2.3. Correlation between Plasma PL Marine-Derived n-3 LC PUFAs and Biochemical Parameters of Subjects
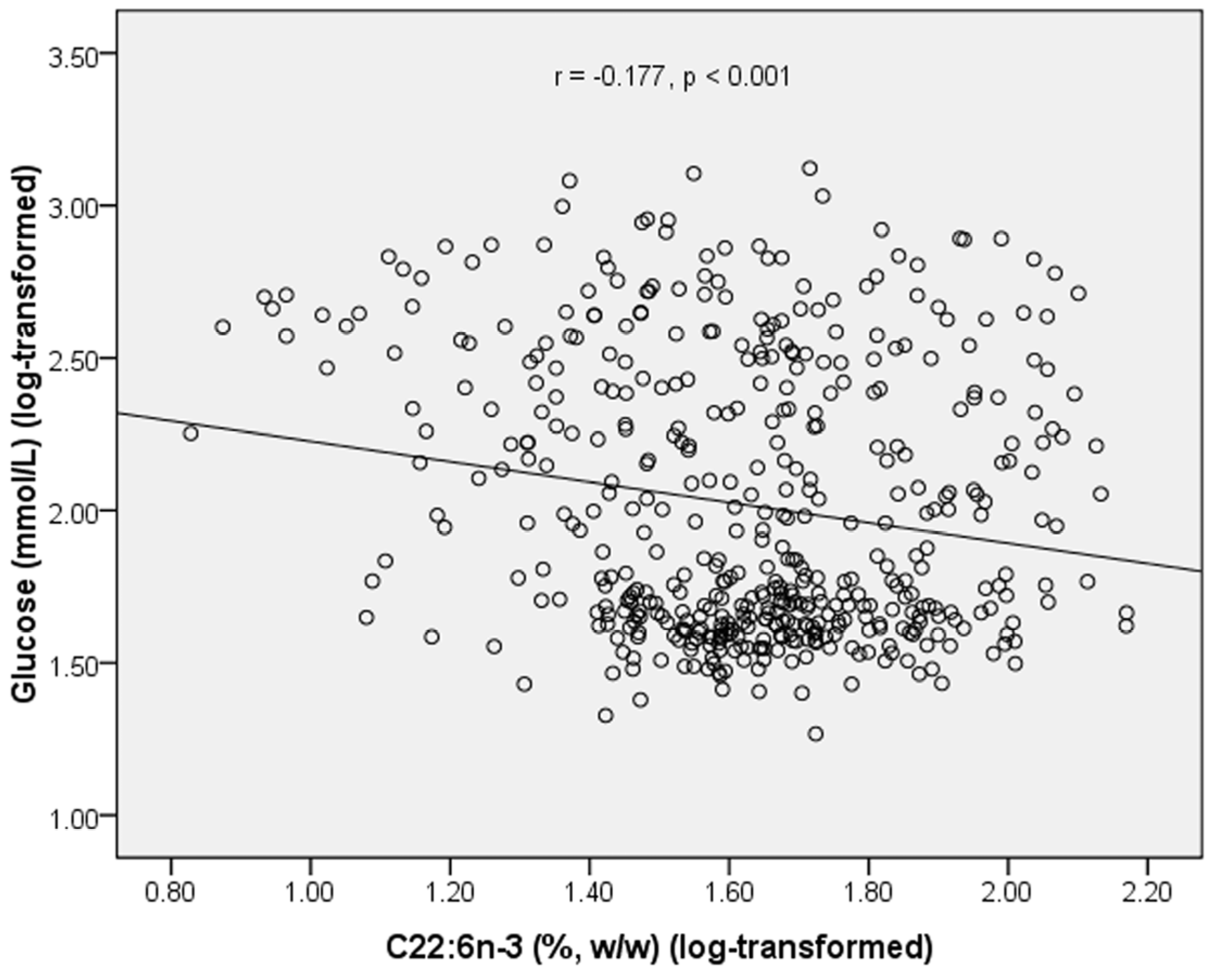
In 479 subjects, plasma PL C22:6
n-3 was negatively correlated with glucose level (
r = −0.177,
p < 0.001) (
Figure 1). Plasma PL C20:5
n-3 was positively correlated with HDL-C level (
r = 0.097,
p = 0.036) (
Table 3). Plasma level of UA was positively correlated with TG (
r = 0.320,
p < 0.001) and TC (
r = 0.156,
p < 0.001), and negatively correlated with glucose (
r = −0.239,
p < 0.001) and HDL-C (
r = −0.105,
p = 0.023) (
Table 3).
Figure 1.
Correlation between plasma C22:6n-3 and fasting glucose in all subjects.
Figure 1.
Correlation between plasma C22:6n-3 and fasting glucose in all subjects.
Table 3.
Pearson’s correlation coefficient between plasma marine-derived n-3 long chain polyunsaturated fatty acids (LC PUFAs) and biochemical parameters in all subjects.
Table 3.
Pearson’s correlation coefficient between plasma marine-derived n-3 long chain polyunsaturated fatty acids (LC PUFAs) and biochemical parameters in all subjects.
| | C20:5n-3 | C22:5n-3 | C22:6n-3 | Glucose | UA | TG | TC | HDL-C | LDL-C |
|---|
| C20:5n-3 | 1 | 0.187 *** | 0.380 *** | −0.032 | 0.082 | 0.044 | 0.048 | 0.097 * | 0.037 |
| C22:5n-3 | 0.187 *** | 1 | 0.187 *** | 0.006 | −0.013 | −0.084 | −0.030 | −0.040 | 0.037 |
| C22:6n-3 | 0.380 *** | 0.187 *** | 1 | −0.177 *** | 0.018 | 0.048 | 0.084 | 0.029 | 0.024 |
| Glucose | −0.032 | 0.006 | −0.177 *** | 1 | −0.239 *** | 0.074 | −0.121 ** | −0.262 *** | 0.153 *** |
| UA | 0.082 | −0.013 | 0.018 | −0.239 *** | 1 | 0.320 *** | 0.156 *** | −0.105 * | 0.040 |
| TG | 0.044 | −0.084 | 0.048 | 0.074 | 0.320 *** | 1 | 0.349 *** | −0.284 *** | 0.278 *** |
| TC | 0.048 | −0.030 | 0.084 | −0.121 ** | 0.156 *** | 0.349 *** | 1 | 0.357 *** | 0.715 *** |
| HDL-C | 0.097* | −0.040 | 0.029 | −0.262 *** | −0.105 * | −0.284 *** | 0.357 *** | 1 | 0.072 |
| LDL-C | 0.037 | 0.037 | 0.024 | 0.153 *** | 0.040 | 0.278 | 0.715 *** | 0.072 | 1 |
We also analyzed the correlation between plasma PL
n-3 LC PUFAs and biochemical parameters in healthy subjects and T2DM subjects separately (
Table 4 and
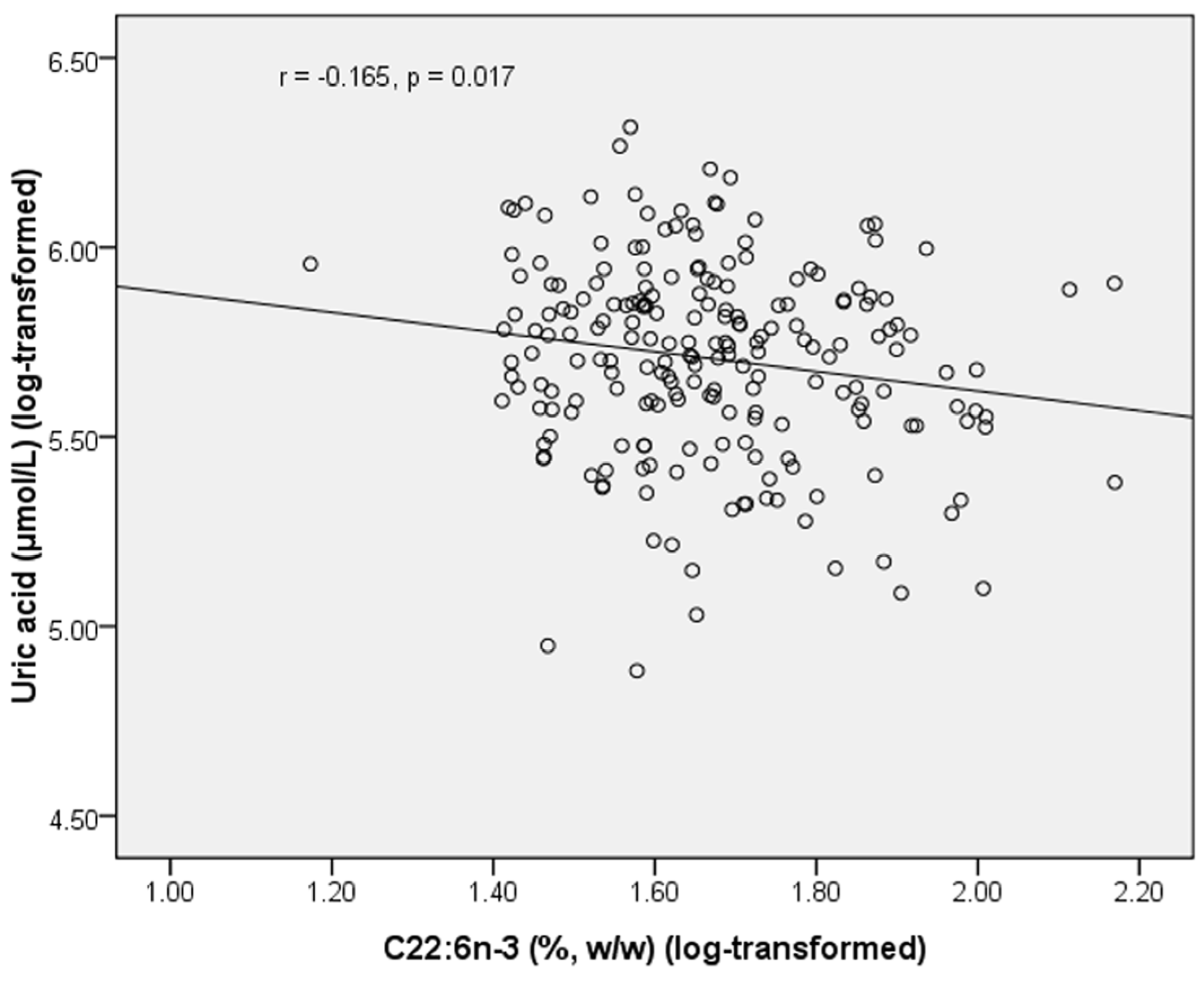
Table 5). In healthy subjects, PL C22:6
n-3 was negatively correlated with uric acid (
r = −0.165,
p =0.017) (
Figure 2). In T2DM subjects, PL C20:5
n-3 was negatively correlated with glucose (
r = −0.158,
p = 0.011), and positively correlated with UA (
r = 0.136,
p = 0.029) and HDL-C (
r = 0.131,
p = 0.036). In T2DM subjects, C22:5
n-3 was negatively correlated with UA (
r = −0.124,
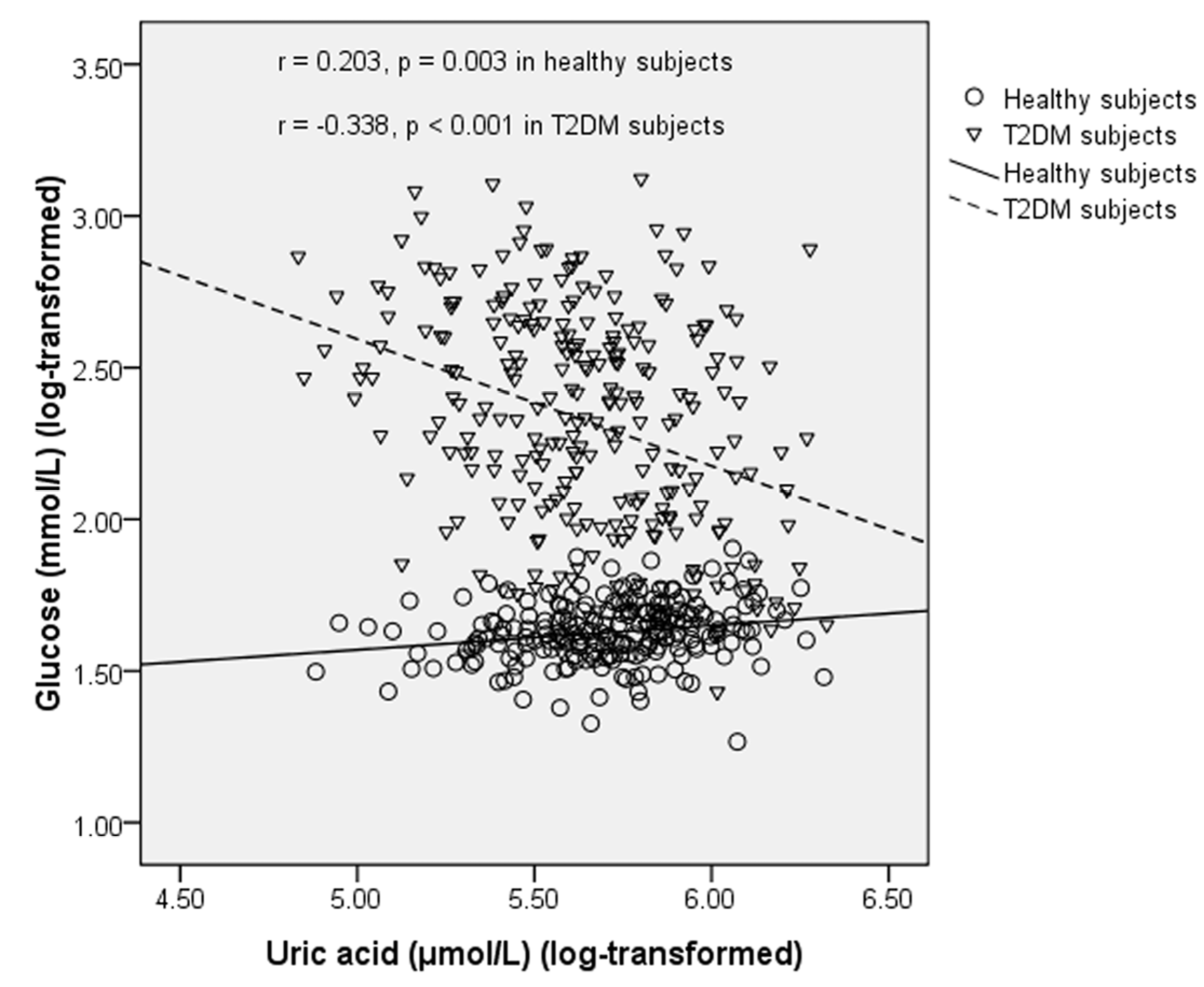
p = 0.044). UA was positively correlated with glucose in healthy subjects (
r = 0.203,
p = 0.003), but this correlation became negative in T2DM subjects (
r = −0.338,
p < 0.001) (
Figure 3). UA was positively correlated with TG in both healthy subjects (
r = 0.429,
p < 0.001) and T2DM subjects (
r = 0.278,
p < 0.001). UA was positively correlated with TC in both healthy subjects (
r = 0.168,
p = 0.015) and T2DM subjects (
r = 0.122,
p = 0.049). UA was negatively correlated with HDL-C (
r = −0.279,
p < 0.001) and positively correlated with LDL-C (
r = 0.168,
p = 0.014) in healthy subjects but these correlations became non-significant in T2DM subjects.
Table 4.
Pearson’s correlation coefficient between plasma marine-derived n-3 long chain polyunsaturated fatty acids (LC PUFAs) and biochemical parameters in healthy subjects.
Table 4.
Pearson’s correlation coefficient between plasma marine-derived n-3 long chain polyunsaturated fatty acids (LC PUFAs) and biochemical parameters in healthy subjects.
| | C20:5n-3 | C22:5n-3 | C22:6n-3 | GLU | UA | TG | TC | HDL-C | LDL-C |
|---|
| C20:5n-3 | 1 | 0.097 | 0.111 | 0.078 | 0.01 | −0.057 | 0.121 | 0.114 | 0.039 |
| C22:5n-3 | 0.097 | 1 | 0.016 | −0.014 | 0.137 | −0.057 | 0.001 | −0.031 | 0.058 |
| C22:6n-3 | 0.111 | 0.016 | 1 | 0.01 | −0.165 * | −0.016 | 0.124 | −0.032 | 0.098 |
| GLU | 0.078 | −0.014 | 0.01 | 1 | 0.203 ** | 0.13 | 0.004 | −0.012 | 0.011 |
| UA | 0.01 | 0.137 | −0.165 * | 0.203 ** | 1 | 0.429 *** | 0.168 * | −0.279 *** | 0.168 * |
| TG | −0.057 | −0.057 | −0.016 | 0.13 | 0.429 *** | 1 | 0.299 *** | −0.491 *** | 0.181 ** |
| TC | 0.121 | 0.001 | 0.124 | 0.004 | 0.168 * | 0.299 *** | 1 | 0.169 * | 0.841 *** |
| HDL-C | 0.114 | −0.031 | −0.032 | −0.012 | −0.279 *** | −0.491 *** | 0.169 * | 1 | 0.009 |
| LDL-C | 0.039 | 0.058 | 0.098 | 0.011 | 0.168* | 0.181 ** | 0.841 *** | 0.009 | 1 |
Table 5.
Pearson’s correlation coefficient between plasma marine-derived n-3 long chain polyunsaturated fatty acids (LC PUFAs) and biochemical parameters in Type 2 diabetes mellitus subjects.
Table 5.
Pearson’s correlation coefficient between plasma marine-derived n-3 long chain polyunsaturated fatty acids (LC PUFAs) and biochemical parameters in Type 2 diabetes mellitus subjects.
| | C20:5n-3 | C22:5n-3 | C22:6n-3 | GLU | UA | TG | TC | HDL-C | LDL-C |
|---|
| C20:5n-3 | 1 | 0.256 *** | 0.526 *** | −0.158 * | 0.136 * | 0.115 | 0.027 | 0.131 * | 0.026 |
| C22:5n-3 | 0.256 *** | 1 | 0.284 *** | 0.042 | −0.124 * | −0.105 | −0.055 | −0.066 | 0.027 |
| C22:6n-3 | 0.526 *** | 0.284 *** | 1 | −0.117 | 0.066 | 0.112 | 0.038 | −0.027 | 0.032 |
| GLU | −0.158 * | 0.042 | −0.117 | 1 | −0.338 *** | −0.068 | 0.054 | 0.064 | 0.063 |
| UA | 0.136 * | −0.124 * | 0.066 | −0.338 *** | 1 | 0.278 *** | 0.122 * | −0.083 | 0.006 |
| TG | 0.115 | −0.105 | 0.112 | −0.068 | 0.278 *** | 1 | 0.444 *** | −0.054 | 0.327 *** |
| TC | 0.027 | −0.055 | 0.038 | 0.054 | 0.122 * | 0.444 *** | 1 | 0.405 *** | 0.749 *** |
| HDL-C | 0.131 * | −0.066 | −0.027 | 0.064 | −0.083 | −0.054 | 0.405 *** | 1 | 0.224 *** |
| LDL-C | 0.026 | 0.027 | 0.032 | 0.063 | 0.006 | 0.327 *** | 0.749 *** | 0.224 *** | 1 |
Figure 2.
Correlation between plasma C22:6n-3 and uric acid in healthy subjects.
Figure 2.
Correlation between plasma C22:6n-3 and uric acid in healthy subjects.
Figure 3.
Correlation between uric acid and fasting blood glucose in healthy subjects and T2DM subjects.
Figure 3.
Correlation between uric acid and fasting blood glucose in healthy subjects and T2DM subjects.
2.6. Discussion
In the present case-control study, we assessed the association between plasma PL fatty acid composition and blood glucose and lipid metabolism in Chinese Hans. We also assessed the interaction between plasma PL marine-derived n-3 LC PUFAs and UA on glucose metabolism and the risk of T2DM. We found that higher plasma PL C22:6n-3 percentage was associated with a lower risk of T2DM. UA showed positive correlation with fasting glucose levels in healthy subjects, but the correlation became negative in T2DM subjects. A significant interaction was observed between PL C22:6n-3 and UA on fasting glucose levels, and this interaction has not been reported by previous studies.
In the present study, plasma PL C22:6
n-3 was significantly lower in the case group than in the control group, and higher PL C22:6
n-3 was associated with a lower risk of T2DM. This result was consistent with our previous case-control study in Chinese subjects [
1]. Our previous meta-analysis also found that dietary fish intake or marine-derived
n-3 PUFA supplementation can significantly decrease the risk of T2DM in Asian populations [
3]. The negative association between PL C22:6
n-3 and risk of T2DM may be attributed to its beneficial effect on glucose and UA metabolism. T2DM is characterized by high blood glucose and insulin resistance. In the present study, we found that plasma PL C22:6
n-3 was negatively correlated with fasting glucose level. A negative correlation between plasma PL C22:6
n-3 and UA level was also identified in this study. Previous studies have found that UA is an independent risk factor for T2DM [
5,
6]. In the present study, we also found that UA was positively correlated with TG and TC in both healthy subjects and T2DM subjects; UA was positively correlated with fasting glucose, but negatively correlated with HDL-C in healthy subjects. HDL-C had a protective role against insulin resistance [
13,
14]. High TG level has been reported to be associated with insulin resistance [
15,
16]. Therefore, results in the present study also indicate the adverse effect of UA on T2DM.
Our result in healthy subjects showed that UA was positively correlated with blood glucose level. This may be attributed to the increasing effect of UA on insulin resistance [
17]. Previous studies also found a negative association between UA and insulin secretion by β-cell in nondiabetic subjects [
18]. This may also help explain the positive association between UA and blood glucose. However, the increasing effect of UA on the risk of T2DM observed in previous cohort studies [
5,
19] did not recur in the present study. On the contrary, UA was significantly lower in the case group than in the control group. This may be attributed to the osmotic diuresis caused by high blood glucose levels in T2DM, that is, the osmotic diuresis in T2DM increased the clearance rate of UA and thus lowered UA levels. In the present study, we indeed observed that UA was negatively correlated with glucose in T2DM subjects. Another reason may be that high UA levels increased the function of β cell and thus lowered blood glucose. One previous study in T2DM subjects found that UA could increase glucose disposition indices DI30 and DI120 (indices used to assess β-cell function, which combined insulin secretion and insulin resistance together) by increasing insulin secretion, although it also increased insulin resistance [
20]. This may be regarded as a feedback mechanism of the human body to reduce the adverse effects of high blood glucose. However, this increased insulin secretion may accelerate the decay of β-cell function [
20]. The opposite effect of UA on insulin secretion by β-cell in nondiabetic subjects and T2DM subjects observed in previous studies [
18,
20] may help explain why UA was positively correlated with glucose in healthy subjects but negatively correlated with glucose in T2DM subjects and why UA was lower in T2DM subjects than in healthy subjects.
Previous studies have reported the interaction between UA and prehypertension, triglyceride as well as vitamin D
3 on chronic diseases [
10,
11,
12]. However, the interaction between marine-derived LC
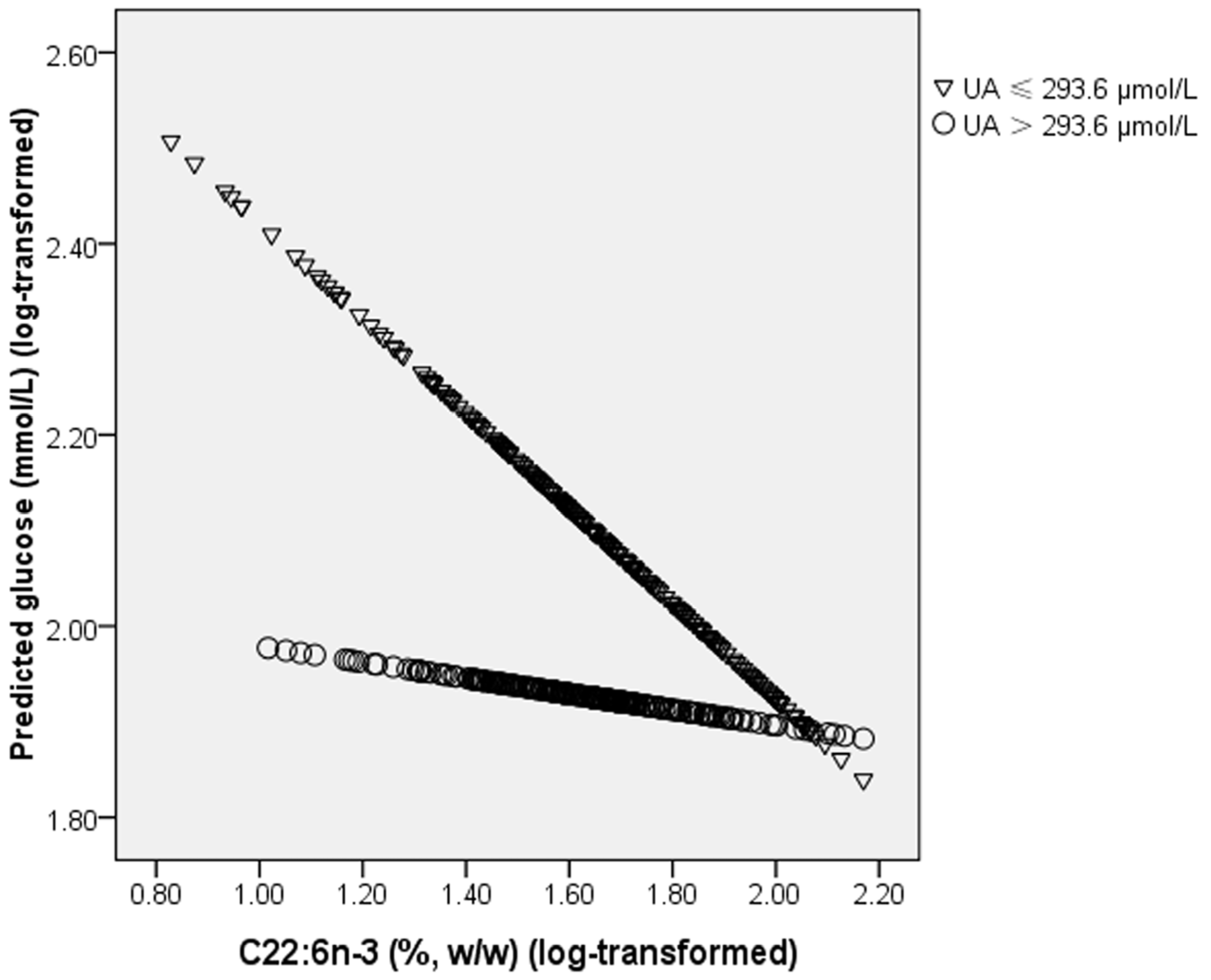
n-3 PUFAs on chronic diseases has not been reported. In the present study, we first identified an interaction between plasma PL C22:6
n-3 and UA on fasting glucose levels: the lowering effect of C22:6
n-3 on blood glucose was only observed in subjects with a lower level of UA. Insulin receptor substrate 1/2 (IRS-1/2) plays an important role in insulin signal transduction [
21]. Tyrosine phosphorylation of IRS leads to Src homology 2 (SH2) domain proteins binding to IRS, activates serine/threonine-specific protein kinase (AKT) and modulates glucose level in sequence [
22]. A previous study in mice found that C22:6
n-3 could significantly increase the expression of IRS-1 and IRS-2 in adipose tissue and liver [
8]. This can help explain the beneficial effect of C22:6
n-3 on glucose metabolism. One previous study found that high UA levels significantly increased serine phosphorylation of IRS-1 in mouse liver, muscle, and adipose tissue [
9]. Serine phosphorylation of IRS-1 can decrease its threonine phosphorylation, and thus block the insulin signal transduction [
23]. Therefore, when subjects had a higher UA level, the increased serine phosphorylation of IRS-1 counteracted the increasing effect of C22:6
n-3 on IRS-1 expression. This can help to explain why the lowering effect of C22:6
n-3 on glucose levels was only significant in subjects with a lower level of UA but not in subjects with a higher level of UA.
Interestingly, in the present study, the association of C20:5
n-3 and C22:5
n-3 with fasting glucose and the risk of T2DM, and the interaction between the two marine-derived LC PUFAs with UA on fasting glucose and the risk of T2DM were not significant. As mentioned in the paragraph, the effect of C22:6
n-3 and UA and their interaction on glucose may be attributed to their modulating effect on insulin signal transduction pathway involving IRS-1 and IRS-2. However, previous study in rats found that C20:5
n-3 supplementation also exhibited anti-hyperglycemic effects by increased insulin secretion, glycogen synthesis, and expression of IRS-1 [
24]. The increasing effect of C20:5
n-3 on expression of IRS-1 was also observed in a study in hepatoma cells [
25]. These results seem inconsistent with our study. Two reasons may exist for the different results observed for the three marine-derived LC PUFAs in the present study. On one hand, C22:6
n-3 is the major
n-3 PUFA in plasma phospholipids, and the content of C22:6
n-3 was more than four times of C20:5
n-3 or C22:5
n-3 in the present study. Therefore, C22:6
n-3 in plasma phospholipids may have a greater influence on glucose than C20:5
n-3 and C22:5
n-3, and thus the association of C20:5
n-3 and C22:5
n-3 with glucose may be hidden by the variation of glucose between subjects caused by C22:6
n-3. On the other hand, C20:5
n-3 can be metabolized to C22:5
n-3 and then to C22:6
n-3
in vivo [
26]. Although supplementation of C20:5
n-3 can increase the expression of IRS-1 and thus exhibit anti-hyperglycemic effect, whether the beneficial effect on glucose control was induced by C20:5
n-3 directly or by C22:6
n-3 synthesized from C20:5
n-3 is still unknown. Therefore, if the modulating effect of C20:5
n-3 or C22:5
n-3 supplementation on glucose was attributed to C22:6
n-3 synthesized from them, the association of C20:5
n-3 and C22:5
n-3 with fasting glucose and the risk of T2DM, and the interaction between the two marine-derived LC PUFAs with UA on fasting glucose and the risk of T2DM may become non-significant, because C22:6
n-3 in plasma can also be ingested from food directly.
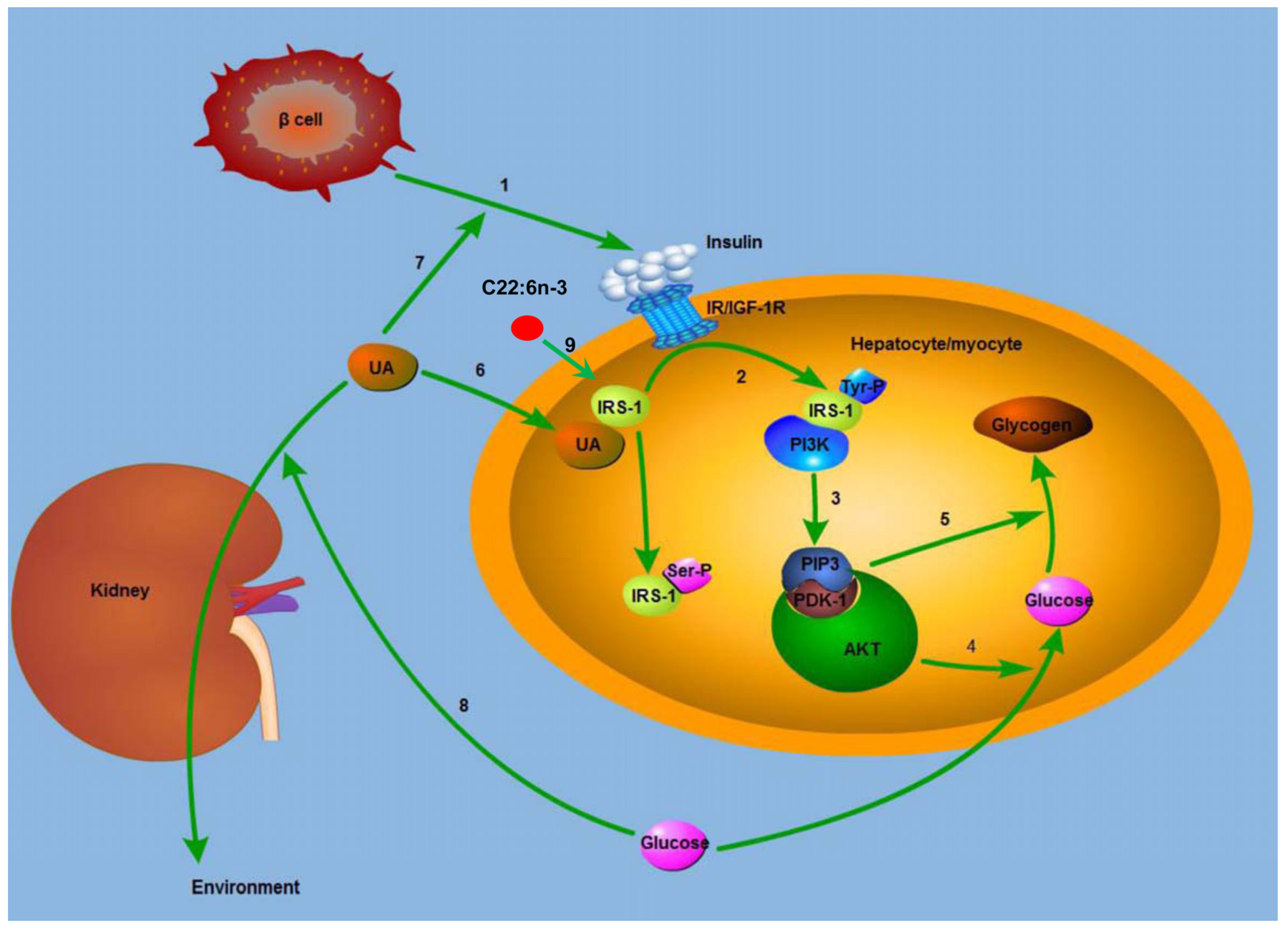
Figure 5.
Mechanism for the modulating effect of C22:6n-3 and uric acid on glucose metabolism. IRS-1, insulin receptor substrate 1; IR, insulin receptor; IGF-1R, insulin growth factor 1 receptor; Tyr, tyrosine; Ser, serine; PI3K, phosphatidylinositol 3-kinase; PIP3, phosphatidylinositol 3,4,5-trisphosphate; PDK1, 3-phosphatidylinositol-dependent kinase; AKT, protein kinase B; UA, uric acid. (1) β cell secreted insulin, and insulin combined with IR or IGF-1R; (2) the complex of insulin and IR lead to the Tyr-phosphorylation of IRS-1, and the Tyr-phosphorylated IRS-1 binds to PI3K; (3) the complex of Tyr-phosphorylated IRS-1 and PI3K generated PIP3, which recruited AKT, and AKT is activated by PDK1; (4) activated AKT increased the uptake of glucose; (5) AKT increased the synthesis of glycogen; (6) UA leads to the Ser-phosphorylation of IRS-1, and thus blocked the insulin signal pathway; (7) UA increases the secretion of insulin by β cell in T2DM subjects, and thus lowered blood glucose; (8) high blood glucose leads to osmotic diuresis, increases the clearance rate of UA, and decreases plasma UA level in sequence; (9) C22:6n-3 increases the expression of IRS-1, and in this way lowers blood glucose.
Figure 5.
Mechanism for the modulating effect of C22:6n-3 and uric acid on glucose metabolism. IRS-1, insulin receptor substrate 1; IR, insulin receptor; IGF-1R, insulin growth factor 1 receptor; Tyr, tyrosine; Ser, serine; PI3K, phosphatidylinositol 3-kinase; PIP3, phosphatidylinositol 3,4,5-trisphosphate; PDK1, 3-phosphatidylinositol-dependent kinase; AKT, protein kinase B; UA, uric acid. (1) β cell secreted insulin, and insulin combined with IR or IGF-1R; (2) the complex of insulin and IR lead to the Tyr-phosphorylation of IRS-1, and the Tyr-phosphorylated IRS-1 binds to PI3K; (3) the complex of Tyr-phosphorylated IRS-1 and PI3K generated PIP3, which recruited AKT, and AKT is activated by PDK1; (4) activated AKT increased the uptake of glucose; (5) AKT increased the synthesis of glycogen; (6) UA leads to the Ser-phosphorylation of IRS-1, and thus blocked the insulin signal pathway; (7) UA increases the secretion of insulin by β cell in T2DM subjects, and thus lowered blood glucose; (8) high blood glucose leads to osmotic diuresis, increases the clearance rate of UA, and decreases plasma UA level in sequence; (9) C22:6n-3 increases the expression of IRS-1, and in this way lowers blood glucose.
![Marinedrugs 13 05564 g005]()
The present study had several limitations. T2DM subjects may take some medications to treat hyperglycemia, hyperlipidemia, or hyperuricemia. The use of these medications in T2DM subjects may be a potential confounding factor for our results. However, information of medications use was not available in the present study, which may bias our final results. Well-designed randomized controlled trials are still needed in the future, on one hand to exclude the influence of medications use on the results, and on the other hand to see whether supplementation of C22:6n-3 showed different effect on glucose control in subjects with different UA level. In addition, subjects in the case group were significantly older than those in the control group, and age may bias our results. Therefore, we adjusted our primary results, the interaction between marine-derived n-3 LC PUFAs and UA on glucose and risk of T2DM and the association of marine-derived n-3 LC PUFAs and UA with risk of T2DM, for age in a multivariate linear regression model and a logistic regression model. However, after adjusting for age and sex, the interaction between marine-derived n-3 LC PUFAs and UA on glucose and the association between C22:6n-3 and risk of T2DM still remained significant, indicating that these significant results were independent of age and sex.
The potential mechanism for the modulating effect of C22:6
n-3 and UA on glucose metabolism is shown in
Figure 5.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




