Accumulation and Tissue Distribution of Dinophysitoxin-1 and Dinophysitoxin-3 in the Mussel Crenomytilus grayanus Feeding on the Benthic Dinoflagellate Prorocentrum foraminosum

Abstract
:1. Introduction
2. Results
2.1. Diarrhetic Shellfish Toxins in C. grayanus during Acclimatization
2.2. DSTs during Feeding on C. muelleri (Control Study)
2.3. Dinophysistoxin-1 Exposure
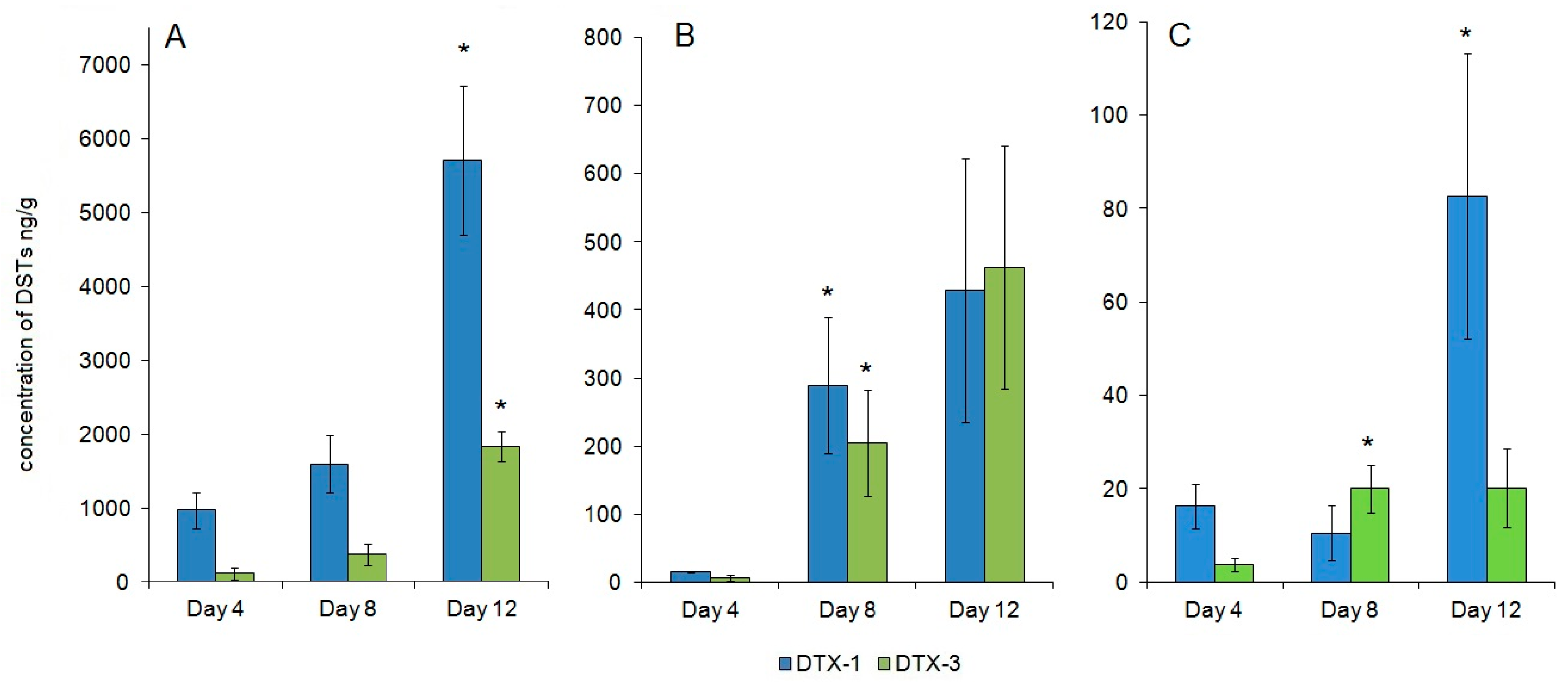
2.4. DSTs during Feeding on P. foraminosum
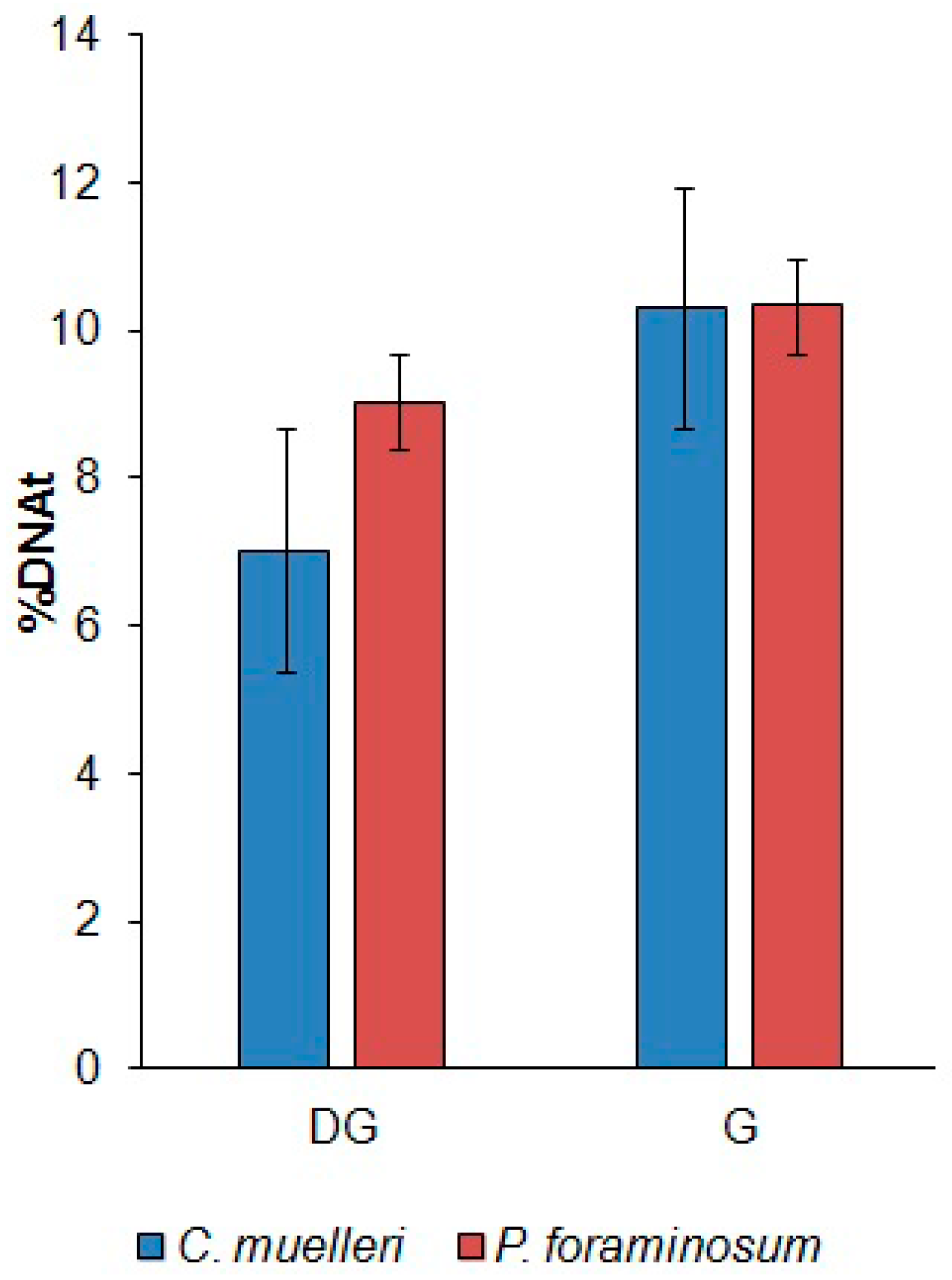
2.5. Alkaline Comet Assay
3. Discussion
3.1. Acclimatization and Feeding with C. muelleri
3.2. P. foraminosum as a Source of DXT-1 and the Exposure to DXT-1
3.3. DSTs Accumulation
3.4. Influence of DTX-1 on DNA Damage
4. Materials and Methods
4.1. Reagents
4.2. Culturing of Microalgae
4.3. Cell Number Counting
4.4. Mussel Acclimatization
4.5. Mussel Feeding
4.6. Sampling of Mussels
4.7. Toxin Extraction
4.7.1. Mussels’ Tissues
4.7.2. Algal Cells
4.7.3. Media
4.7.4. Toxin Hydrolysis
4.8. High-Performance Liquid Chromatography Mass-Spectrometric Analysis of DTX-1
4.9. Using Monensin as an Internal Standard
4.10. Quantitative DTX-1 Calculations
4.11. HPLC-MS Analysis of DTX-3
4.12. Lipids Extraction
4.13. Analysis of Fatty Acids Composition of P. foraminosum and C. grayanus
4.14. Isolation of Gill and Digestive Gland Cells for Alkaline Comet Assay
4.15. Alkaline Comet Assay
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hoppenrath, M.; Chomerat, N.; Horiguchi, T.; Schweikert, M.; Nagahama, Y.; Murray, S. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)-A proposal and review. Harmful Algae 2013, 27, 1–28. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suarez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Takai, A.; Murata, M.; Torigoe, K.; Isobe, M.; Mieskes, G.; Yasumoto, T. Inhibitory effect of okadaic acid derivatives on protein phosphatases. A study on structure-affinity relationship. Biochem. J. 1992, 284, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Ceulemans, H.; Bollen, M. Functional diversity of protein phosphatase-1, a cellular economizer and reset button. Physiol. Rev. 2004, 84, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.K.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A.D.M. First confirmation of human diarrhoeic poisonings by okadaic acid esters after ingestion of razor clams (Solen marginatus) and green crabs (Carcinus maenas) in Aveiro lagoon, Portugal and detection of okadaic acid esters in phytoplankton. Toxicon 2002, 40, 989–996. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pasaro, E.; Mendez, J.; Laffon, B. Okadaic Acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed]
- Marr, J.C.; Hu, T.; Pleasance, S.; Quilliam, M.A.; Wright, J.L.C. Detection of new 7-O-acyl derivatives of diarrhetic shellfish poisoning toxins by liquid chromatography-mass spectrometry. Toxicon 1992, 30, 1621–1630. [Google Scholar] [CrossRef]
- McNabb, P. Chemistry, metabolism and chemical analysis of okadaic acid group toxins. In Seafood and Freshwater Toxins; Botana, L., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 209–228. [Google Scholar]
- Vale, P.; Antónia, M.; Sampayo, M. Esters of okadaic acid and dinophysistoxin-2 in Portuguese bivalves related to human poisonings. Toxicon 1999, 37, 1109–1121. [Google Scholar] [CrossRef]
- Rodríguez, I.; Alfonso, A.; Antelo, A.; Alvarez, M.; Botana, L.M. Evaluation of the impact of mild steaming and heat treatment on the concentration of okadaic acid, dinophysistoxin-2 and dinophysistoxin-3 in mussels. Toxins 2016, 8, 175. [Google Scholar] [CrossRef] [PubMed]
- Konoki, K.; Onoda, T.; Watanabe, R.; Cho, Y.; Kaga, S.; Suzuki, T.; Yotsu-Yamashita, M. In vitro acylation of okadaic acid in the presence of various bivalves’ extracts. Mar. Drugs 2013, 11, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Prego-Faraldo, M.V.; Valdiglesias, V.; Laffon, B.; Mendez, J.; Eirin Lopez, J.M. Early genotoxic and cytotoxic effects of the toxic dinoflagellate Prorocentrum lima in the mussel Mytilus galloprovincialis. Toxins 2016, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Rossignoli, A.E.; Fernandez, D.; Regueiro, J.; Marino, C.; Blanco, J. Esterification of okadaic acid in the mussel Mytilus galloprovincialis. Toxicon 2011, 57, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ota, H.; Yamasaki, M. Direct evidence of transformation of dinophysistoxin-1 to 7-O-acyl-dinophysistoxin-1 (dinophysistoxin-3) in the scallop Patinopecten yessoensis. Toxicon 1999, 37, 187–198. [Google Scholar] [CrossRef]
- MacKenzie, L.; Holland, P.; McNabb, P.; Beuzenberg, V.; Selwood, A.; Suzuki, T. Complex toxin profiles in phytoplankton and Greenshell mussels (Perna canaliculus), revealed by LC-MS/MS analysis. Toxicon 2002, 40, 1321–1330. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A. Esterification of DSP toxins by Portuguese bivalves from the Northwest coast determined by LC-MS—A widespread phenomenon. Toxicon 2002, 40, 33–42. [Google Scholar] [CrossRef]
- Torgersen, T.; Sandvik, M.; Lundve, B.; Lindegarth, S. Profiles and levels of fatty acid esters of okadaic acid group toxins and pectenotoxins during toxin depuration. Part II: Blue mussels (Mytilus edulis) and flat oyster (Ostrea edulis). Toxicon 2008, 52, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Moroño, A.; Arévalo, F.; Fernández, M.L.; Maneiro, J.; Pazos, Y.; Salgado, C.; Blanco, J. Accumulation and transformation of DSP toxins in mussels Mytilus galloprovincialis during a toxic episode caused by Dinophysis acuminata. Aquat. Toxicol. 2003, 62, 269–280. [Google Scholar] [CrossRef]
- Blanco, J.; Mariño, C.; Martín, H.; Acosta, C.P. Anatomical distribution of diarrhetic shellfish poisoning (DSP) toxins in the mussel Mytilus galloprovincialis. Toxicon 2007, 50, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Marcaillou, C.; Haure, J.; Mondeguer, F.; Courcoux, A.; Dupuy, B.; Pénisson, C. Effect of food supply on the detoxification in the blue mussel, Mytilus edulis, contaminated by diarrhetic shellfish toxins. Aquat. Living Resour. 2010, 23, 255–266. [Google Scholar] [CrossRef]
- Bauder, A.G.; Cembella, A.D.; Bricelj, V.M.; Quilliam, M.A. Uptake and fate of diarrhetic shellfish poisoning toxins from the dinoflagellate Prorocentrum lima in the bay scallop Argopecten irradians. Mar. Ecol. Prog. Ser. 2001, 213, 39–52. [Google Scholar] [CrossRef]
- Kameneva, P.A.; Efimova, K.V.; Rybin, V.G.; Orlova, T.Y. Detection of dinophysistoxin-1 in clonal culture of marine dinoflagellate Prorocentrum foraminosum (Faust M.A., 1993) from the Sea of Japan. Toxins 2015, 7, 3947–3959. [Google Scholar] [CrossRef] [PubMed]
- Selina, M.S. Morphology and seasonal dynamics of the potentially toxic microalga Prorocentrum foraminosum Faust 1993 (Dinophyta) in Peter the Great Bay, the Sea of Japan. Russ. J. Mar. Biol. 2017, 43, 169–174. [Google Scholar] [CrossRef]
- Orlova, T.Y.; Kameneva, P.A.; Stonik, I.V.; Morozova, T.V.; Efimova, K.V.; Moore, L.; Eberhart, B.-T.L.; Wells, M.L.; Trainer, V.L. Diarrhetic shellfish toxins in Primorsky Krai, Russia. J. Shellfish Res. 2015, 34, 1151–1160. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Prassopoulou, E.; Katikou, P.; Georgantelis, D.; Kyritsakis, A. Detection of okadaic acid and related esters in mussels during diarrhetic shellfish poisoning (DSP) episodes in Greece using the mouse bioassay, the PP2A inhibition assay and HPLC with fluorimetric detection. Toxicon 2009, 53, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; Nguyen, L.; Aasen, J.; Keogh, M.; Kilcoyne, J.; McCarron, P.; Aune, T. Tissue distribution, effects of cooking and parameters affecting the extraction of azaspiracids from mussels, Mytilus edulis, prior to analysis by liquid chromatography coupled to mass spectrometry. Toxicon 2005, 46, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.T.; Hansen, P.J.; Krock, B.; Vismann, B. Accumulation, transformation and breakdown of DSP toxins from the toxic dinoflagellate Dinophysis acuta in blue mussels, Mytilus edulis. Toxicon 2016, 117, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Kamiyama, T.; Okumura, Y.; Ishihara, K.; Matsushima, R.; Kaneniwa, M. Liquid-chromatographic hybrid triple-quadrupole linear-ion-trap MS/MS analysis of fatty-acid esters of dinophysistoxin-1 in bivalves and toxic dinoflagellates in Japan. Fish. Sci. 2009, 75, 1039–1048. [Google Scholar] [CrossRef]
- Vale, P. Detailed profiles of 7-O-acyl esters in plankton and shellfish from the Portuguese coast. J. Chromatogr. A 2006, 1128, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, R.; Uchida, H.; Nagai, S.; Watanabe, R.; Kamio, M.; Nagai, H.; Kaneniwa, M.; Suzuki, T. Assimilation, accumulation, and metabolism of dinophysistoxins (DTXs) and pectenotoxins (PTXs) in the several tissues of japanese scallop Patinopecten yessoensis. Toxins 2015, 7, 5141–5154. [Google Scholar] [CrossRef] [PubMed]
- European Union Reference Laboratory for Marine Biotoxins. EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS; Version 5; European Union Reference Laboratory for Marine Biotoxins: Vigo, Spain, 2015; pp. 1–31. Available online: www.aesan.msps.es/en/CRLMB/web/home.shtml (accessed on 24 October 2017).
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipids extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Christie, W.W. Equivalent chain-lengths of methyl ester derivatives of fatty acids on gas chromatography A reappraisal. J. Chromatogr. A 1988, 447, 305–314. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Scheider, E.L. A simple technique for quantification of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 237, 123–130. [Google Scholar]
- Slobodskova, V.V.; Solodova, E.E.; Slinko, E.N.; Chelomin, V.P. Evaluation of the genotoxicity of cadmium in gill cells of the clam Corbicula japonica using the comet assay. Russ. J. Mar. Biol. 2010, 36, 311–315. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Days of Feeding | Culture Volume (mL) | Cell Density (×103 Cells/mL) | Total Cells Added (×103 Cells) | Initial Cell Density in Tanks (×103 Cell/L) | Cell Density in Tanks after 24 h (×103 Cells/L) | Total Cells Ingested (×103 Cells) |
|---|---|---|---|---|---|---|
| Day 1 | 90 | 9.1 | 820.4 | 91.2 | 10.5 | 726.1 |
| Day 2 | 95 | 6.9 | 655.5 | 72.8 | 4.3 | 616.5 |
| Day 3 | 100 | 9.6 | 963.3 | 107.0 | 7.0 | 900.1 |
| Day 4 | 102 | 10.2 | 1041.1 | 115.7 | 16.7 | 891.3 |
| Day 5 | 100 | 10.7 | 1067.3 | 118.6 | 54.8 | 574.0 |
| Day 6 | 100 | 5.9 | 588.0 | 65.3 | 19.2 | 415.6 |
| Day 7 | 100 | 10.2 | 1015.0 | 112.7 | 24.7 | 792.1 |
| Day 8 | 100 | 7.5 | 750.0 | 83.3 | 65.8 | 157.7 |
| Day 9 | 80 | 11.6 | 925.6 | 102.8 | 0 | 925.6 |
| Day 10 | 87 | 8.2 | 716.6 | 79.6 | 25.4 | 488.3 |
| Day 11 | 85 | 12.7 | 1082.3 | 120.3 | 24.7 | 860.0 |
| Day 12 | 90 | 6.1 | 551.4 | 61.2 | 23.0 | 344.3 |
| Source of DTX-1 | Day 4 | Day 8 | Day 12 |
|---|---|---|---|
| DTX-1 from cells (µg) | 5.95 | 9.63 | 14.61 |
| DTX-1 from media (µg) | 16.95 | 34.47 | 49.50 |
| Acyl Group | Acyl Chain of DTX-3 1 | Fatty Acids (FA) 2 | |||
|---|---|---|---|---|---|
| Digestive Gland | Kidney | P. foraminosum | C. grayanus Digestive Gland Neutral Lipids | C. grayanus Digestive Gland Polar Lipids | |
| 14:0 | 6.15 ± 1.47 | 5.12 ± 0.76 | 11.09 | 9.90 | 11.52 |
| 15:0 | 1.33 ± 0.67 | nd | nd | 0.73 | 0.29 |
| 16:0 | 34.98 ± 2.01 | 38.49 ± 0.74 | 16.02 | 15.46 | 12.59 |
| 17:0 | nd | nd | nd | 1.57 | 1.78 |
| 18:0 | 3.31 ± 0.71 | 3.76 ± 1.06 | 1.38 | 2.27 | 15.27 |
| 20:0 | nd | nd | 0.19 | 0.10 | 0.05 |
| 22:0 | nd | nd | nd | 0.02 | nd |
| 14:1 | nd | nd | 0.29 | 0.26 | 0.07 |
| 16:1 * | 27.65 ± 4.96 | 25.34 ± 2.39 | 2.90 | 16.32 | 1.96 |
| 18:1 * | 25.66 ± 4.32 | 26.37 ± 4.18 | 23.74 | 6.31 | 2.58 |
| 20:1 | nd | nd | nd | 3.36 | 8.49 |
| 18:2 * | 2.50 ± 1.05 | 1.90 ± 0.00 | 0.17 | 2.67 | 0.61 |
| 18:3 | 0.36 ± 0.00 | nd | 1.84 | 0.93 | 0.37 |
| 18:4 | 0.20 ± 0.00 | nd | 4.17 | 0.39 | 0.15 |
| 18:5 | nd | nd | 2.50 | nd | nd |
| 20:4 | nd | nd | nd | 1.94 | 4.74 |
| 20:5 | nd | nd | 12.38 | 20.60 | 8.42 |
| 22:6 | nd | nd | 18.62 | 6.26 | 5.43 |
| Other | na | na | 4.71 | 10.93 | 19.16 |
| Organ | Mussel 1 | Mussel 2 | Mussel 3 | Mussel 4 | Mussel 5 |
|---|---|---|---|---|---|
| Digestive gland | 9.679 | 13.306 | 5.245 | 5.811 | 1.229 |
| Kidneys | 0.064 | 0.905 | 0.026 | 0.181 | 0.002 |
| Gills | 0.060 | 0.012 | 0.062 | 0.060 | 0.023 |
| Total per mussel (µg) | 9.803 | 14.223 | 5.333 | 6.052 | 1.254 |
| ∑ total of mussels (µg) | - | - | - | - | 36.665 |
| Compound | Certified Concentration | Measured Concentration |
|---|---|---|
| Active forms | ||
| Okadaic acid | 1.07 ± 0.08 | 1.039 |
| Dinophysistoxin-1 | 1.07 ± 0.11 | 0.963 |
| Dinophysistoxin-2 | 0.86 ± 0.08 | 0.699 |
| Active+conjugated forms | ||
| Okadaic acid | 2.4 | 2.450 |
| Dinophysistoxin-1 | 1.1 | 0.998 |
| Dinophysistoxin-2 | 2.2 | 2.123 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kameneva, P.A.; Krasheninina, E.A.; Slobodskova, V.V.; Kukla, S.P.; Orlova, T.Y. Accumulation and Tissue Distribution of Dinophysitoxin-1 and Dinophysitoxin-3 in the Mussel Crenomytilus grayanus Feeding on the Benthic Dinoflagellate Prorocentrum foraminosum. Mar. Drugs 2017, 15, 330. https://doi.org/10.3390/md15100330
Kameneva PA, Krasheninina EA, Slobodskova VV, Kukla SP, Orlova TY. Accumulation and Tissue Distribution of Dinophysitoxin-1 and Dinophysitoxin-3 in the Mussel Crenomytilus grayanus Feeding on the Benthic Dinoflagellate Prorocentrum foraminosum. Marine Drugs. 2017; 15(10):330. https://doi.org/10.3390/md15100330
Chicago/Turabian StyleKameneva, Polina A., Ekaterina A. Krasheninina, Valentina V. Slobodskova, Sergey P. Kukla, and Tatiana Yu. Orlova. 2017. "Accumulation and Tissue Distribution of Dinophysitoxin-1 and Dinophysitoxin-3 in the Mussel Crenomytilus grayanus Feeding on the Benthic Dinoflagellate Prorocentrum foraminosum" Marine Drugs 15, no. 10: 330. https://doi.org/10.3390/md15100330
APA StyleKameneva, P. A., Krasheninina, E. A., Slobodskova, V. V., Kukla, S. P., & Orlova, T. Y. (2017). Accumulation and Tissue Distribution of Dinophysitoxin-1 and Dinophysitoxin-3 in the Mussel Crenomytilus grayanus Feeding on the Benthic Dinoflagellate Prorocentrum foraminosum. Marine Drugs, 15(10), 330. https://doi.org/10.3390/md15100330