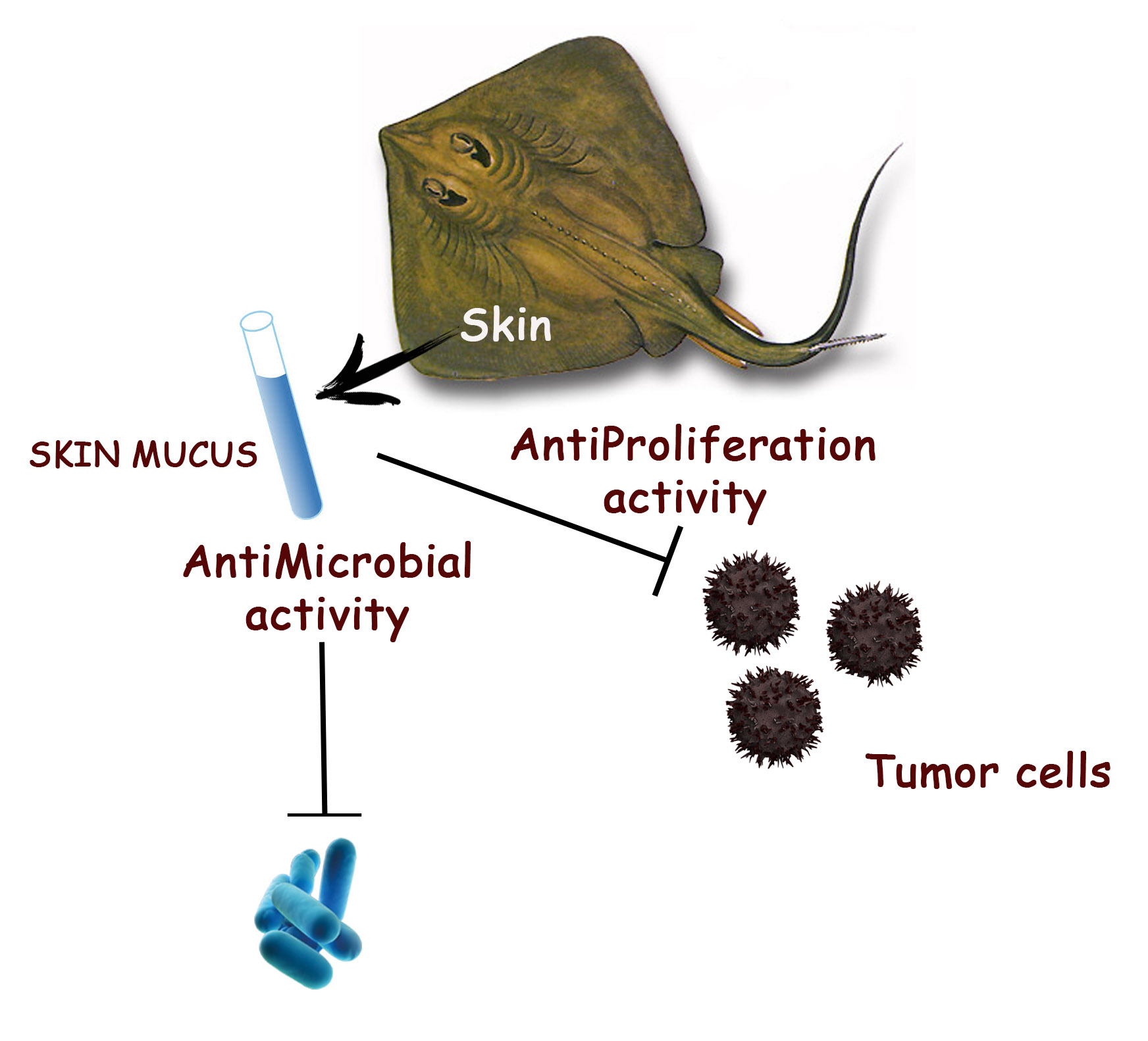
Antimicrobial and Anti-Proliferative Effects of Skin Mucus Derived from Dasyatis pastinaca (Linnaeus, 1758)

 ,
,  ,
, 
 , ,
, ,  , ,
, ,  ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Growth Inhibition Curves by Cell Counting
2.2. Antifungal Assay
2.3. Western Blot Analysis
2.4. Chitotriosidase Activity Assay
2.5. Cell Cultures
2.6. Cell Viability Assay
2.7. Bacterial Grown Inibition Calculation
2.8. Statistical Analysis
3. Results
3.1. Bacterial and Fungal Growth Inhibition Curves
3.2. Chitinase Expression and Activity
3.3. Anti-Proliferative Activity
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lewis, K. New approaches to antimicrobial discovery. Biochem. Pharmacol. 2017, 134, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Shephard, K.L. Functions for fish mucus. Rev. Fish Biol. Fish. 1994, 4, 401–429. [Google Scholar] [CrossRef]
- Kumari, U.; Nigam, A.K.; Mitial, S.; Mitial, A.K. Antibacterial properties of the skin mucus of the freshwater fishes, Rita rita and Channa punctatus. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 781–786. [Google Scholar] [PubMed]
- Austin, B.; McIntosh, D. Natural antibacterial com- pounds on the surface of rainbow trout, Salmo gairdneri Richardson. J. Fish Dis. 1988, 11, 275–277. [Google Scholar] [CrossRef]
- Nagashima, Y.; Sendo, A.; Shimakura, K.; Shiomi, K.; Kobayashi, T.; Kimura, B.; Fujii, T. Antibacterial factors in skin mucus of rabbitfishes. J. Fish Biol. 2001, 58, 1761–1765. [Google Scholar] [CrossRef]
- Hellio, C.; Pons, A.M.; Beaupoil, C.; Bourgougnon, N.; Gal, Y.L. Antibacterial, antifungal and cytotoxic activities of extracts from fish epidermis and epidermal mucus. Int. J. Antimicrob. Agents 2002, 20, 214–219. [Google Scholar] [CrossRef]
- Kuppulakshmi, C.; Prakash, M.; Gunasekaran, G.; Manimegalai, G.; Sarojini, S. Antibacterial properties of fish mucus from Channa punctatus and Cirrhinus mrigala. Eur. Rev. Med. Pharmacol. Sci. 2008, 12, 149–153. [Google Scholar] [PubMed]
- Wei, O.Y.; Xavier, R.; Marimuthu, K. Screening of antibacterial activity of mucus extract of snakehead fish, Channa striatus (Bloch). Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 675–681. [Google Scholar] [PubMed]
- Alexander, J.B.; Ingram, G.A. Noncellular nonspecific defence mechanisms of fish. Ann. Rev. Fish Dis. 1992, 2, 249–279. [Google Scholar] [CrossRef]
- Villarroel, F.; Bastias, A.; Casado, A.; Amthauer, R.; Concha, M.I. Apolipoprotein A-I, an antimicrobial protein in Oncorhynchus mykiss: Evaluation of its expression in primary defence barriers and plasma levels in sick and healthy fish. Fish Shellfish Immunol. 2007, 23, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Kitani, Y.; Tsukamoto, C.; Zhang, G.; Nagai, H.; Ishida, M.; Ishizaki, S.; Shimakura, K.; Shiomi, K.; Nagashima, Y. Identification of an antibacterial protein as l-amino acid oxidase in the skin mucus of rockfish Sebastes schlegeli. FEBS J. 2007, 274, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Ben Bacha, A.; Abid, I.; Horchani, H.; Mejdoub, H. Enzymatic properties of stingray Dasyatis pastinaca group V, IIA and IB phospholipases A(2): A comparative study. Int. J. Biol. Macromol. 2013, 62, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Ben Bacha, A.; Abid, I.; Horchani, H. Antibacterial properties of intestinal phospholipase A2 from the common stingray Dasyatis pastinaca. Appl. Biochem. Biotechnol. 2012, 168, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Kwak, C.H.; Lee, S.H.; Lee, S.K.; Ha, S.H.; Suh, S.J.; Kwon, K.M.; Chung, T.W.; Ha, K.T.; Chang, Y.C.; Lee, Y.C.; et al. Induction of apoptosis and antitumor activity of eel skin mucus, containing lactose-binding molecules, on human leukemic K562 cells. Mar. Drugs 2015, 13, 3936–3949. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI Supplement M100S; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2017; ISBN 1-56238-804-5. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Ausceptibility Testing of Yeast, 3rd ed.; Approved Standard M27-A3; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2008; ISBN 1-56238-666-2. [Google Scholar]
- Sacerdoti, D.; Colombrita, C.; Ghattas, M.H.; Ismaeil, E.F.; Scapagnini, G.; Bolognesi, M.; Li Volti, G.; Abraham, N.G. Heme oxygenase-1 transduction in endothelial cells causes downregulation of monocyte chemoattractant protein-1 and of genes involved in inflammation and growth. Cell. Mol. Biol. 2005, 51, 363–370. [Google Scholar] [PubMed]
- Torella, D.; Leosco, D.; Indolfi, C.; Curcio, A.; Coppola, C.; Ellison, G.M.; Russo, V.G.; Torella, M.; Li Volti, G.; Rengo, F.; et al. Aging exacerbates negative remodeling and impairs endothelial regeneration after balloon injury. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H2850–H2860. [Google Scholar] [CrossRef] [PubMed]
- Salomone, F.; Barbagallo, I.; Puzzo, L.; Piazza, C.; Li Volti, G. Efficacy of adipose tissue-mesenchymal stem cell transplantation in rats with acetaminophen liver injury. Stem Cell Res. 2013, 11, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Tibullo, D.; Barbagallo, I.; Giallongo, C.; La Cava, P.; Parrinello, N.; Vanella, L.; Stagno, F.; Palumbo, G.A.; Li Volti, G.; Di Raimondo, F. Nuclear translocation of heme oxygenase-1 confers resistance to imatinib in chronic myeloid leukemia cells. Curr. Pharm. Des. 2013, 19, 2765–2770. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Grasso, S.; Tomassoni, D.; Traini, E.; Raciti, G.; Viola, M.; Li Volti, G.; Campisi, A.; Amenta, F.; Avola, R. Effect of growth factors and steroid hormones on heme oxygenase and cyclin D1 expression in primary astroglial cell cultures. J. Neurosci. Res. 2015, 93, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Bramanti, V.; Bronzi, D.; Tomassoni, D.; Li Volti, G.; Cannavo, G.; Raciti, G.; Napoli, M.; Vanella, A.; Campisi, A.; Ientile, R.; et al. Effect of choline-containing phospholipids on transglutaminase activity in primary astroglial cell cultures. Clin. Exp. Hypertens. 2008, 30, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Tibullo, D.; Caporarello, N.; Giallongo, C.; Anfuso, C.D.; Genovese, C.; Arlotta, C.; Puglisi, F.; Parrinello, N.L.; Bramanti, V.; Romano, A.; et al. Antiproliferative and antiangiogenic effects of Punica granatum juice (PGJ) in multiple myeloma (MM). Nutrients 2016, 8, 611. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Ramalingam, S.; Nagegowda, D.; Taylor, P.W.; Chye, M.L. Brassica juncea chitinase BjCHI1 inhibits growth of fungal phytopathogens and agglutinates Gram-negative bacteria. J. Exp. Bot. 2008, 59, 3475–3484. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Gustafson, K.R. Marine pharmacology in 2005–2006: Antitumour and cytotoxic compounds. Eur. J. Cancer 2008, 44, 2357–2387. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Hamann, M.T. Marine pharmacology in 2000: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antituberculosis, and antiviral activities; affecting the cardiovascular, immune, and nervous systems and other miscellaneous mechanisms of action. Mar. Biotechnol. 2004, 6, 37–52. [Google Scholar] [PubMed]
- Balasubramanian, S.; Revathi, A.; Gunasekaran, G. Studies on anticancer, haemolytic activity and chemical composition of crude epidermal mucus of fish Mugil cephalus. Int. J. Fish. Aquat. Stud. 2016, 4, 438–443. [Google Scholar]
- Nissen-Meyer, J.; Nes, I.F. Ribosomally synthesized antimicrobial peptides: Their function, structure, biogenesis, and mechanism of action. Arch. Microbiol. 1997, 167, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shen, Z.; Wu, J. Expression, purification and in vitro antifungal activity of acidic mammalian chitinase against Candida albicans, Aspergillus fumigatus and Trichophyton rubrum strains. Clin. Exp. Dermatol. 2009, 34, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Mu, Y.; Xiang, L.X.; Fan, D.; Feng, M.; Zhang, S.; Shi, Q.; Zhu, L.Y.; Li, T.; Ding, Y.; et al. Genome sequencing of the perciform fish Larimichthys crocea provides insights into molecular and genetic mechanisms of stress adaptation. PLoS Genet. 2015, 11, e1005118. [Google Scholar] [CrossRef] [PubMed]
- Provan, F.; Jensen, L.B.; Uleberg, K.E.; Larssen, E.; Rajalahti, T.; Mullins, J.; Obach, A. Proteomic analysis of epidermal mucus from sea lice-infected Atlantic salmon, Salmo salar L. J. Fish Dis. 2013, 36, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Valdenegro-Vega, V.A.; Crosbie, P.; Bridle, A.; Leef, M.; Wilson, R.; Nowak, B.F. Differentially expressed proteins in gill and skin mucus of Atlantic salmon (Salmo salar) affected by amoebic gill disease. Fish Shellfish Immunol. 2014, 40, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Brinchmann, M.F. Immune relevant molecules identified in the skin mucus of fish using-omics technologies. Mol. Biosyst. 2016, 12, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Cha, G.H.; Liu, Y.; Peng, T.; Huang, M.Z.; Xie, C.Y.; Xiao, Y.C.; Wang, W.N. Molecular cloning, expression of a galectin gene in Pacific white shrimp Litopenaeus vannamei and the antibacterial activity of its recombinant protein. Mol. Immunol. 2015, 67, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Pockley, A.G.; Muthana, M.; Calderwood, S.K. The dual immunoregulatory roles of stress proteins. Trends Biochem. Sci. 2008, 33, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; McBride, R.; Berger, O.; Razi, N.; Heimburg-Molinaro, J.; Rodrigues, L.C.; Gourdine, J.P.; Noll, A.J.; von Gunten, S.; et al. Microbial glycan microarrays define key features of host-microbial interactions. Nat. Chem. Biol. 2014, 10, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Tibullo, D.; Di Rosa, M.; Giallongo, C.; La Cava, P.; Parrinello, N.L.; Romano, A.; Conticello, C.; Brundo, M.V.; Saccone, S.; Malaguarnera, L.; et al. Bortezomib modulates CHIT1 and YKL40 in monocyte-derived osteoclast and in myeloma cells. Front. Pharmacol. 2015, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L. Chitotriosidase: The yin and yang. Cell. Mol. Life Sci. 2006, 63, 3018–3029. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M.; Tibullo, D.; Saccone, S.; Distefano, G.; Basile, M.S.; Di Raimondo, F.; Malaguarnera, L. CHI3L1 nuclear localization in monocyte derived dendritic cells. Immunobiology 2016, 221, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Koch, B.E.; Stougaard, J.; Spaink, H.P. Keeping track of the growing number of biological functions of chitin and its interaction partners in biomedical research. Glycobiology 2015, 25, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Kanneganti, M.; Kamba, A.; Mizoguchi, E. Role of chitotriosidase (chitinase 1) under normal and disease conditions. J. Epithel. Biol. Pharmacol. 2012, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, B.; Pathaw, N.; Chrungoo, N.K.; Bhattacharjee, A. Molecular modelling, dynamics simulation and characterization of antifungal chitinase from Sechium edule. Gene 2017, 606, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Berini, F.; Presti, I.; Beltrametti, F.; Pedroli, M.; Varum, K.M.; Pollegioni, L.; Sjoling, S.; Marinelli, F. Production and characterization of a novel antifungal chitinase identified by functional screening of a suppressive-soil metagenome. Microb. Cell Fact. 2017, 16, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ruan, X.; Wang, X.; Liu, Z.; Hu, L.; Li, C. Overexpression of a chitinase gene from trichoderma asperellum increases disease resistance in transgenic soybean. Appl. Biochem. Biotechnol. 2016, 180, 1542–1558. [Google Scholar] [CrossRef] [PubMed]
- Metwally, N.S.; Kholeif, T.E.; Ghanem, K.Z.; Farrag, A.R.; Ammar, N.M.; Abdel-Hamid, A.H. The protective effects of fish oil and artichoke on hepatocellular carcinoma in rats. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1429–1444. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains Tested | Escherichia coli ATCC 35218 | Escherichia coli ATCC 25922 | Klebsiella pneumoniae ATCC 700603 | Pseudomonas aeruginosa ATCC 27853 | ||||
|---|---|---|---|---|---|---|---|---|
| 16.50 µg/µL (22 h) | Control+ (22 h) | 16.50 µg/µL (22 h) | Control+ (22 h) | 16.50 µg/µL (22 h) | Control+ (22 h) | 16.50 µg/µL (22 h) | Control+ (22 h) | |
| Ymax (CFU/mL) | 6.74 × 108 | 9.24 × 108 | 6.97 × 108 | 9.07 × 108 | 4.83 × 108 | 8.11 × 108 | 7.36 × 108 | 8.87 × 108 |
| Ymax Standard Error (CFU/mL) | 4.99 × 106 | 6.08 × 106 | 5.39 × 106 | 4.17 × 106 | 3.23 × 106 | 7.37 × 106 | 5.91 × 106 | 7.36 × 106 |
| Inhibition (%) | 27.05 | 23.15 | 40.44 | 17.02 | ||||
| Strains | MIC µg·µL−1 | |
|---|---|---|
| 24 h | 48 h | |
| Candida albicans ATCC90028 | 4.12 | 4.12 |
| Candida albicans ATCC10231 | 4.12 | 4.12 |
| Candida albicans (6) clinical strain | 4.12 | 4.12 |
| Candida albicans (10) clinical strain | 4.12 | 4.12 |
| Candida glabrata (14) clinical strain | 4.12 | 4.12 |
| Candida tropicalis (21) clinical strain | 4.12 | 4.12 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuochi, V.; Li Volti, G.; Camiolo, G.; Tiralongo, F.; Giallongo, C.; Distefano, A.; Petronio Petronio, G.; Barbagallo, I.; Viola, M.; Furneri, P.M.; et al. Antimicrobial and Anti-Proliferative Effects of Skin Mucus Derived from Dasyatis pastinaca (Linnaeus, 1758). Mar. Drugs 2017, 15, 342. https://doi.org/10.3390/md15110342
Fuochi V, Li Volti G, Camiolo G, Tiralongo F, Giallongo C, Distefano A, Petronio Petronio G, Barbagallo I, Viola M, Furneri PM, et al. Antimicrobial and Anti-Proliferative Effects of Skin Mucus Derived from Dasyatis pastinaca (Linnaeus, 1758). Marine Drugs. 2017; 15(11):342. https://doi.org/10.3390/md15110342
Chicago/Turabian StyleFuochi, Virginia, Giovanni Li Volti, Giuseppina Camiolo, Francesco Tiralongo, Cesarina Giallongo, Alfio Distefano, Giulio Petronio Petronio, Ignazio Barbagallo, Maria Viola, Pio Maria Furneri, and et al. 2017. "Antimicrobial and Anti-Proliferative Effects of Skin Mucus Derived from Dasyatis pastinaca (Linnaeus, 1758)" Marine Drugs 15, no. 11: 342. https://doi.org/10.3390/md15110342
APA StyleFuochi, V., Li Volti, G., Camiolo, G., Tiralongo, F., Giallongo, C., Distefano, A., Petronio Petronio, G., Barbagallo, I., Viola, M., Furneri, P. M., Di Rosa, M., Avola, R., & Tibullo, D. (2017). Antimicrobial and Anti-Proliferative Effects of Skin Mucus Derived from Dasyatis pastinaca (Linnaeus, 1758). Marine Drugs, 15(11), 342. https://doi.org/10.3390/md15110342