
Excavatolide B Attenuates Rheumatoid Arthritis through the Inhibition of Osteoclastogenesis

 ,
, 
 and
and 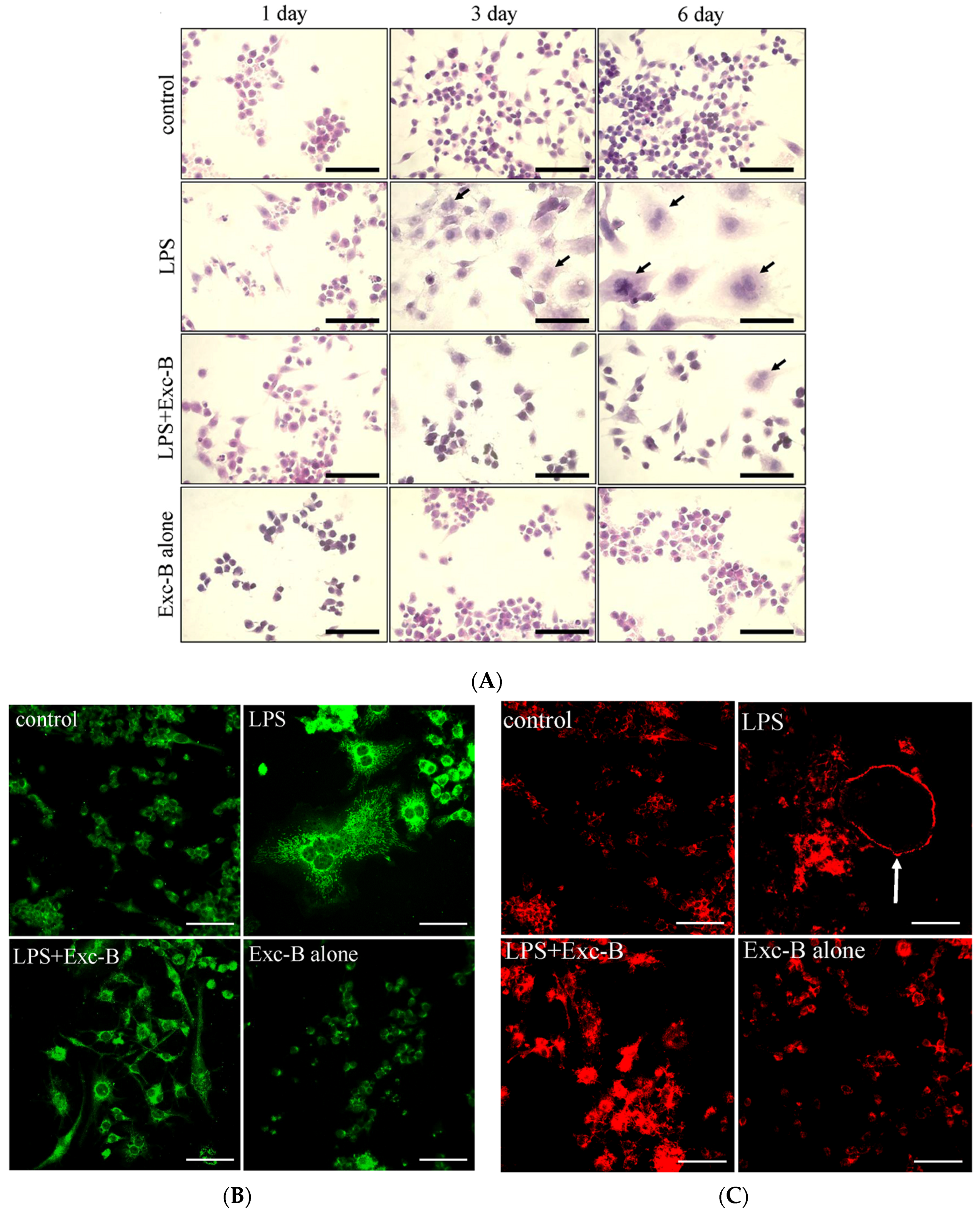{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
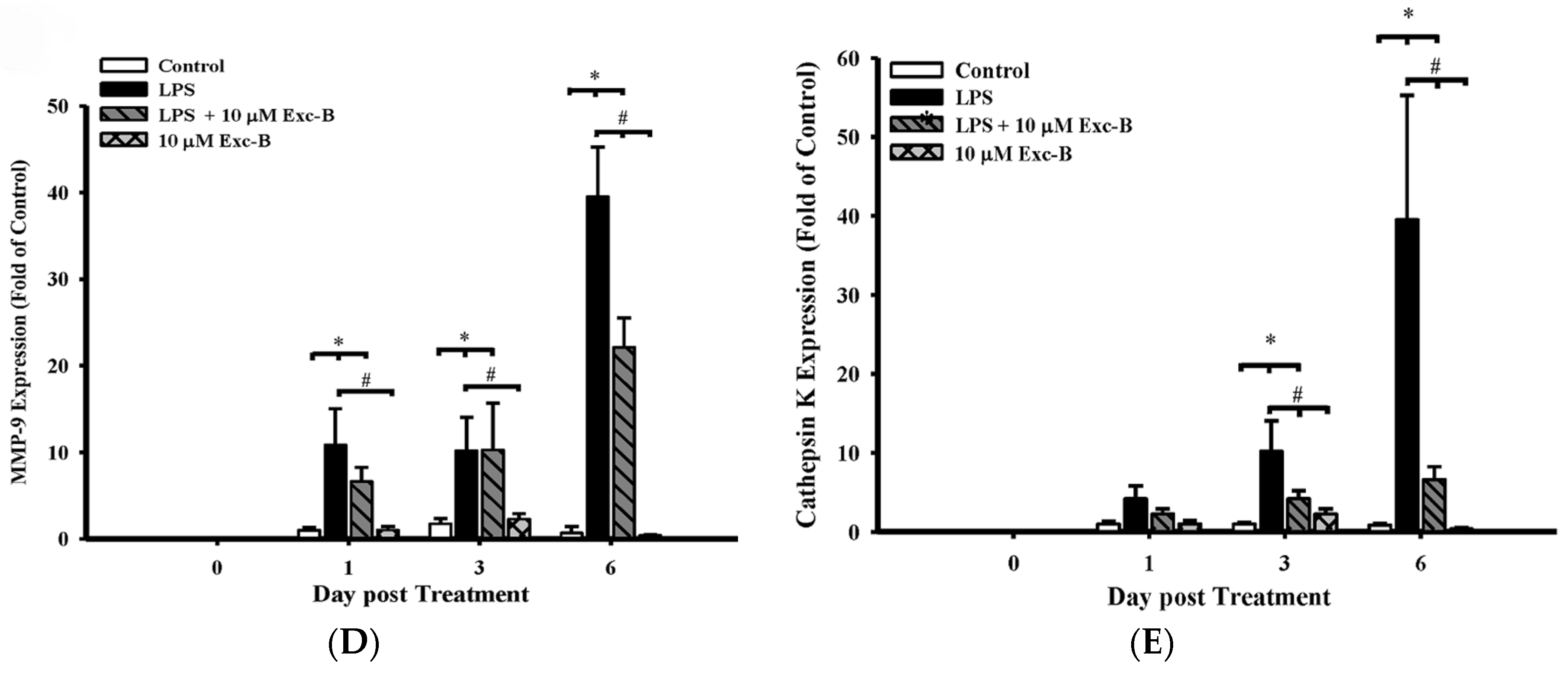
2.1. Exc-B Inhibits LPS-Induced Osteoclast-Like Cell Formation
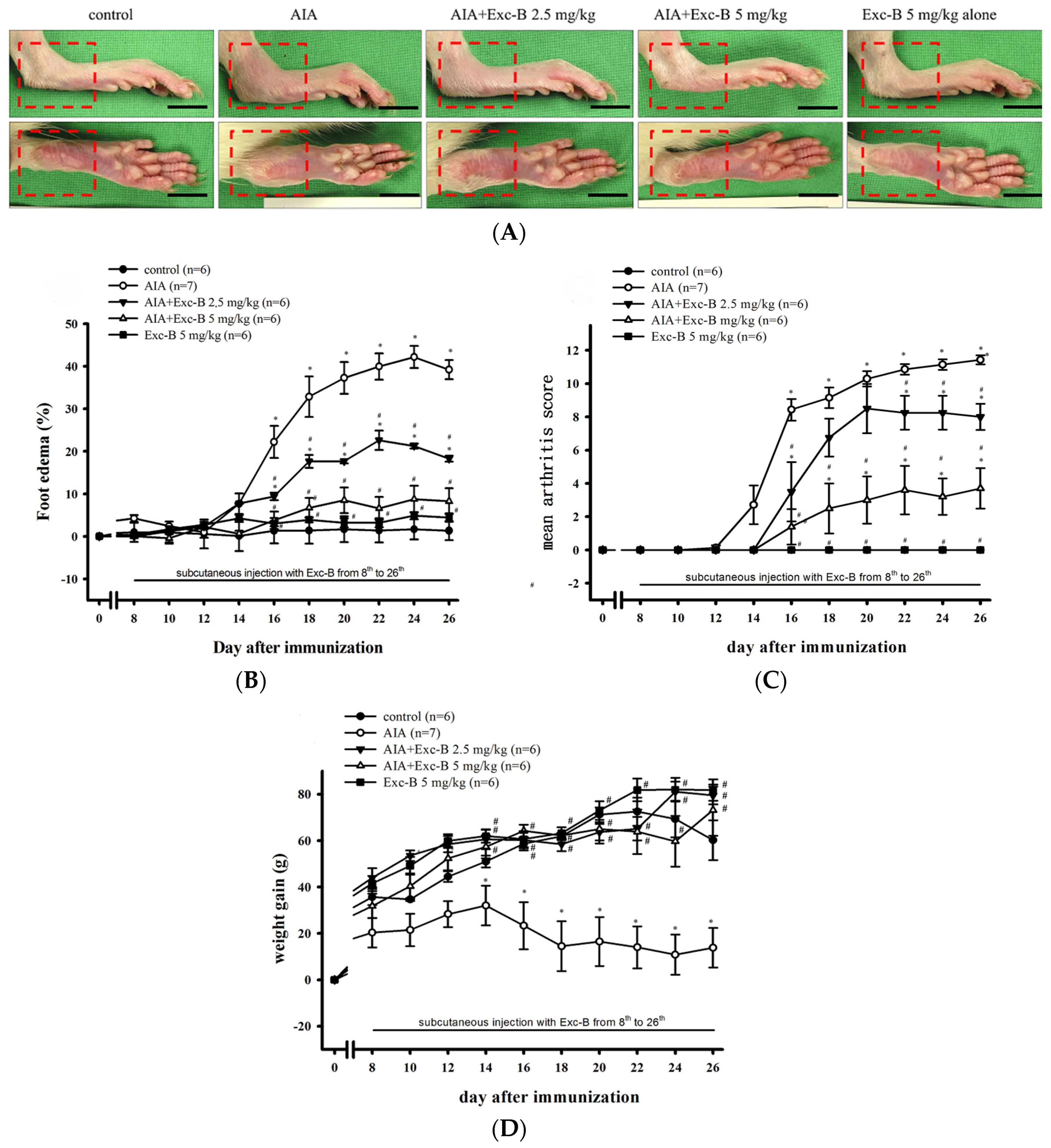
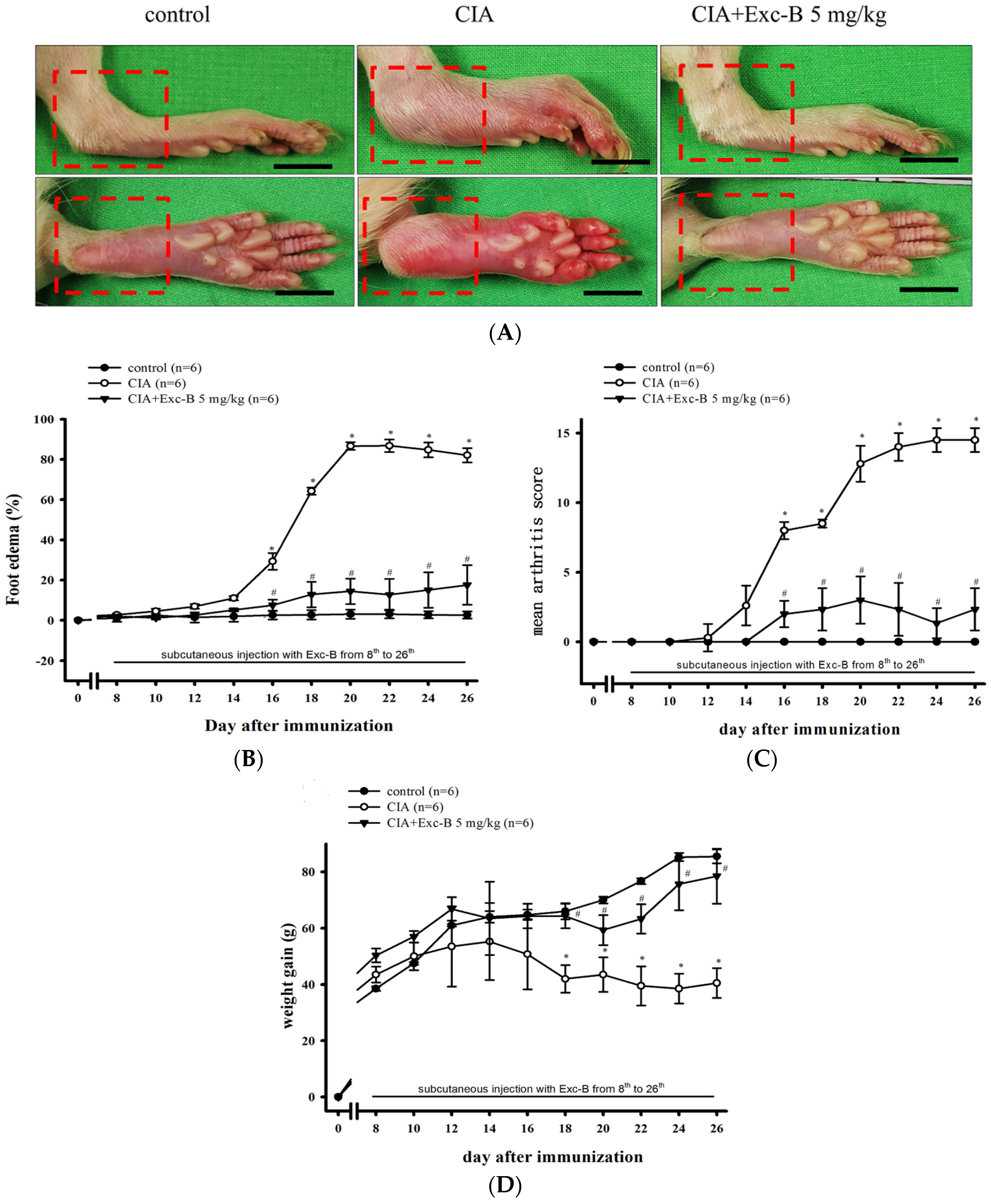
2.2. Effect of Exc-B on the Clinical Features of RA
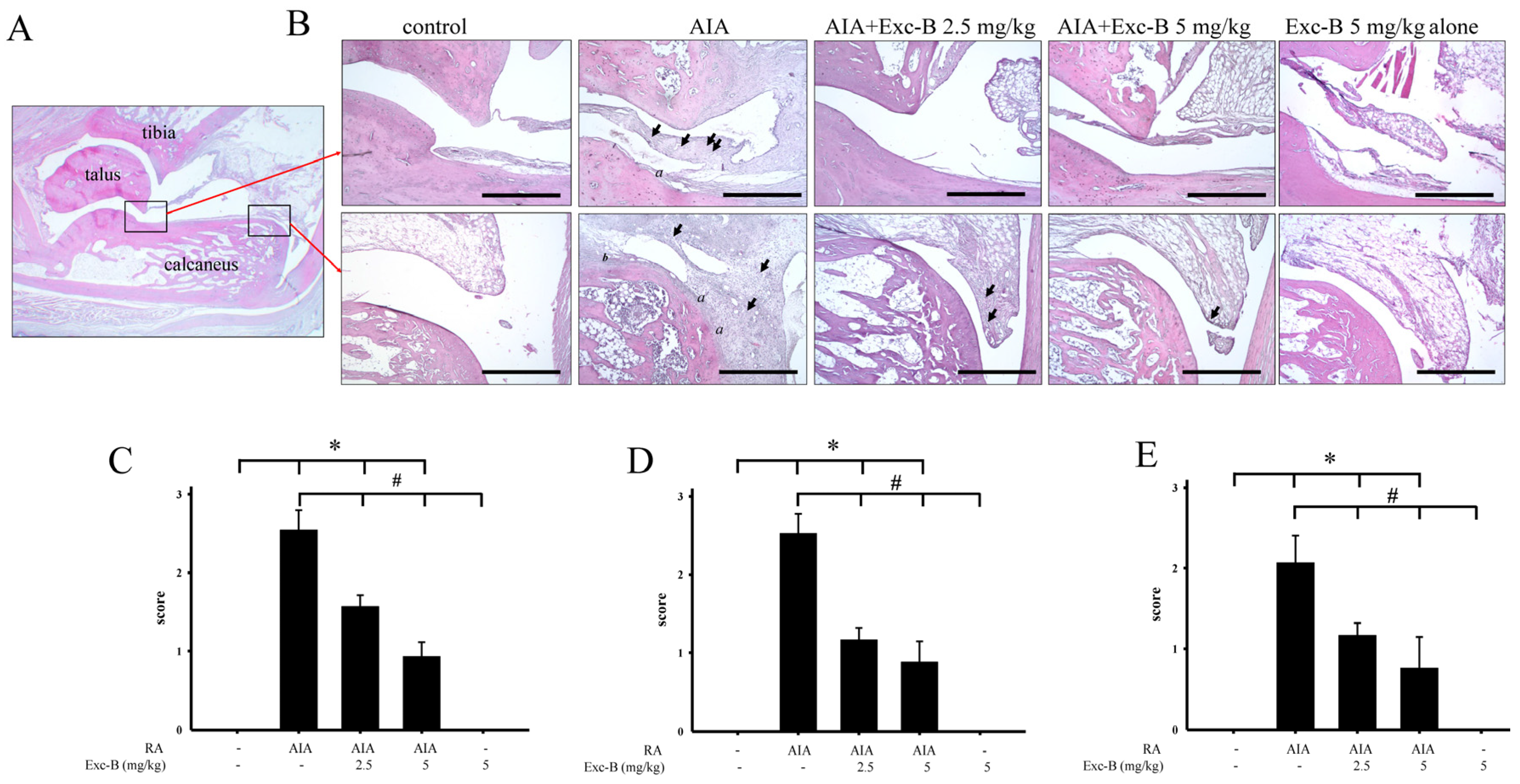
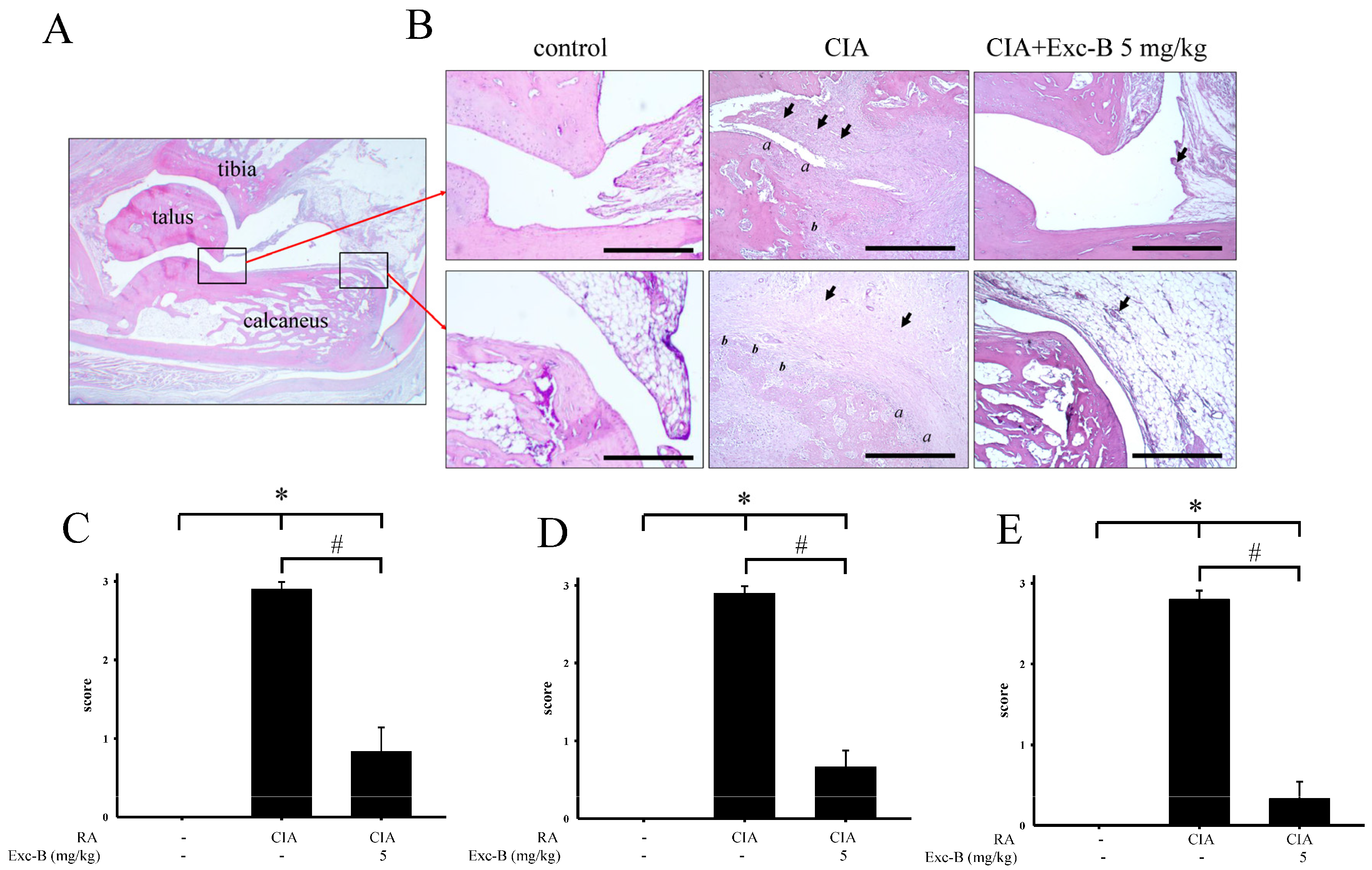
2.3. Effect of Exc-B on Histological Features of RA
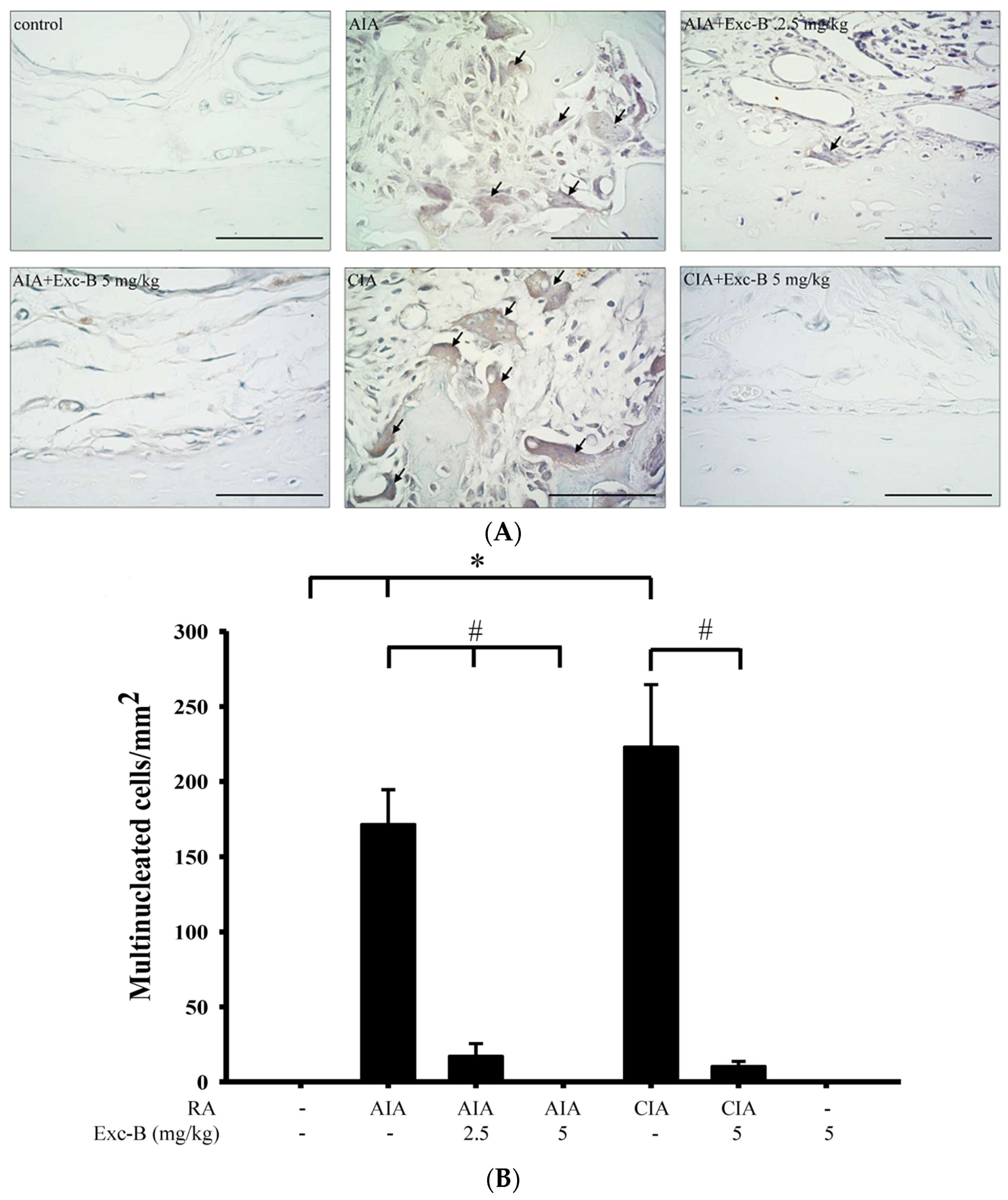
2.4. Effect of Exc-B on TRAP-Positive Multinucleated Cell Formation In Vivo
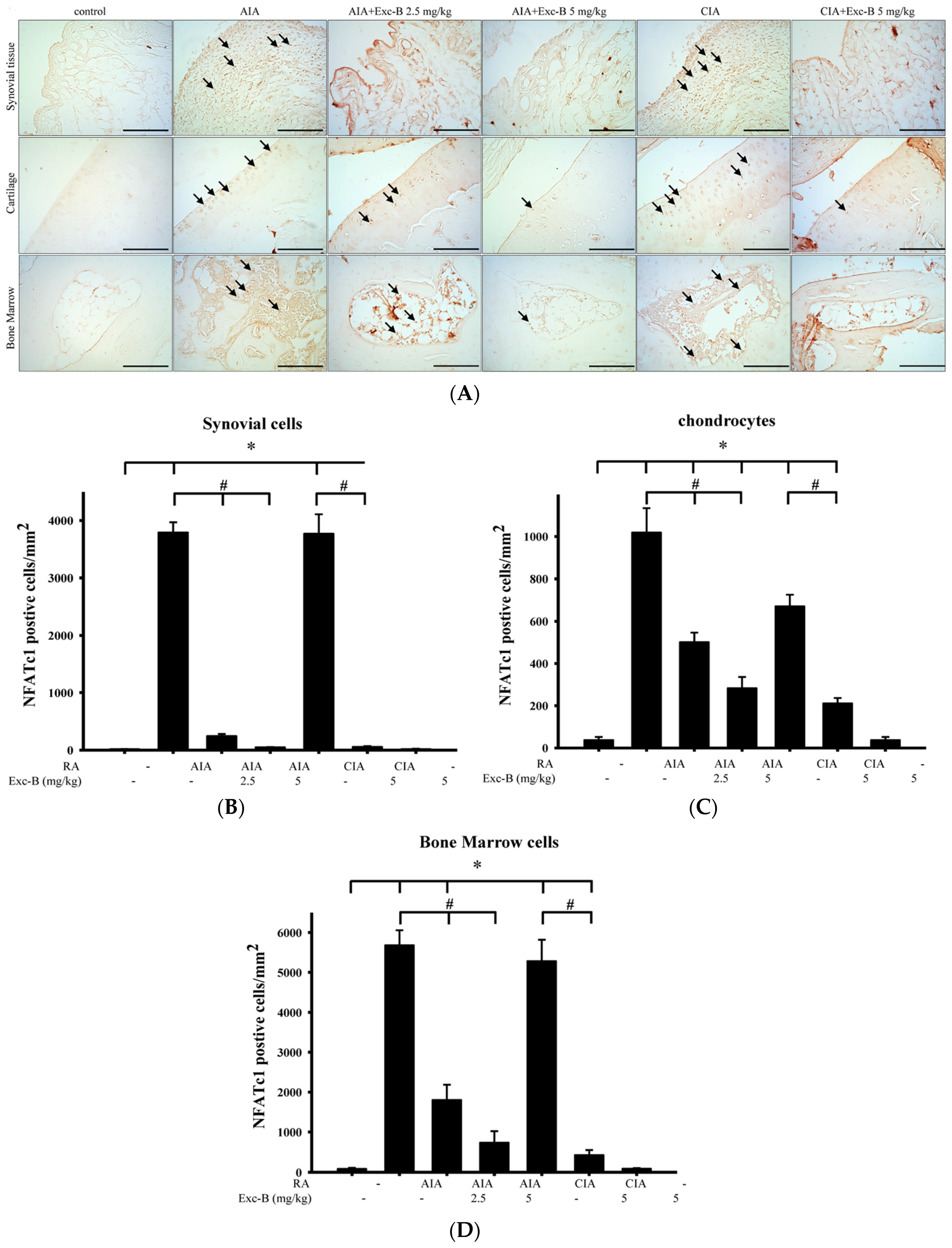
2.5. Effect of Exc-B on NFATc1 in Arthritic Rats
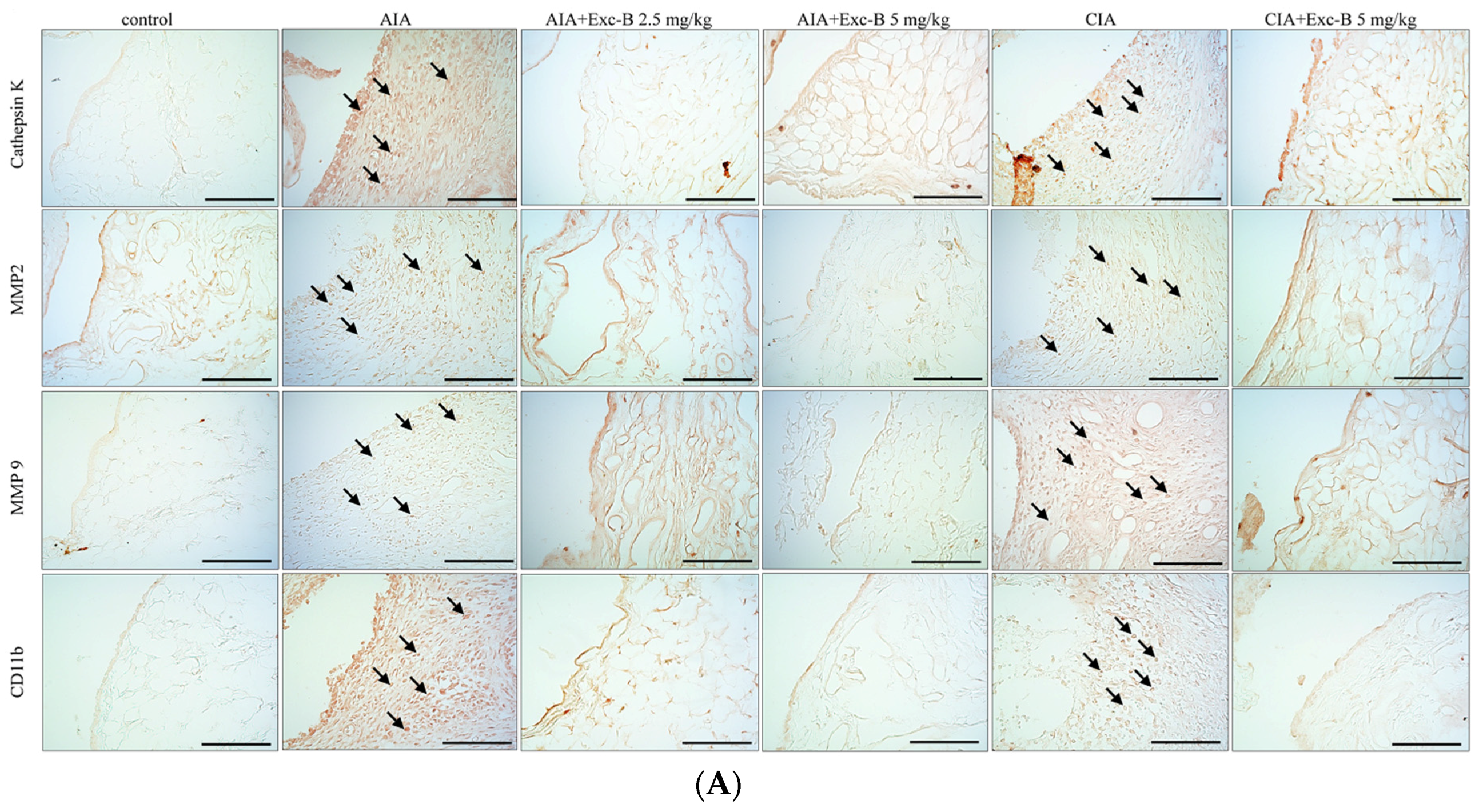
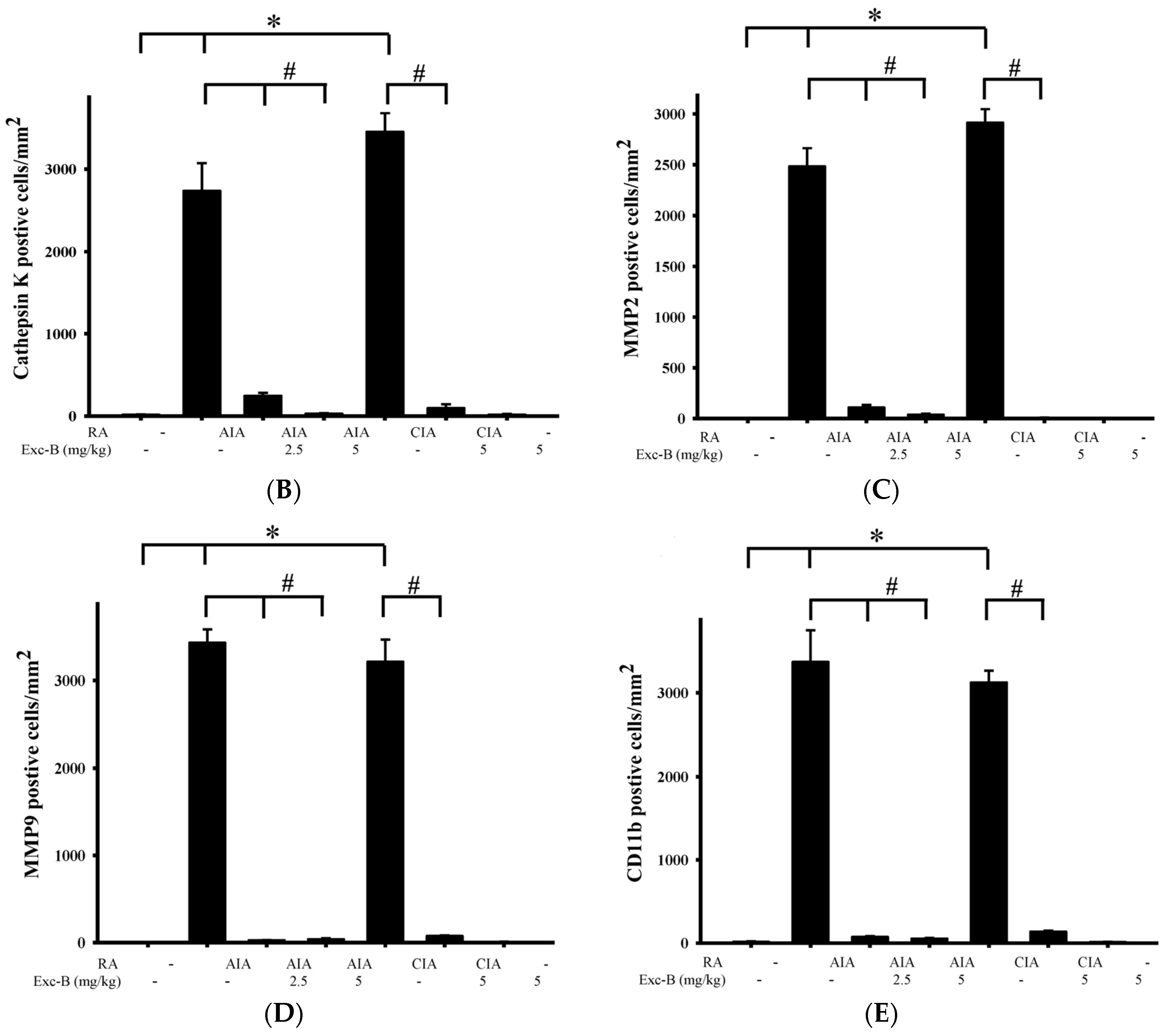
2.6. Effect of Exc-B on Cathepsin K, MMP-2, MMP-9, and CD11b in Synovial Tissue
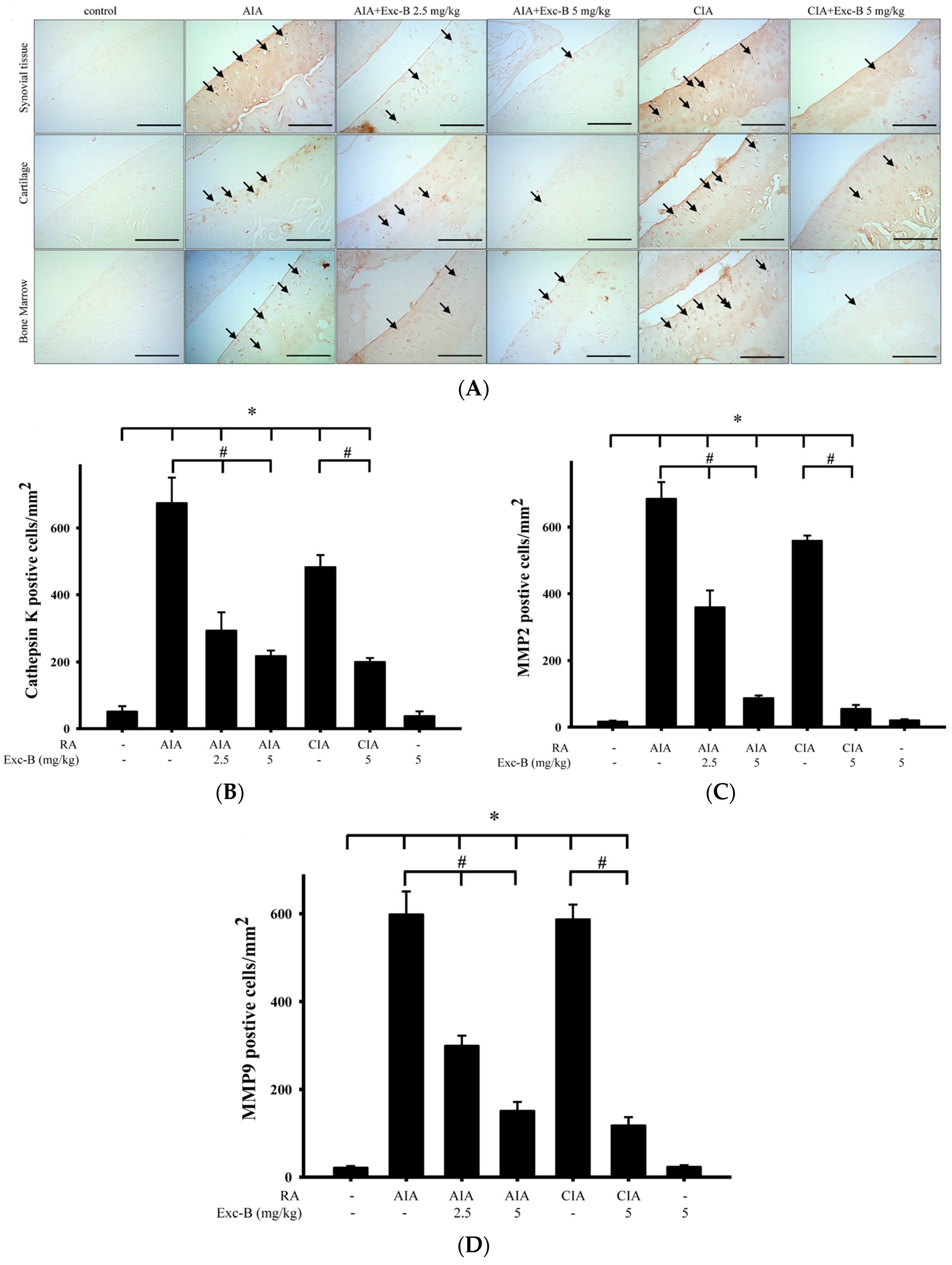
2.7. Effect of Exc-B on Cathepsin K, MMP-2, and MMP-9 in Cartilage
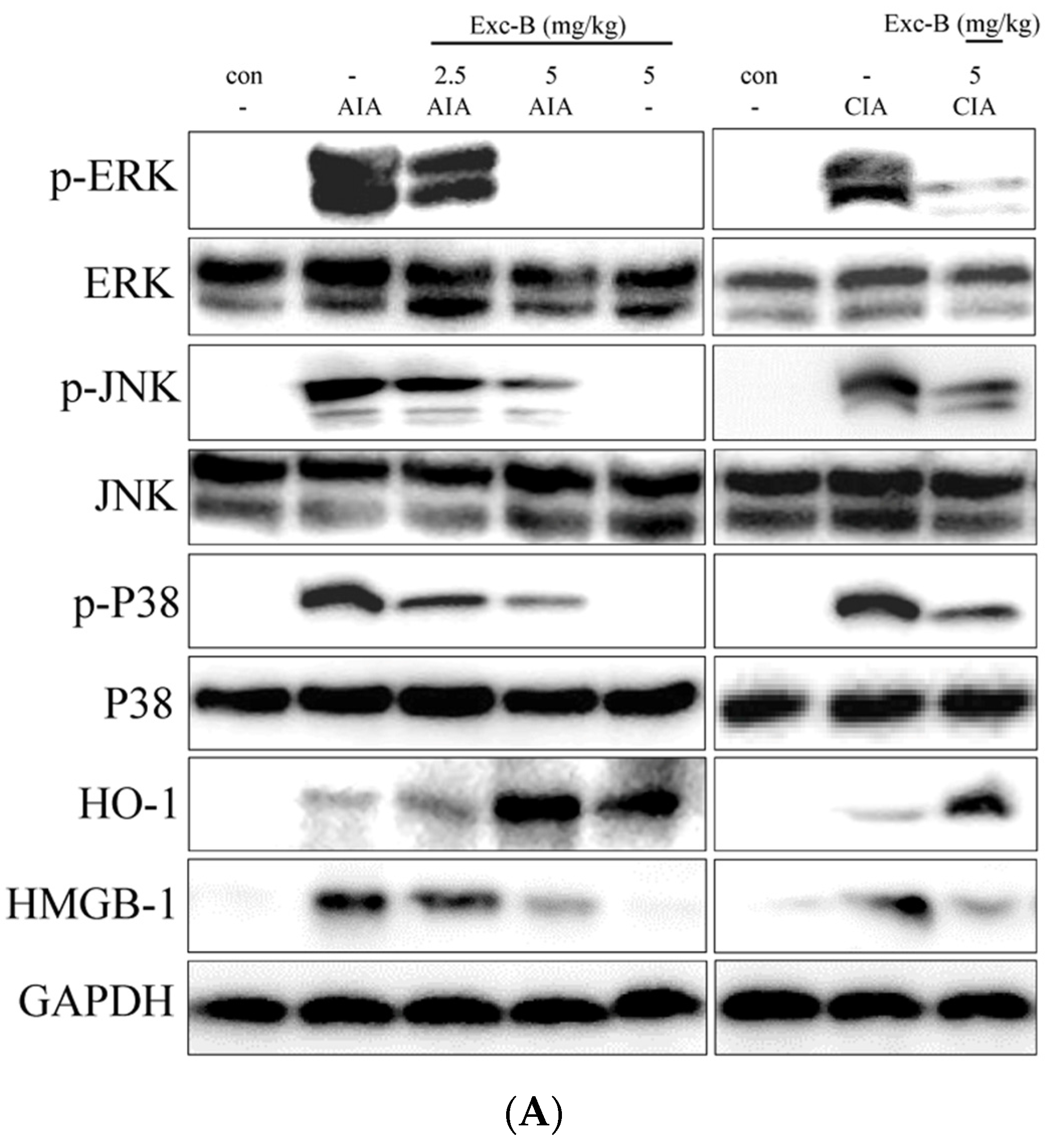
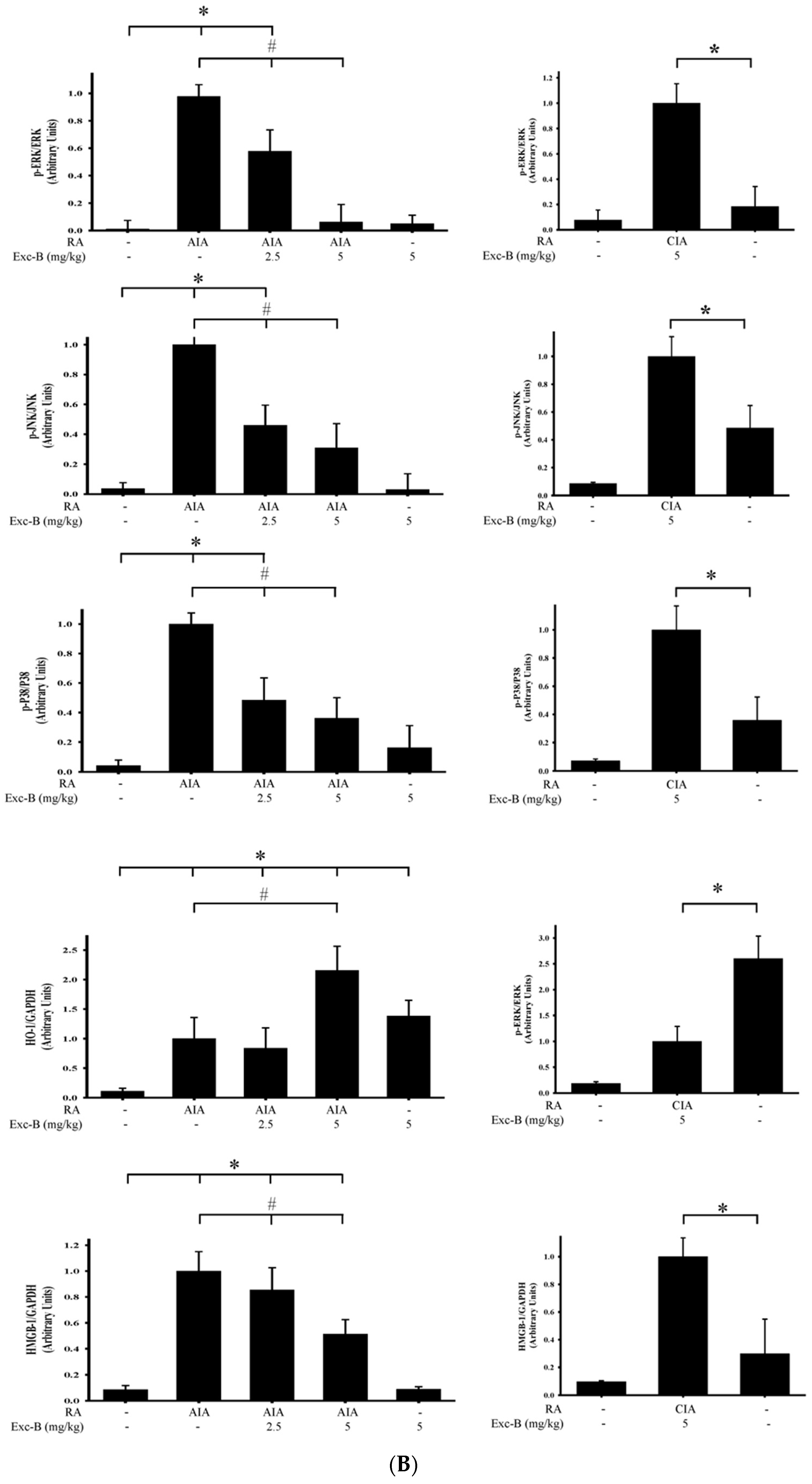
2.8. Effect of Exc-B on MAPK, HO-1, and HMGB-1 Protein Expression in Synovial Tissue
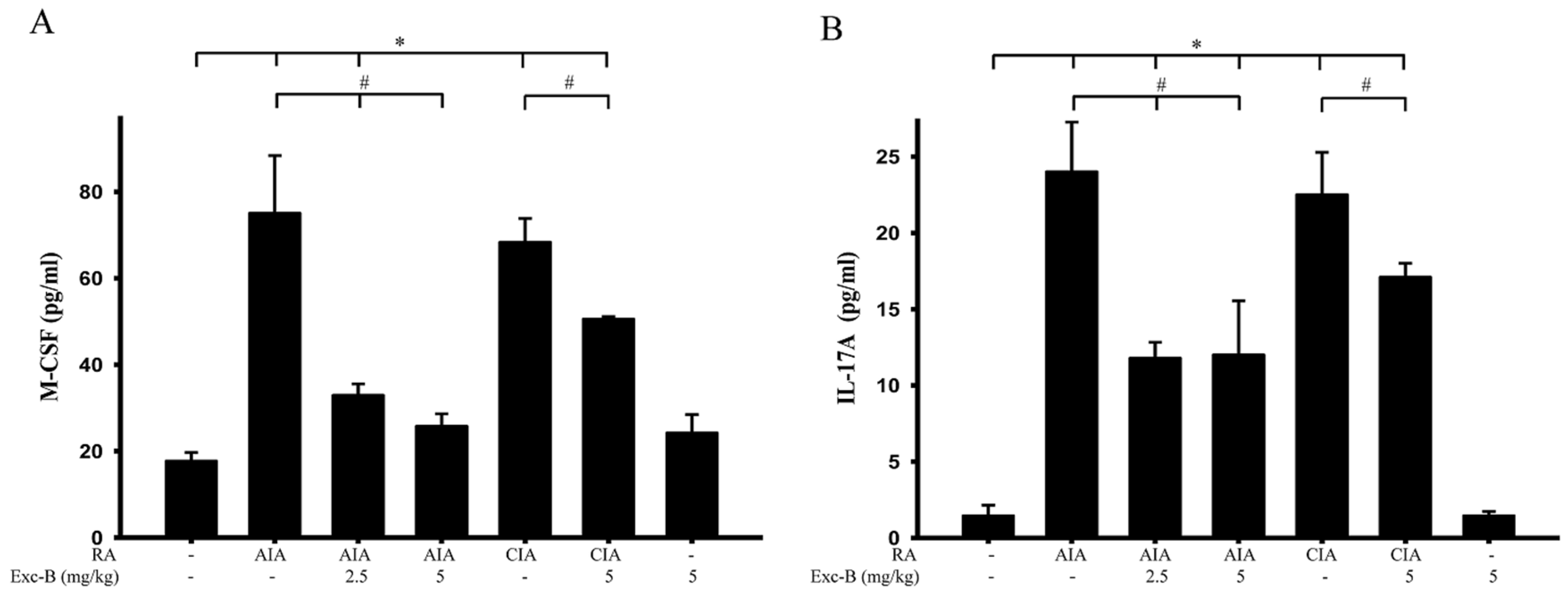
2.9. Effect of Exc-B on M-CSF and IL-17A of Blood Serum
3. Discussion
3.1. Summary
3.2. Anti-Inflammatory Effect of Exc-B In Vitro and In Vivo
3.3. Anti-Osteoclast Activity of Exc-B
3.4. The Possible Therapeutic Mechanism of Exc-B in RA
3.5. Coral Aquaculture for Anti-Inflammatory and Anti-Arthritic Agents
4. Materials and Methods
4.1. Extraction of Exc-B
4.2. LPS-Induced Osteoclast-Like Cell Model
4.3. Real-Time PCR Analysis for Cathepsin K and MMP-9 mRNA
4.4. Immunocytochemistry
4.5. Animals
4.6. Adjuvant-Induced Arthritis
4.7. Collagen-Induced Arthritis
4.8. Experimental Design
4.9. Histopathological Examination of the Ankle Joint
4.10. Immunohistochemical Staining
4.11. Preparation of Synovial Tissue for Western Blot Analysis
4.12. Measurements of Serum IL-17A and M-CSF Levels
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Silman, A.J.; Pearson, J.E. Epidemiology and genetics of rheumatoid arthritis. Arthritis Res. 2002, 4 (Suppl. 3), S265–S272. [Google Scholar] [CrossRef] [PubMed]
- Koenders, M.I.; van den Berg, W.B. Novel therapeutic targets in rheumatoid arthritis. Trends Pharmacol. Sci. 2015, 36, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Kimoto, A.; Kobayashi, S.; Yoshino, T.; Miyata, K.; Sasamata, M. Effect of celecoxib, a cyclooxygenase-2 inhibitor, on the pathophysiology of adjuvant arthritis in rat. Eur. J. Pharmacol. 2005, 513, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.L.; Smith, C.; Kingsley, G. Joint damage and disability in rheumatoid arthritis: An updated systematic review. Clin. Exp. Rheumatol. 2003, 21, S20–S27. [Google Scholar] [PubMed]
- Shiozawa, S.; Tsumiyama, K.; Yoshida, K.; Hashiramoto, A. Pathogenesis of joint destruction in rheumatoid arthritis. Arch. Immunol. Ther. Exp. 2011, 59, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Paleolog, E.M. Angiogenesis in rheumatoid arthritis. Arthritis Res. 2002, 4 (Suppl. 3), S81–S90. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.C.; Distler, O.; Tarner, I.; Gay, R.E.; Gay, S.; Pap, T. Synovial fibroblasts: Key players in rheumatoid arthritis. Rheumatology 2006, 45, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Pettit, A.R.; Walsh, N.C.; Manning, C.; Goldring, S.R.; Gravallese, E.M. Rankl protein is expressed at the pannus-bone interface at sites of articular bone erosion in rheumatoid arthritis. Rheumatology 2006, 45, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.M.; Axmann, R.; Zwerina, J.; Polzer, K.; Guckel, E.; Skapenko, A.; Schulze-Koops, H.; Horwood, N.; Cope, A.; Schett, G. Treg cells suppress osteoclast formation: A new link between the immune system and bone. Arthritis Rheumatol. 2007, 56, 4104–4112. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Nakahama, K.; Sato, T.; Tuchiya, T.; Asakawa, Y.; Maemura, T.; Tanaka, M.; Morita, M.; Morita, I. The role of MAC-1 (CD11b/CD18) in osteoclast differentiation induced by receptor activator of nuclear factor-kappaB ligand. FEBS Lett. 2008, 582, 3243–3248. [Google Scholar] [CrossRef] [PubMed]
- Long, C.L.; Humphrey, M.B. Osteoimmunology: The expanding role of immunoreceptors in osteoclasts and bone remodeling. Bonekey Rep. 2012, 1. [Google Scholar] [CrossRef] [PubMed]
- McFarland, H.I.; Nahill, S.R.; Maciaszek, J.W.; Welsh, R.M. CD11b (MAC-1): A marker for CD8+ cytotoxic t cell activation and memory in virus infection. J. Immunol. 1992, 149, 1326–1333. [Google Scholar] [PubMed]
- Roeleveld, D.M.; Koenders, M.I. The role of the Th17 cytokines IL-17 and IL-22 in rheumatoid arthritis pathogenesis and developments in cytokine immunotherapy. Cytokine 2015, 74, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Kellner, H. Targeting interleukin-17 in patients with active rheumatoid arthritis: Rationale and clinical potential. Ther. Adv. Musculoskelet. Dis. 2013, 5, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Lubberts, E. The IL-23-IL-17 axis in inflammatory arthritis. Nat. Rev. Rheumatol. 2015, 11, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, W.B.; Miossec, P. IL-17 as a future therapeutic target for rheumatoid arthritis. Nat. Rev. Rheumatol. 2009, 5, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. The role of nfat in osteoclast formation. Ann. N. Y. Acad. Sci. 2007, 1116, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Sitara, D.; Aliprantis, A.O. Transcriptional regulation of bone and joint remodeling by nfat. Immunol. Rev. 2010, 233, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Jean, Y.H.; Chen, W.F.; Duh, C.Y.; Huang, S.Y.; Hsu, C.H.; Lin, C.S.; Sung, C.S.; Chen, I.M.; Wen, Z.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory and analgesic effects of the natural marine compound lemnalol from formosan soft coral lemnalia cervicorni. Eur. J. Pharmacol. 2008, 578, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Hamade, E.; Habib, A.; Hachem, A.; Hussein, A.H.; Abbas, M.; Hirz, T.; Al Masri, M.; Faour, W.H. Biological and anti-inflammatory evaluation of two thiazole compounds in raw cell line: Potential cyclooxygenase-2 specific inhibitors. Med. Chem. 2012, 8, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Hassan, F.; Tumurkhuu, G.; Dagvadorj, J.; Koide, N.; Naiki, Y.; Mori, I.; Yoshida, T.; Yokochi, T. Bacterial lipopolysaccharide induces osteoclast formation in raw 264.7 macrophage cells. Biochem. Biophys. Res. Commun. 2007, 360, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Turull, N.; Queralt, J. Effect of the COX-2 selective inhibitor L-745,337 on inflammation and organ prostaglandin E2 (PGE2) levels in adjuvant arthritic rats. Inflammation 2000, 24, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.X.; Chen, J.Y.; Kou, J.Q.; Xu, Y.L.; Wang, S.Z.; Zhu, Q.; Yang, L.; Qin, Z.H. Suppression of inflammation and arthritis by orally administrated cardiotoxin from Naja naja atra. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Subramaneyaan, M.; Arora, V.K.; Banerjee, B.D.; Ahmed, R.S. Effect of withania somnifera (ashwagandha) root extract on amelioration of oxidative stress and autoantibodies production in collagen-induced arthritic rats. J. Complement. Integr. Med. 2015, 12, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Netsu, T.; Kondo, N.; Arai, K.; Ogose, A.; Endo, N. Osteoconductive action of alendronate after implantation of beta tricalcium phosphate in rat adjuvant-induced arthritis. J. Bone Miner. Metab. 2012, 30, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Marchesan, J.T.; Gerow, E.A.; Schaff, R.; Taut, A.D.; Shin, S.Y.; Sugai, J.; Brand, D.; Burberry, A.; Jorns, J.; Lundy, S.K.; et al. Porphyromonas gingivalis oral infection exacerbates the development and severity of collagen-induced arthritis. Arthritis Res. Ther. 2013, 15, R186. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Hogan, J.K.; Shlyakhter, D.; Oh, L.; Salituro, F.G.; Farmer, L.; Hoock, T.C. VX-509 (decernotinib) is a potent and selective janus kinase 3 inhibitor that attenuates inflammation in animal models of autoimmune disease. J. Pharmacol. Exp. Ther. 2015, 353, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.; Peixe, L.; Gomes, N.C.; Calado, R. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Calado, R.; Sheridan, C.; Alimonti, A.; Osinga, R. Coral aquaculture to support drug discovery. Trends Biotechnol. 2013, 31, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.C.; Sung, P.J.; Duh, C.Y.; Chen, B.W.; Sheu, J.H.; Yang, N.S. Anti-inflammatory activities of natural products isolated from soft corals of taiwan between 2008 and 2012. Mar. Drugs 2013, 11, 4083–4126. [Google Scholar] [CrossRef] [PubMed]
- Liaw, C.C.; Shen, Y.C.; Lin, Y.S.; Hwang, T.L.; Kuo, Y.H.; Khalil, A.T. Frajunolides E-K, briarane diterpenes from Junceella fragilis. J. Nat. Prod. 2008, 71, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Liaw, C.C.; Lin, Y.C.; Lin, Y.S.; Chen, C.H.; Hwang, T.L.; Shen, Y.C. Four new briarane diterpenoids from taiwanese gorgonian junceella fragilis. Mar. Drugs 2013, 11, 2042–2053. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Sung, P.J.; Cheng, M.C.; Liu, H.Y.; Fang, L.S.; Duh, C.Y.; Chiang, M.Y. Novel cytotoxic diterpenes, excavatolides A-E, isolated from the formosan gorgonian briareum excavatum. J. Nat. Prod. 1998, 61, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.C.; Lin, S.Y.; Chen, Y.J.; Wen, C.C.; Huang, C.Y.; Palanisamy, A.; Yang, N.S.; Sheu, J.H. Topical application of marine briarane-type diterpenes effectively inhibits 12-O-tetradecanoylphorbol-13-acetate-induced inflammation and dermatitis in murine skin. J. Biomed. Sci. 2011, 18, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Lin, S.C.; Feng, C.W.; Chen, P.C.; Su, Y.D.; Li, C.M.; Yang, S.N.; Jean, Y.H.; Sung, P.J.; Duh, C.Y.; et al. Anti-inflammatory and analgesic effects of the marine-derived compound excavatolide B isolated from the culture-type formosan gorgonian briareum excavatum. Mar. Drugs 2015, 13, 2559–2579. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Takayanagi, H. Regulation of bone by the adaptive immune system in arthritis. Arthritis Res. Ther. 2011, 13, 219. [Google Scholar] [CrossRef] [PubMed]
- Bendele, A. Animal models of rheumatoid arthritis. J. Musculoskelet. Neuronal Interact. 2001, 1, 377–385. [Google Scholar] [PubMed]
- Cremer, M.A.; Townes, A.S.; Kang, A.H. Adjuvant-induced arthritis in rats. Evidence that autoimmunity to homologous collagens types I, II, IX and XI is not involved in the pathogenesis of arthritis. Clin. Exp. Immunol. 1990, 82, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Trentham, D.E.; Townes, A.S.; Kang, A.H. Autoimmunity to type II collagen an experimental model of arthritis. J. Exp. Med. 1977, 146, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Kaminska, B. Mapk signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Thalhamer, T.; McGrath, M.A.; Harnett, M.M. Mapks and their relevance to arthritis and inflammation. Rheumatology 2008, 47, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Hou, G.Q.; Li, X.D.; Xia, X.; Liu, D.X.; Huang, D.Y.; Du, S.X. Quercetin triggers apoptosis of lipopolysaccharide (LPS)-induced osteoclasts and inhibits bone resorption in RAW264.7 cells. Cell. Physiol. Biochem. 2012, 30, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Chen, Y.H.; Chen, Y.H.; Su, Y.D.; Chang, Y.C.; Su, J.H.; Weng, C.F.; Lee, C.H.; Fang, L.S.; Wang, W.H.; et al. Briarane diterpenoids isolated from gorgonian corals between 2011 and 2013. Mar. Drugs 2014, 12, 2164–2181. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.H.; Park, H.C.; Choi, S.; Kim, S.; Bao, C.; Kim, H.W.; Choi, H.K.; Lee, H.J.; Auh, J.H. Metabolomic analysis reveals cyanidins in black raspberry as candidates for suppression of lipopolysaccharide-induced inflammation in murine macrophages. J. Agric. Food Chem. 2015, 63, 5449–5458. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.D.; Su, T.R.; Wen, Z.H.; Hwang, T.L.; Fang, L.S.; Chen, J.J.; Wu, Y.C.; Sheu, J.H.; Sung, P.J. Briarenolides K and L, new anti-inflammatory briarane diterpenoids from an Octocoral briareum sp. (briareidae). Mar. Drugs 2015, 13, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Shuto, T.; Jimi, E.; Kukita, T.; Hirata, M.; Koga, T. Granulocyte-macrophage colony stimulating factor suppresses lipopolysaccharide-induced osteoclast-like cell formation in mouse bone marrow cultures. Endocrinology 1994, 134, 831–837. [Google Scholar] [PubMed]
- Abu-Amer, Y.; Ross, F.P.; Edwards, J.; Teitelbaum, S.L. Lipopolysaccharide-stimulated osteoclastogenesis is mediated by tumor necrosis factor via its P55 receptor. J. Clin. Investig. 1997, 100, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- De Klerck, B.; Carpentier, I.; Lories, R.J.; Habraken, Y.; Piette, J.; Carmeliet, G.; Beyaert, R.; Billiau, A.; Matthys, P. Enhanced osteoclast development in collagen-induced arthritis in interferon-gamma receptor knock-out mice as related to increased splenic cd11b+ myelopoiesis. Arthritis Res. Ther. 2004, 6, R220–R231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.C.; Wang, H.; Jreyssaty, C.; Benderdour, M.; Lavigne, P.; Qiu, X.; Winnik, F.M.; Zhang, X.; Dai, K.; Shi, Q. Bone-protective effects of nonviral gene therapy with folate-chitosan DNA nanoparticle containing interleukin-1 receptor antagonist gene in rats with adjuvant-induced arthritis. Mol. Ther. 2008, 16, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Hoshino, M.; Aoki, K.; Ohya, K.; Komano, Y.; Nanki, T.; Miyasaka, N.; Umezawa, K. NF-kappab inhibitor dehydroxymethylepoxyquinomicin suppresses osteoclastogenesis and expression of NFATc1 in mouse arthritis without affecting expression of RANKL, osteoprotegerin or macrophage colony-stimulating factor. Arthritis Res. Ther. 2007, 9, R97. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Sakai, E.; Sakamoto, H.; Fumimoto, R.; Fukuma, Y.; Nishishita, K.; Okamoto, K.; Tsukuba, T. Inhibitory effects of tert-butylhydroquinone on osteoclast differentiation via up-regulation of heme oxygenase-1 and down-regulation of HMGB1 release and NFATc1 expression. J. Appl. Toxicol. 2014, 34, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Takahashi, N.; Katagiri, T.; Tamura, T.; Wada, S.; Findlay, D.M.; Martin, T.J.; Hirota, H.; Taga, T.; Kishimoto, T.; et al. Interleukin (IL)-6 induction of osteoclast differentiation depends on IL-6 receptors expressed on osteoblastic cells but not on osteoclast progenitors. J. Exp. Med. 1995, 182, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Redlich, K.; Hayer, S.; Ricci, R.; David, J.P.; Tohidast-Akrad, M.; Kollias, G.; Steiner, G.; Smolen, J.S.; Wagner, E.F.; Schett, G. Osteoclasts are essential for TNF-alpha-mediated joint destruction. J. Clin. Investig. 2002, 110, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- De Gois, S.; Schafer, M.K.; Defamie, N.; Chen, C.; Ricci, A.; Weihe, E.; Varoqui, H.; Erickson, J.D. Homeostatic scaling of vesicular glutamate and gaba transporter expression in rat neocortical circuits. J. Neurosci. 2005, 25, 7121–7133. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.J.; Chen, W.F.; Hung, H.C.; Jean, Y.H.; Sung, C.S.; Chakraborty, C.; Lee, H.P.; Chen, N.F.; Wen, Z.H. Effects of propofol on proliferation and anti-apoptosis of neuroblastoma SH-SY5Y cell line: New insights into neuroprotection. Brain Res. 2011, 1384, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Hla, T.; Maier, J.A.; Crofford, L.J.; Case, J.P.; Maciag, T.; Wilder, R.L. In vivo cyclooxygenase expression in synovial tissues of patients with rheumatoid arthritis and osteoarthritis and rats with adjuvant and streptococcal cell wall arthritis. J. Clin. Investig. 1992, 89, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Mazzon, E.; Dugo, L.; Serraino, I.; De Sarro, A.; Caputi, A.P.; Cuzzocrea, S. Amelioration of joint disease in a rat model of collagen-induced arthritis by M40403, a superoxide dismutase mimetic. Arthritis Rheumatol. 2001, 44, 2909–2921. [Google Scholar] [CrossRef]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-induced arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Jean, Y.H.; Lee, H.P.; Chen, W.F.; Sun, Y.M.; Su, J.H.; Lu, Y.; Huang, S.Y.; Hung, H.C.; Sung, P.J.; et al. A soft coral-derived compound, 11-epi-sinulariolide acetate suppresses inflammatory response and bone destruction in adjuvant-induced arthritis. PLoS ONE 2013, 8, e62926. [Google Scholar] [CrossRef] [PubMed]
- Bas, D.B.; Su, J.; Sandor, K.; Agalave, N.M.; Lundberg, J.; Codeluppi, S.; Baharpoor, A.; Nandakumar, K.S.; Holmdahl, R.; Svensson, C.I. Collagen antibody-induced arthritis evokes persistent pain with spinal glial involvement and transient prostaglandin dependency. Arthritis Rheumatol. 2012, 64, 3886–3896. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, H.; Matsui, Y.; Hayashida, K.; Yamane, S.; Maeda-Tanimura, M.; Nampei, A.; Hashimoto, J.; Suzuki, R.; Yoshikawa, H.; Ochi, T. Tartrate resistant acid phosphatase (TRAP) positive cells in rheumatoid synovium may induce the destruction of articular cartilage. Ann. Rheum. Dis. 2003, 62, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Morko, J.; Kiviranta, R.; Joronen, K.; Saamanen, A.M.; Vuorio, E.; Salminen-Mankonen, H. Spontaneous development of synovitis and cartilage degeneration in transgenic mice overexpressing cathepsin k. Arthritis Rheumatol. 2005, 52, 3713–3717. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.P.; Lin, Y.Y.; Duh, C.Y.; Huang, S.Y.; Wang, H.M.; Wu, S.F.; Lin, S.C.; Jean, Y.H.; Wen, Z.H. Lemnalol attenuates mast cell activation and osteoclast activity in a gouty arthritis model. J. Pharm. Pharmacol. 2015, 67, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Lu, L.J.; Fu, Q.; Dai, M.; Teng, J.L.; Fan, W.; Chen, S.L.; Ye, P.; Shen, N.; Huang, X.F.; et al. T-614, a novel immunomodulator, attenuates joint inflammation and articular damage in collagen-induced arthritis. Arthritis Res. Ther. 2008, 10. [Google Scholar] [CrossRef] [PubMed]














© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-Y.; Jean, Y.-H.; Lee, H.-P.; Lin, S.-C.; Pan, C.-Y.; Chen, W.-F.; Wu, S.-F.; Su, J.-H.; Tsui, K.-H.; Sheu, J.-H.; et al. Excavatolide B Attenuates Rheumatoid Arthritis through the Inhibition of Osteoclastogenesis. Mar. Drugs 2017, 15, 9. https://doi.org/10.3390/md15010009
Lin Y-Y, Jean Y-H, Lee H-P, Lin S-C, Pan C-Y, Chen W-F, Wu S-F, Su J-H, Tsui K-H, Sheu J-H, et al. Excavatolide B Attenuates Rheumatoid Arthritis through the Inhibition of Osteoclastogenesis. Marine Drugs. 2017; 15(1):9. https://doi.org/10.3390/md15010009
Chicago/Turabian StyleLin, Yen-You, Yen-Hsuan Jean, Hsin-Pai Lee, Sung-Chun Lin, Chieh-Yu Pan, Wu-Fu Chen, Shu-Fen Wu, Jui-Hsin Su, Kuan-Hao Tsui, Jyh-Horng Sheu, and et al. 2017. "Excavatolide B Attenuates Rheumatoid Arthritis through the Inhibition of Osteoclastogenesis" Marine Drugs 15, no. 1: 9. https://doi.org/10.3390/md15010009
APA StyleLin, Y. -Y., Jean, Y. -H., Lee, H. -P., Lin, S. -C., Pan, C. -Y., Chen, W. -F., Wu, S. -F., Su, J. -H., Tsui, K. -H., Sheu, J. -H., Sung, P. -J., & Wen, Z. -H. (2017). Excavatolide B Attenuates Rheumatoid Arthritis through the Inhibition of Osteoclastogenesis. Marine Drugs, 15(1), 9. https://doi.org/10.3390/md15010009