
A Phospholipid-Protein Complex from Antarctic Krill Reduced Plasma Homocysteine Levels and Increased Plasma Trimethylamine-N-Oxide (TMAO) and Carnitine Levels in Male Wistar Rats

Abstract
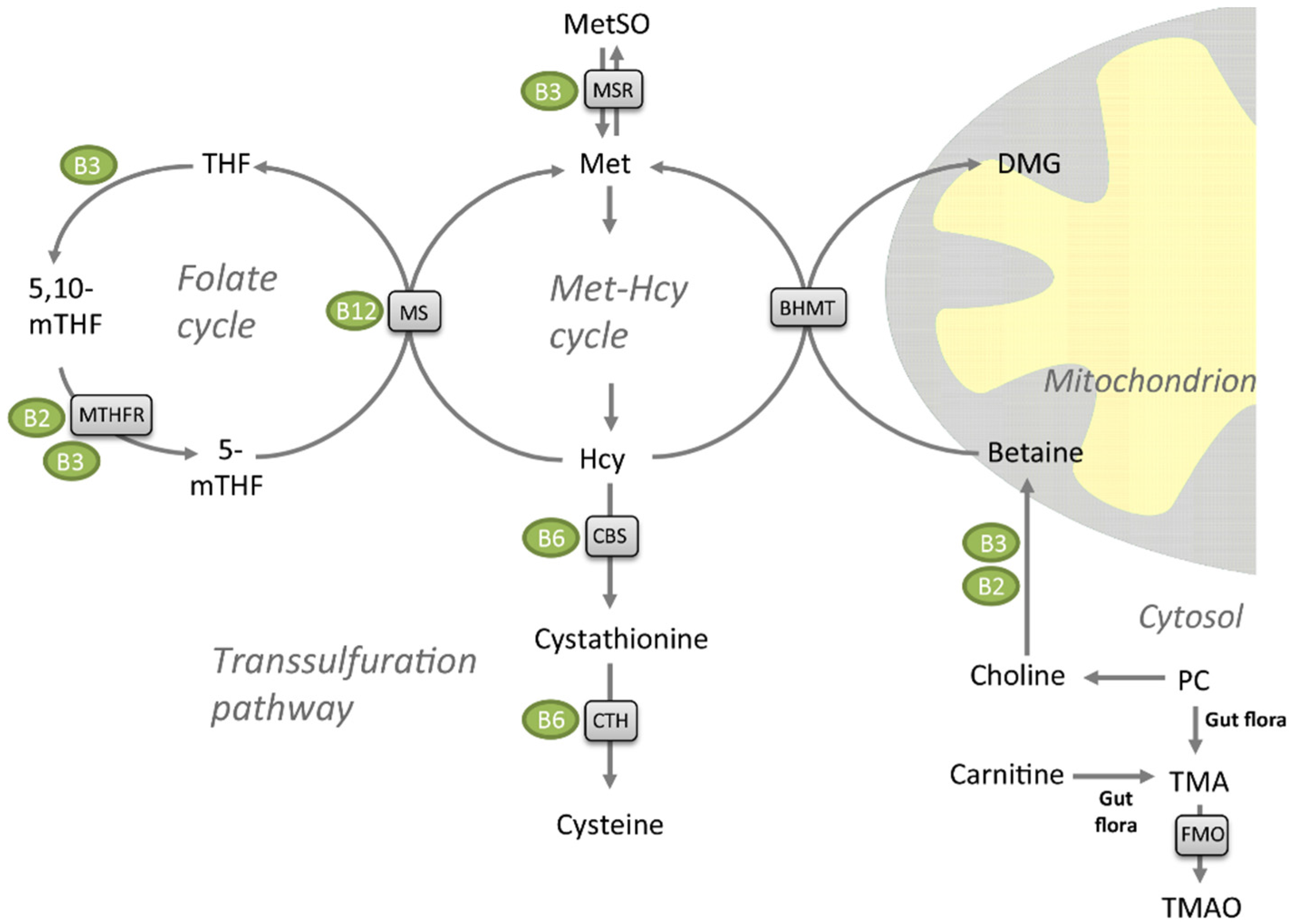
:1. Introduction

2. Results
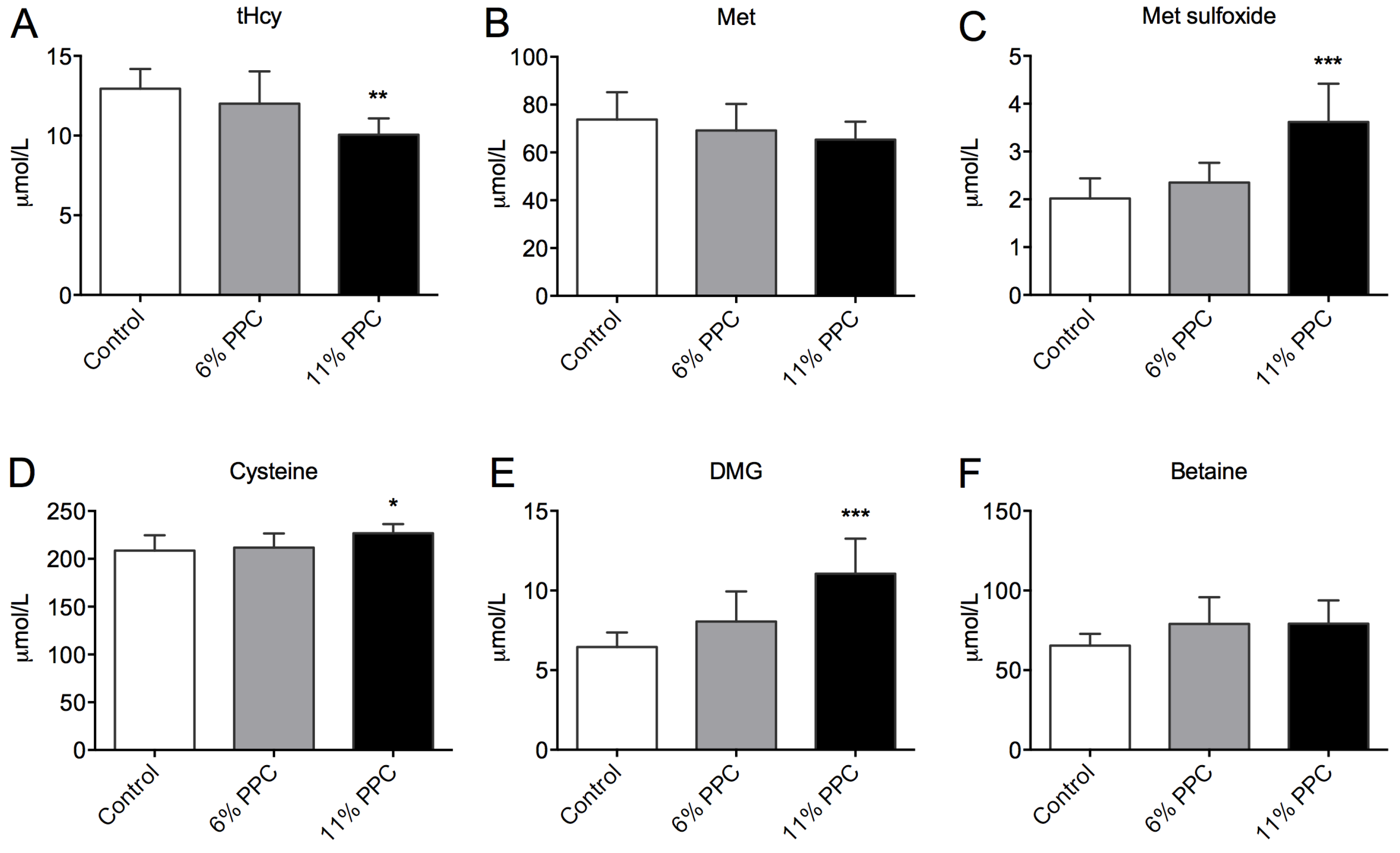
2.1. Metabolites of the Methionine-Homocysteine Cycle

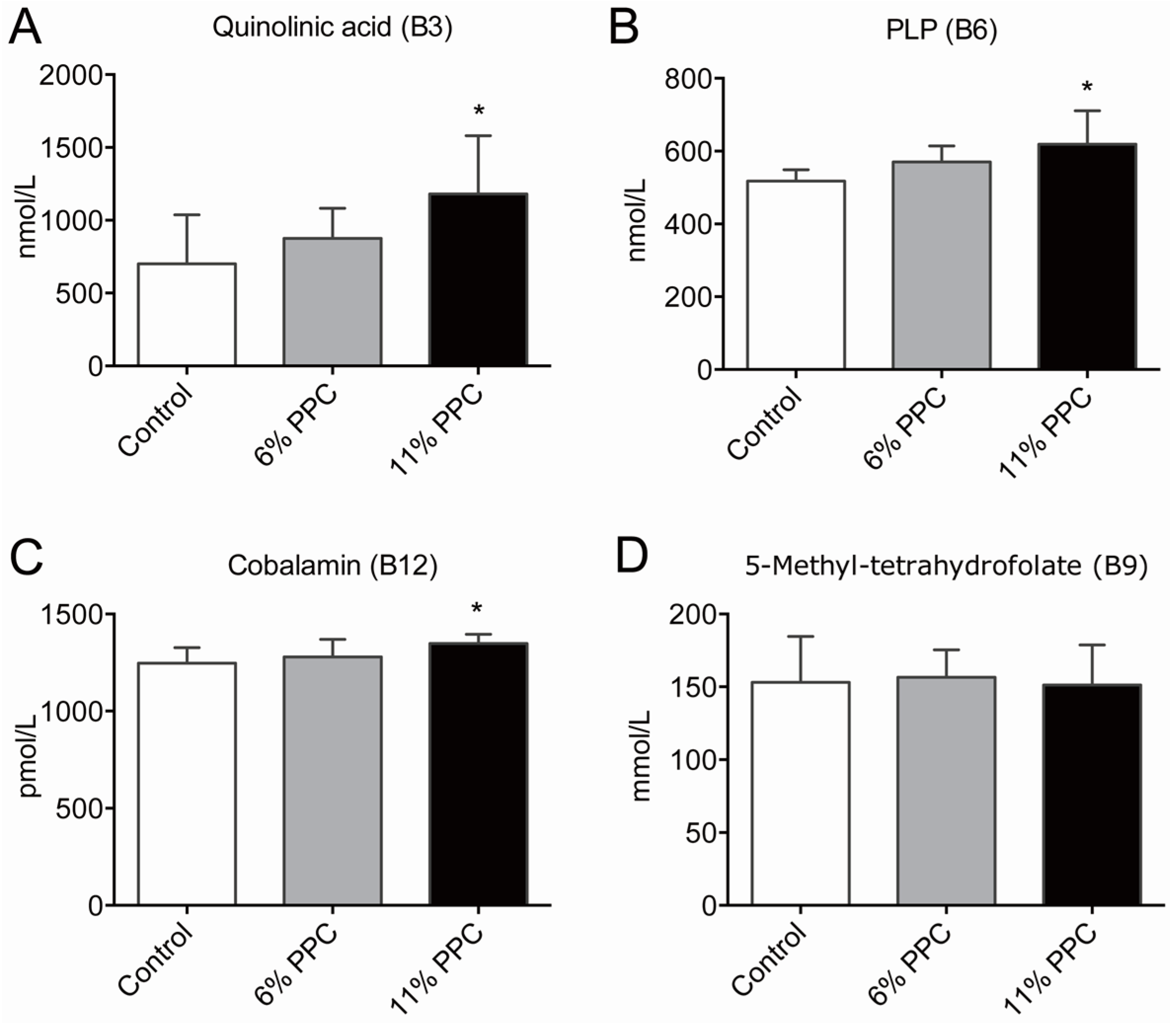
2.2. Vitamin B Levels

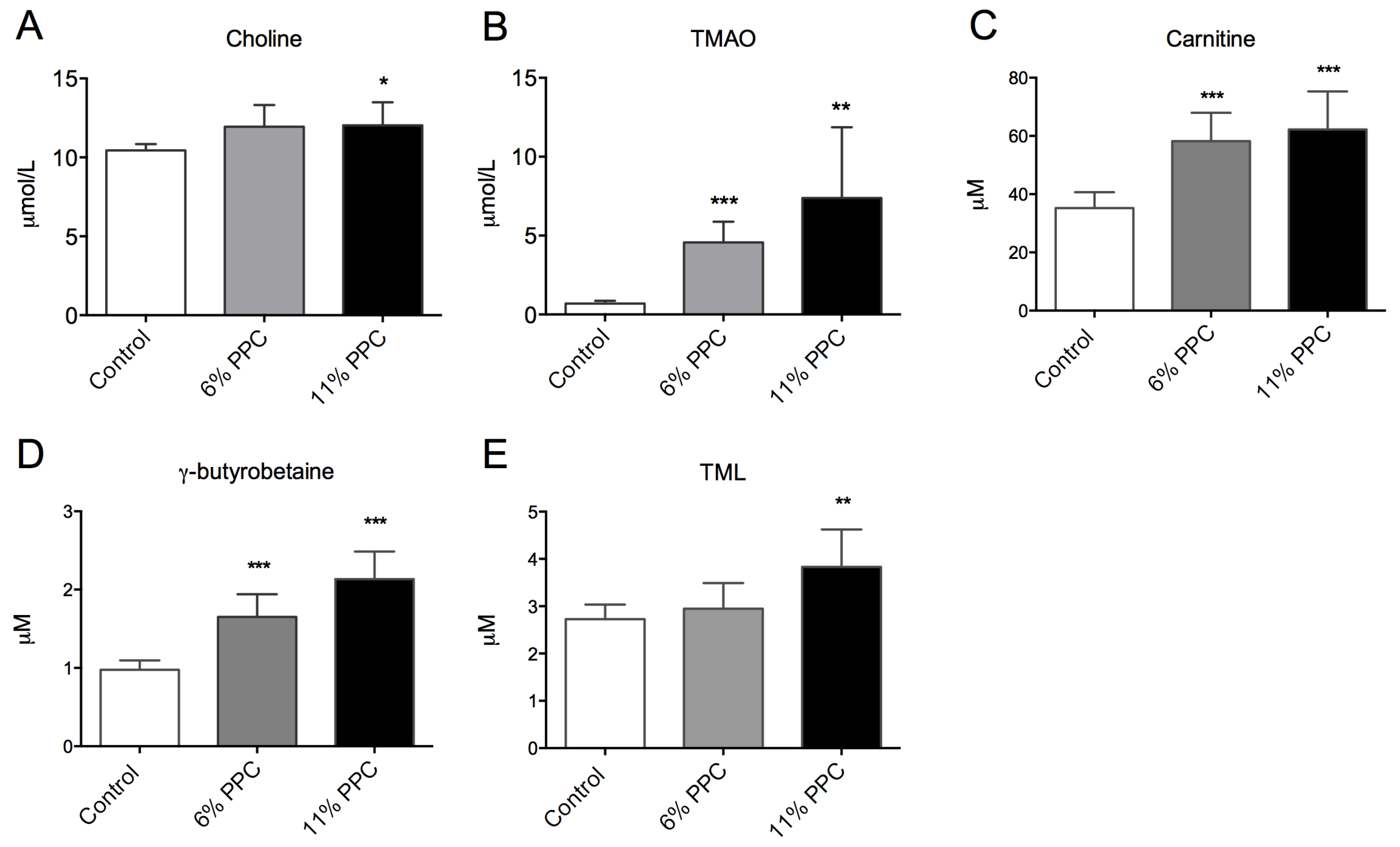
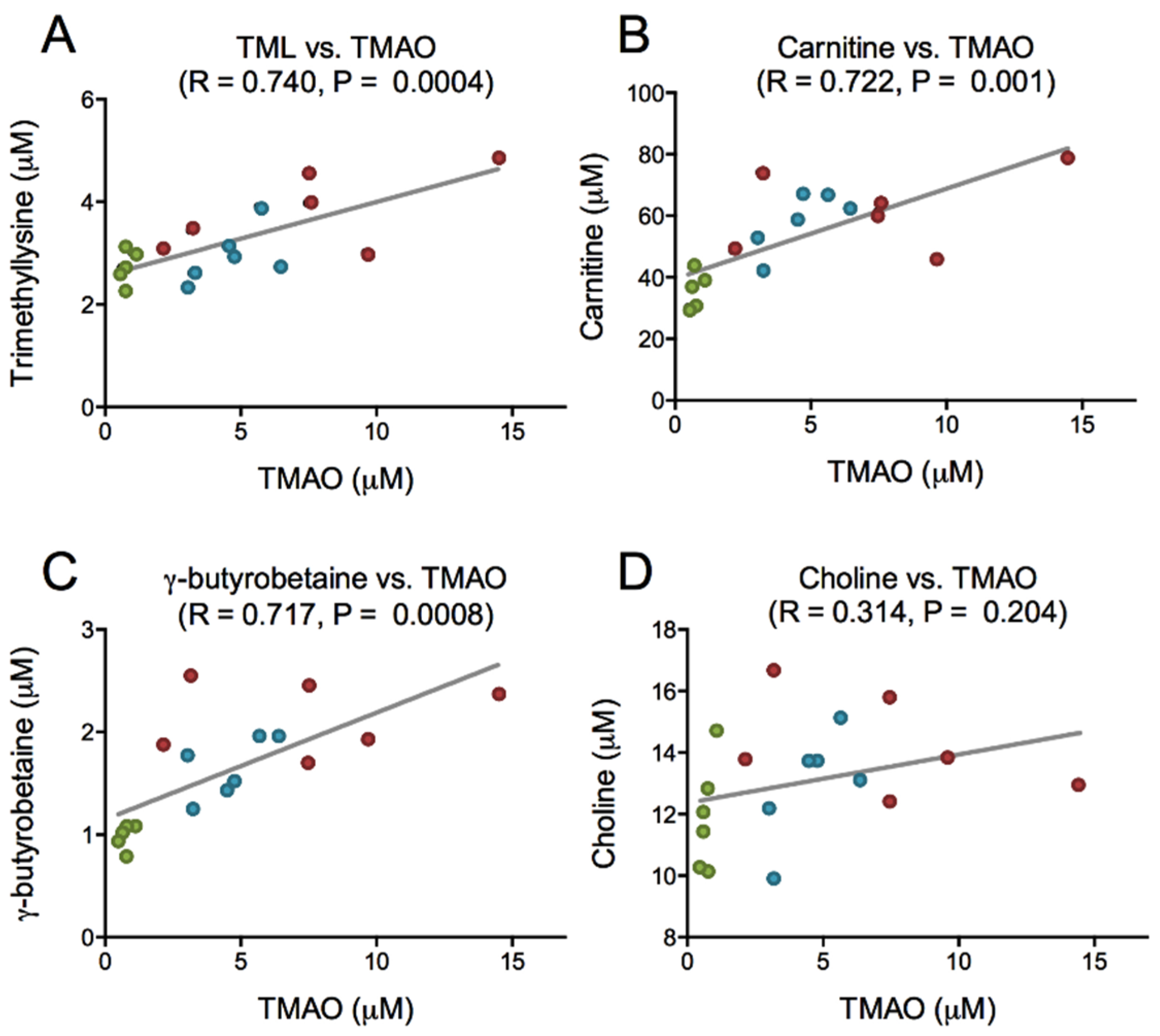
2.3. Choline, Carnitine, Carnitine Precursors and TMAO

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Compound | Control 1 | 11% PPC |
|---|---|---|
| TMAO | <0.001 | 12.6 |
| Free choline | 17.1 | 20.9 |
| Betaine | <0.001 | 3.9 |
| Carnitine | 0.25 | 0.27 |
| γ-Butyrobetaine | <0.001 | 0.009 |
| TML | <0.001 | 0.003 |
| TMA 1 | - | 7.06 |


3. Discussion
4. Materials and Methods
4.1. Animals and Diets
| Fatty Acid | PPC 1 |
|---|---|
| C14:0 | 6.7 |
| C16:0 | 16.0 |
| C18:0 | 0.9 |
| C20:0 | <01 |
| C22:0 | <0.1 |
| C16:1n-7 | 2.6 |
| C18:1(n-9) + (n-7) + (n-5) | 12.7 |
| C20:1(n-9) + (n-7) | 0.6 |
| C22:1(n-11) + (n-9) + (n-7) | 0.3 |
| C24:1n-9 | <0.01 |
| C18:2n-6 (LA) | 1.6 |
| C18:3n-6 | 0.1 |
| C20:2n-6 | <0.1 |
| C20:3n-6 | <0.1 |
| C20:4n-6 (AA) | 0.1 |
| C22:4n-6 | <0.1 |
| C18:3n-3 (ALA) | 3.2 |
| C18:4n-3 | 6.9 |
| C20:3n-3 | 0.1 |
| C20:4n-3 | 0.5 |
| C20:5n-3 (EPA) | 13.0 |
| C21:5n-3 | 0.5 |
| C22:5n-3 (DPA) | 0.4 |
| C22:6n-3 (DHA) | 7.9 |
| ∑SFAs | 23.6 |
| ∑MUFAs | 16.2 |
| ∑PUFAs | 35.1 |
| n-6 PUFAs | 1.8 |
| n-3 PUFAs | 32.5 |
| Sum of fatty acids | 74.9 |
| Lipids | |
| Total polar lipids | 46.9 |
| Total neutral lipids | 53.1 |
| Sum of lipids | 100 |
| Amino acid 1 | Dietary Component | |
|---|---|---|
| Casein 2 | PPC 3 | |
| Aspartic acid | 6.5 | 10.6 |
| Glutaminic acid | 20.8 | 12.7 |
| Hydroksyproline | - | <0.01 |
| Serine | 5.4 | 4.8 |
| Glycine | 1.8 | 4.6 |
| Histidine | 2.6 | 2.7 |
| Arginine | 3.6 | 6.2 |
| Threonine | 3.8 | 5.4 |
| Alanine | 2.6 | 5.1 |
| Proline | 11.7 | 4.5 |
| Tyrosine | 5.3 | 4.7 |
| Valine | 5.7 | 5.8 |
| Methionine | 2.6 | 3.3 |
| Isoleucine | 4.8 | 6.5 |
| Leucine | 8.8 | 8.8 |
| Phenylalanine | 5.0 | 5.4 |
| Lysine | 7.4 | 9.0 |
| Cysteine | 0.4 | - |
| Tryptophan | 1.2 | - |
| Methionine/glycine | 1.4 | 0.7 |
| Lysine/arginine | 2.1 | 1.5 |
4.2. Biochemical Analyses
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harris, W.S.; Dayspring, T.D.; Moran, T.J. Omega-3 fatty acids and cardiovascular disease: New developments and applications. Postgrad. Med. 2013, 125, 100–113. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Rimm, E.B. Fish intake, contaminants, and human health: Evaluating the risks and the benefits. JAMA 2006, 296, 1885–1899. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, J.; Huang, K.; Sun, Y.; Zeng, X. Purification of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide with an antihypertensive effect from loach (Misgurnus anguillicaudatus). J. Agric. Food Chem. 2012, 60, 1320–1325. [Google Scholar] [CrossRef]
- Wergedahl, H.; Gudbrandsen, O.A.; Rost, T.H.; Berge, R.K. Combination of fish oil and fish protein hydrolysate reduces the plasma cholesterol level with a concurrent increase in hepatic cholesterol level in high-fat-fed Wistar rats. Nutrition 2009, 25, 98–104. [Google Scholar] [CrossRef]
- Bjorndal, B.; Vik, R.; Brattelid, T.; Vigerust, N.F.; Burri, L.; Bohov, P.; Nygard, O.; Skorve, J.; Berge, R.K. Krill powder increases liver lipid catabolism and reduces glucose mobilization in tumor necrosis factor-alpha transgenic mice fed a high-fat diet. Metabolism 2012, 61, 1461–1472. [Google Scholar] [CrossRef]
- Berge, K.; Piscitelli, F.; Hoem, N.; Silvestri, C.; Meyer, I.; Banni, S.; di Marzo, V. Chronic treatment with krill powder reduces plasma triglyceride and anandamide levels in mildly obese men. Lipids Health Dis. 2013, 12, 78. [Google Scholar] [CrossRef]
- Collaboration, H.S. Homocysteine and risk of ischemic heart disease and stroke: A meta-analysis. JAMA 2002, 288, 2015–2022. [Google Scholar] [CrossRef]
- He, Y.; Li, Y.; Chen, Y.; Feng, L.; Nie, Z. Homocysteine level and risk of different stroke types: A meta-analysis of prospective observational studies. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1158–1165. [Google Scholar] [CrossRef]
- Clarke, R.; Bennett, D.A.; Parish, S.; Verhoef, P.; Dotsch-Klerk, M.; Lathrop, M.; Xu, P.; Nordestgaard, B.G.; Holm, H.; Hopewell, J.C.; et al. Homocysteine and coronary heart disease: Meta-analysis of MTHFR case-control studies, avoiding publication bias. PLoS Med. 2012, 9, e1001177. [Google Scholar] [CrossRef]
- Selhub, J.; Jacques, P.F.; Wilson, P.W.; Rush, D.; Rosenberg, I.H. Vitamin status and intake as primary determinants of homocysteinemia in an elderly population. JAMA 1993, 270, 2693–2698. [Google Scholar] [CrossRef]
- Ubbink, J.B.; Vermaak, W.J.; van der Merwe, A.; Becker, P.J. Vitamin B-12, vitamin B-6, and folate nutritional status in men with hyperhomocysteinemia. Am. J. Clin. Nutr. 1993, 57, 47–53. [Google Scholar]
- Marti-Carvajal, A.J.; Sola, I.; Lathyris, D.; Karakitsiou, D.E.; Simancas-Racines, D. Homocysteine-lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef]
- De La Haba, G.; Cantoni, G.L. The enzymatic synthesis of S-adenosyl-l-homocysteine from adenosine and homocysteine. J. Biol. Chem. 1959, 234, 603–608. [Google Scholar]
- Loughlin, R.E.; Elford, H.L.; Buchanan, J.M. Enzymatic synthesis of the methyl group of methionine. VII. Isolation of a cobalamin-containing transmethylase (5-methyltetrahydro-folate-homocysteine) from mammalian liver. J. Biol. Chem. 1964, 239, 2888–2895. [Google Scholar]
- Burke, G.T.; Mangum, J.H.; Brodie, J.D. Mechanism of mammalian cobalamin-dependent methionine biosynthesis. Biochemistry 1971, 10, 3079–3085. [Google Scholar]
- Borsook, H.; Dubnoff, J.W. Methionine formation by transmethylation in vitro. J. Biol. Chem. 1947, 169, 247–258. [Google Scholar]
- Finkelstein, J.D.; Harris, B.J.; Kyle, W.E. Methionine metabolism in mammals: Kinetic study of betaine-homocysteine methyltransferase. Arch. Biochem. Biophys. 1972, 153, 320–324. [Google Scholar] [CrossRef]
- Garrow, T.A. Purification, kinetic properties, and cDNA cloning of mammalian betaine-homocysteine methyltransferase. J. Biol. Chem. 1996, 271, 22831–22838. [Google Scholar]
- Lim, J.C.; You, Z.; Kim, G.; Levine, R.L. Methionine sulfoxide reductase A is a stereospecific methionine oxidase. Proc. Natl. Acad. Sci. USA 2011, 108, 10472–10477. [Google Scholar] [CrossRef]
- McCully, K.S. Chemical pathology of homocysteine. IV. Excitotoxicity, oxidative stress, endothelial dysfunction, and inflammation. Ann. Clin. Lab. Sci. 2009, 39, 219–232. [Google Scholar]
- Liao, D.; Yang, X.; Wang, H. Hyperhomocysteinemia and high-density lipoprotein metabolism in cardiovascular disease. Clin. Chem. Lab. Med. 2007, 45, 1652–1659. [Google Scholar] [CrossRef]
- Huang, T.; Zheng, J.; Chen, Y.; Yang, B.; Wahlqvist, M.L.; Li, D. High consumption of omega-3 polyunsaturated fatty acids decrease plasma homocysteine: A meta-analysis of randomized, placebo-controlled trials. Nutrition 2011, 27, 863–867. [Google Scholar] [CrossRef]
- Gudbrandsen, O.A.; Wergedahl, H.; Liaset, B.; Espe, M.; Berge, R.K. Dietary proteins with high isoflavone content or low methionine-glycine and lysine-arginine ratios are hypocholesterolaemic and lower the plasma homocysteine level in male Zucker fa/fa rats. Br. J. Nutr. 2005, 94, 321–330. [Google Scholar] [CrossRef]
- Bremer, J. Carnitine—Metabolism and functions. Physiol. Rev. 1983, 63, 1420–1480. [Google Scholar]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Rebouche, C.J.; Seim, H. Carnitine metabolism and its regulation in microorganisms and mammals. Annu. Rev. Nutr. 1998, 18, 39–61. [Google Scholar] [CrossRef]
- Troseid, M.; Ueland, T.; Hov, J.R.; Svardal, A.; Gregersen, I.; Dahl, C.P.; Aakhus, S.; Gude, E.; Bjorndal, B.; Halvorsen, B.; et al. Microbiota-dependent metabolite trimethylamine-N-oxide is associated with disease severity and survival of patients with chronic heart failure. J. Intern. Med. 2014, 277, 717–726. [Google Scholar] [CrossRef]
- Yancey, P.H.; Fyfe-Johnson, A.L.; Kelly, R.H.; Walker, V.P.; Aunon, M.T. Trimethylamine oxide counteracts effects of hydrostatic pressure on proteins of deep-sea teleosts. J. Exp. Zool. 2001, 289, 172–176. [Google Scholar] [CrossRef]
- Someo, G.N. From dogfish to dogs: Trimethylamines protect proteins from urea. News Physiol. Sci. 1986, 1, 9–12. [Google Scholar]
- Raymond, J.A. Seasonal variations of trimethylamine oxide and urea in the blood of a cold-adapted marine teleost, the rainbow smelt. Fish Physiol. Biochem. 1994, 13, 13–22. [Google Scholar] [CrossRef]
- Yancey, P.H.; Siebenaller, J.F. Trimethylamine oxide stabilizes teleost and mammalian lactate dehydrogenases against inactivation by hydrostatic pressure and trypsinolysis. J. Exp. Biol. 1999, 202, 3597–3603. [Google Scholar]
- Cicero, A.F.G.; Colletti, A. Krill oil: Evidence of a new source of polyunsaturated fatty acids with high bioavailibility. J. Clin. Lipidol. 2015, 10, 1–4. [Google Scholar] [CrossRef]
- Ramsvik, M.S.; Bjorndal, B.; Bruheim, I.; Bohov, P.; Berge, R.K. A Phospholipid-Protein Complex from Krill with Antioxidative and Immunomodulating Properties Reduced Plasma Triacylglycerol and Hepatic Lipogenesis in Rats. Mar. Drugs 2015, 13, 4375–4397. [Google Scholar] [CrossRef]
- Klerk, M.; Verhoef, P.; Clarke, R.; Blom, H.J.; Kok, F.J.; Schouten, E.G. MTHFR 677C-->T polymorphism and risk of coronary heart disease: A meta-analysis. JAMA 2002, 288, 2023–2031. [Google Scholar] [CrossRef]
- Refsum, H.; Ueland, P.M.; Nygard, O.; Vollset, S.E. Homocysteine and cardiovascular disease. Annu. Rev. Med. 1998, 49, 31–62. [Google Scholar] [CrossRef]
- Wald, D.S.; Law, M.; Morris, J.K. Homocysteine and cardiovascular disease: Evidence on causality from a meta-analysis. BMJ 2002, 325, 1202. [Google Scholar] [CrossRef]
- Lee, J.E.; Jacques, P.F.; Dougherty, L.; Selhub, J.; Giovannucci, E.; Zeisel, S.H.; Cho, E. Are dietary choline and betaine intakes determinants of total homocysteine concentration? Am. J. Clin. Nutr. 2010, 91, 1303–1310. [Google Scholar] [CrossRef]
- Chiuve, S.E.; Giovannucci, E.L.; Hankinson, S.E.; Zeisel, S.H.; Dougherty, L.W.; Willett, W.C.; Rimm, E.B. The association between betaine and choline intakes and the plasma concentrations of homocysteine in women. Am. J. Clin. Nutr. 2007, 86, 1073–1081. [Google Scholar]
- Rajaie, S.; Esmaillzadeh, A. Dietary choline and betaine intakes and risk of cardiovascular diseases: Review of epidemiological evidence. ARYA Atheroscler. 2011, 7, 78–86. [Google Scholar]
- Bidulescu, A.; Chambless, L.E.; Siega-Riz, A.M.; Zeisel, S.H.; Heiss, G. Usual choline and betaine dietary intake and incident coronary heart disease: The Atherosclerosis Risk in Communities (ARIC) study. BMC Cardiovasc. Disord. 2007, 7, 20. [Google Scholar] [CrossRef]
- Magnusson, M.; Wang, T.J.; Clish, C.; Engstrom, G.; Nilsson, P.; Gerszten, R.E.; Melander, O. Dimethylglycine Deficiency and the Development of Diabetes. Diabetes 2015, 64, 3010–3016. [Google Scholar] [CrossRef]
- Huang, T.; Wahlqvist, M.L.; Li, D. Docosahexaenoic acid decreases plasma homocysteine via regulating enzyme activity and mRNA expression involved in methionine metabolism. Nutrition 2010, 26, 112–119. [Google Scholar] [CrossRef]
- Zhang, A.Q.; Mitchell, S.C.; Smith, R.L. Dietary precursors of trimethylamine in man: A pilot study. Food Chem. Toxicol. 1999, 37, 515–520. [Google Scholar] [CrossRef]
- Goldstein, L.; Funkhouser, D. Biosynthesis of trimethylamine oxide in the nurse shark, Ginglymostoma cirratum. Comp. Biochem. Physiol. A Comp. Physiol. 1972, 42, 51–57. [Google Scholar] [CrossRef]
- Anfuso, C.D.; Sipione, S.; Lupo, G.; Alberghina, M. Evolutionary comparison of enzyme activities of phosphatidylcholine metabolism in the nervous system of an invertebrate (Loligo pealei), lower vertebrate (Mustelus canis) and the rat. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1995, 112, 493–501. [Google Scholar] [CrossRef]
- Bjorndal, B.; Strand, E.; Gjerde, J.; Bohov, P.; Svardal, A.; Diehl, B.W.; Innis, S.M.; Berger, A.; Berge, R.K. Phospholipids from herring roe improve plasma lipids and glucose tolerance in healthy, young adults. Lipids Health Dis. 2014, 13, 82. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, W.H.; Buffa, J.A.; Fu, X.; Britt, E.B.; Koeth, R.A.; Levison, B.S.; Fan, Y.; Wu, Y.; Hazen, S.L. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur. Heart J. 2014, 35, 904–910. [Google Scholar] [CrossRef]
- Lever, M.; George, P.M.; Slow, S.; Bellamy, D.; Young, J.M.; Ho, M.; McEntyre, C.J.; Elmslie, J.L.; Atkinson, W.; Molyneux, S.L.; et al. Betaine and Trimethylamine-N-Oxide as Predictors of Cardiovascular Outcomes Show Different Patterns in Diabetes Mellitus: An Observational Study. PLoS ONE 2014, 9, e114969. [Google Scholar] [CrossRef]
- Ussher, J.R.; Lopaschuk, G.D.; Arduini, A. Gut microbiota metabolism of l-carnitine and cardiovascular risk. Atherosclerosis 2013, 231, 456–461. [Google Scholar] [CrossRef]
- Vigerust, N.F.; Bjorndal, B.; Bohov, P.; Brattelid, T.; Svardal, A.; Berge, R.K. Krill oil versus fish oil in modulation of inflammation and lipid metabolism in mice transgenic for TNF-alpha. Eur. J. Nutr. 2012, 52, 1315–1325. [Google Scholar] [CrossRef]
- Hongu, N.; Sachan, D.S. Carnitine and choline supplementation with exercise alter carnitine profiles, biochemical markers of fat metabolism and serum leptin concentration in healthy women. J. Nutr. 2003, 133, 84–89. [Google Scholar]
- Pekkinen, J.; Olli, K.; Huotari, A.; Tiihonen, K.; Keski-Rahkonen, P.; Lehtonen, M.; Auriola, S.; Kolehmainen, M.; Mykkanen, H.; Poutanen, K.; et al. Betaine supplementation causes increase in carnitine metabolites in the muscle and liver of mice fed a high-fat diet as studied by nontargeted LC-MS metabolomics approach. Mol. Nutr. Food Res. 2013, 57, 1959–1968. [Google Scholar] [CrossRef]
- Panchal, S.K.; Poudyal, H.; Ward, L.C.; Waanders, J.; Brown, L. Modulation of tissue fatty acids by l-carnitine attenuates metabolic syndrome in diet-induced obese rats. Food Funct. 2015, 6, 2496–2506. [Google Scholar] [CrossRef]
- Shang, R.; Sun, Z.; Li, H. Effective dosing of l-carnitine in the secondary prevention of cardiovascular disease: A systematic review and meta-analysis. BMC Cardiovasc. Disord. 2014, 14, 88. [Google Scholar] [CrossRef]
- Ueland, P.M.; Midttun, O.; Windelberg, A.; Svardal, A.; Skalevik, R.; Hustad, S. Quantitative profiling of folate and one-carbon metabolism in large-scale epidemiological studies by mass spectrometry. Clin. Chem. Lab. Med. 2007, 45, 1737–1745. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bjørndal, B.; Ramsvik, M.S.; Lindquist, C.; Nordrehaug, J.E.; Bruheim, I.; Svardal, A.; Nygård, O.; Berge, R.K. A Phospholipid-Protein Complex from Antarctic Krill Reduced Plasma Homocysteine Levels and Increased Plasma Trimethylamine-N-Oxide (TMAO) and Carnitine Levels in Male Wistar Rats. Mar. Drugs 2015, 13, 5706-5721. https://doi.org/10.3390/md13095706
Bjørndal B, Ramsvik MS, Lindquist C, Nordrehaug JE, Bruheim I, Svardal A, Nygård O, Berge RK. A Phospholipid-Protein Complex from Antarctic Krill Reduced Plasma Homocysteine Levels and Increased Plasma Trimethylamine-N-Oxide (TMAO) and Carnitine Levels in Male Wistar Rats. Marine Drugs. 2015; 13(9):5706-5721. https://doi.org/10.3390/md13095706
Chicago/Turabian StyleBjørndal, Bodil, Marie S. Ramsvik, Carine Lindquist, Jan E. Nordrehaug, Inge Bruheim, Asbjørn Svardal, Ottar Nygård, and Rolf K. Berge. 2015. "A Phospholipid-Protein Complex from Antarctic Krill Reduced Plasma Homocysteine Levels and Increased Plasma Trimethylamine-N-Oxide (TMAO) and Carnitine Levels in Male Wistar Rats" Marine Drugs 13, no. 9: 5706-5721. https://doi.org/10.3390/md13095706
APA StyleBjørndal, B., Ramsvik, M. S., Lindquist, C., Nordrehaug, J. E., Bruheim, I., Svardal, A., Nygård, O., & Berge, R. K. (2015). A Phospholipid-Protein Complex from Antarctic Krill Reduced Plasma Homocysteine Levels and Increased Plasma Trimethylamine-N-Oxide (TMAO) and Carnitine Levels in Male Wistar Rats. Marine Drugs, 13(9), 5706-5721. https://doi.org/10.3390/md13095706