameters are indicated, as well as Pearson’s coefficient of correlation r (p < 0.0001 in all cases).
Figure 3.
Correlations between acrosomal CTB, acrosomal Lyn and Lyn-Ip activity (Lyn-Ip %) values vs. ARC in both groups (CG on left, PG on right). Linear regression parameters are indicated, as well as Pearson’s coefficient of correlation r (p < 0.0001 in all cases).
In the present study, sperm samples were incubated in a buffer which guaranteed an optimal ROS production in normal CG as shown by the high ARC percentage but not sufficient to trigger capacitation in PG. In any case, low percentages of apoptosis (NVC) were ensured, and, more interestingly, viability impairment was prevented, as indicated by the net increase of ARC following Asta addition, thus indicating that cells were in a sort of resting conditions ready for a potential successive activation. This important aspect shed light also to the possible mechanism of action of Asta. In fact, the main difference evidenced in PG was the failure in raft shifting and relocation, compared to the high percentage of CG cells easily achieving capacitating membrane adjustment. By displacing rafts together with rafts-inserted-Lyn to the acrosome region, Asta markedly induced the conformational change required for AR, thus bypassing the impairment present in these cells for the capacitation-related arrangement. Possibly Asta, by inserting in the lipid bilayer, disengages rafts from the blocks that maintain the membrane in non-capacitated form, thus allowing both rafts and relative inserted proteins to reach the capacitated-related relocation. It is hypothesized that mechanisms other than cholesterol extraction may be involved in the membrane reorganization induced by capacitation, since Asta inserts itself into the bilayer, probably disrupting protein-protein or protein-lipid interactions, thus allowing/facilitating both the correct engagement of proteins in newly formed complexes, such as in the case of actin polymerization [
34], and the correct relocation of enzymes, such as Lyn, to the acrosome region. This Lyn relocation had to be accompanied by its activation in order to achieve correct level of Tyr-P. Only when Tyr-P process involved the acrosome region capacitation was accomplished, as demonstrated by the high percentage of ARC in the corresponding cells. Taken together, these results show for the first time the close relation between endogenous ROS correct generation and raft migration to the sperm head as “
sine qua non condicio” for the successive AR. In addition, Lyn, probably embedded and co-migrating with lipid rafts, is activated and catalyzes Tyr-P of proteins localized in the sperm head, third and further step of sperm preparation to achieve AR. When ROS content is insufficient [
13,
18] (PG), rafts cannot migrate, thus blocking successive phases, and this may be the limiting step for distinguishing PG from CG samples, even if routine test values fall into the normal range.




 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}

