LC-MS/MS Detection of Karlotoxins Reveals New Variants in Strains of the Marine Dinoflagellate Karlodinium veneficum from the Ebro Delta (NW Mediterranean)



 and
and 
Abstract
:1. Introduction
2. Results
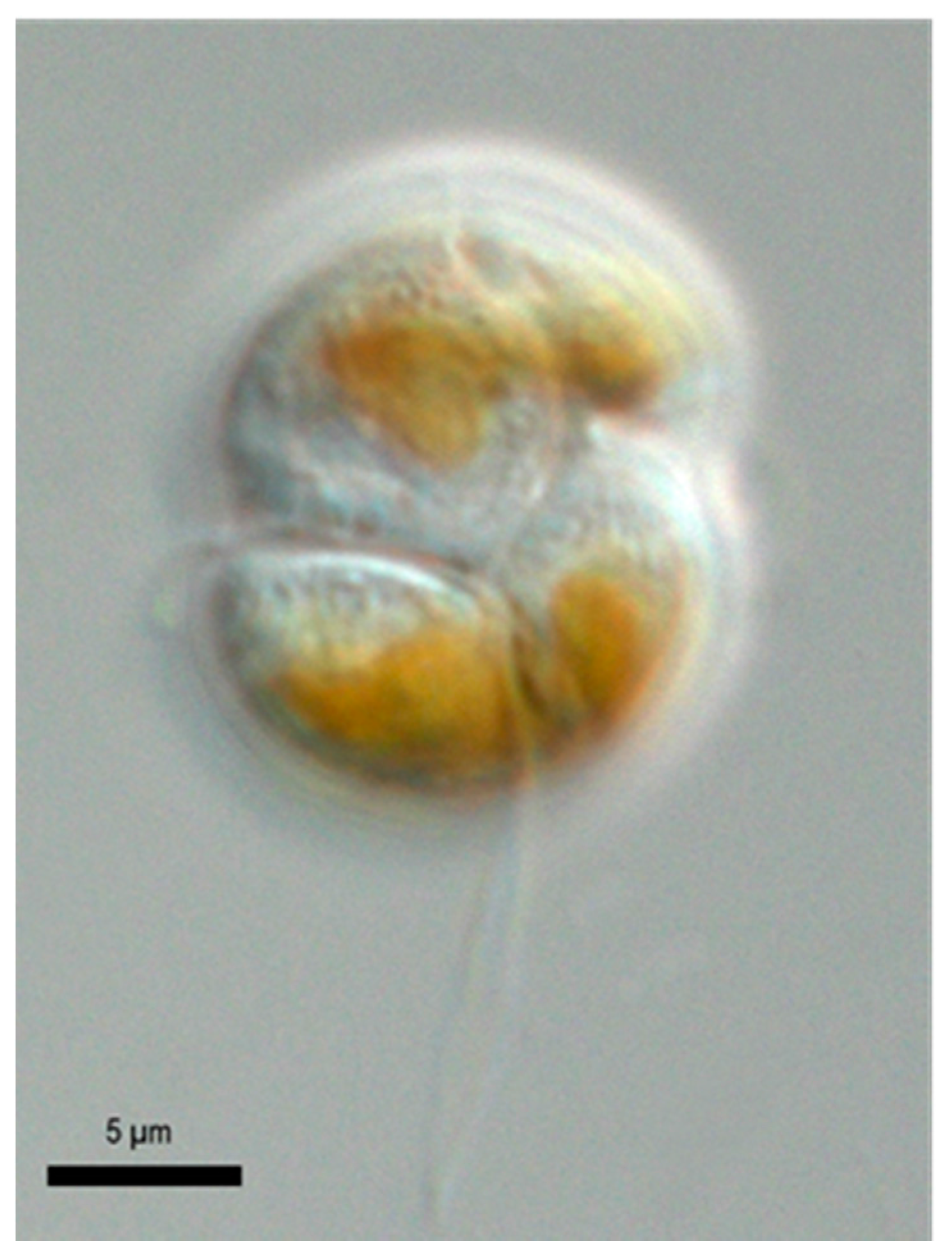
2.1. Species Identification
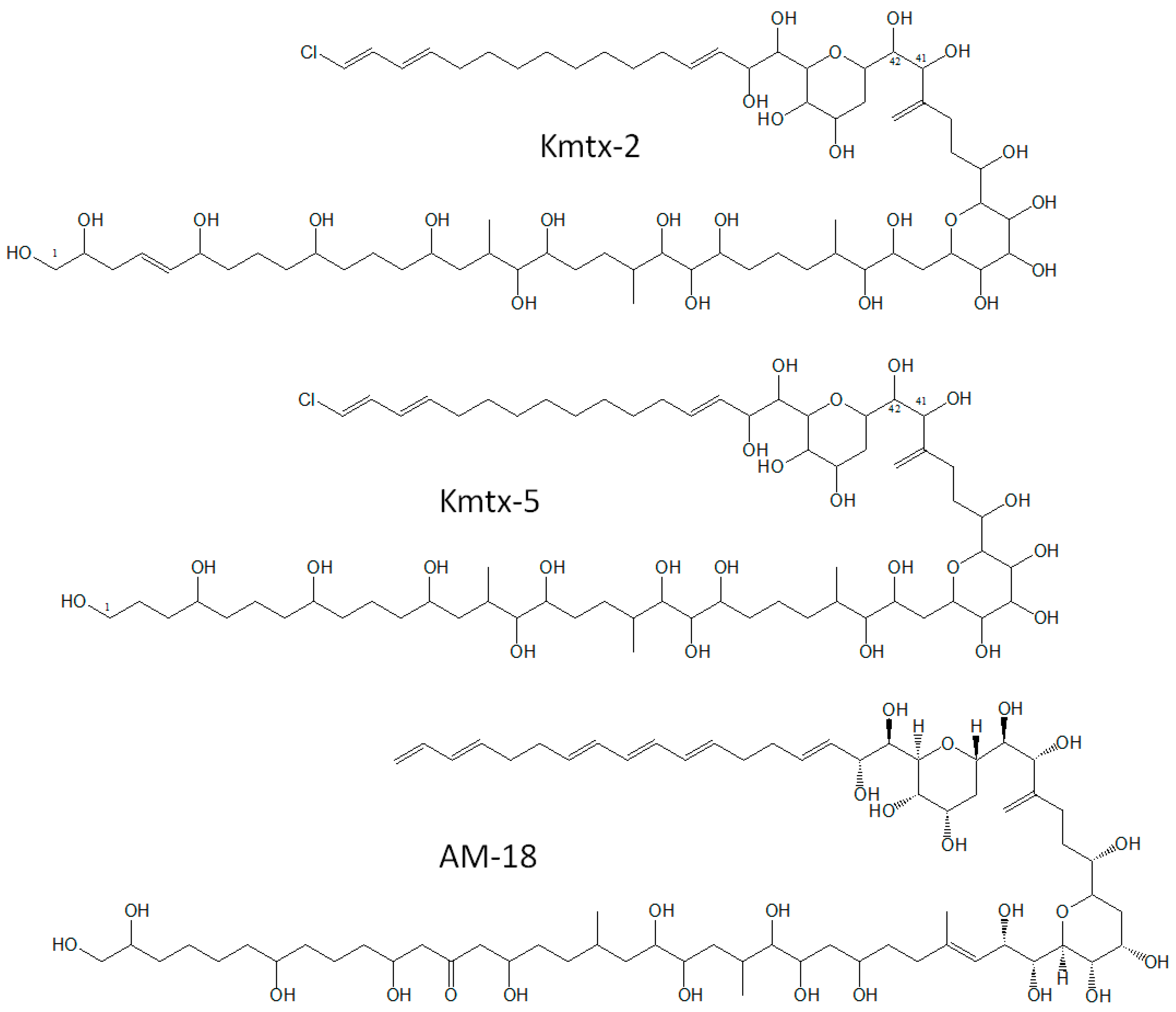
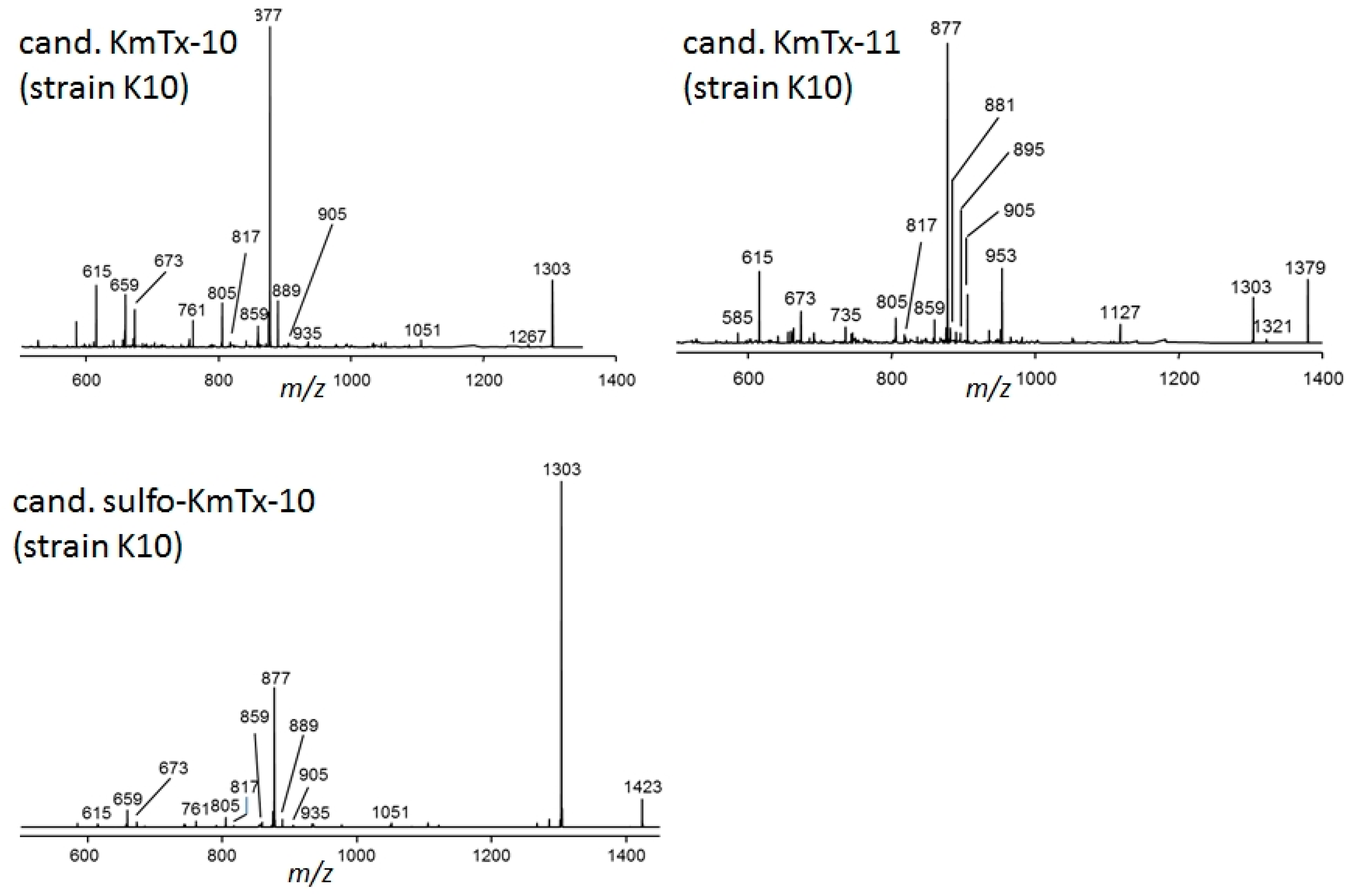
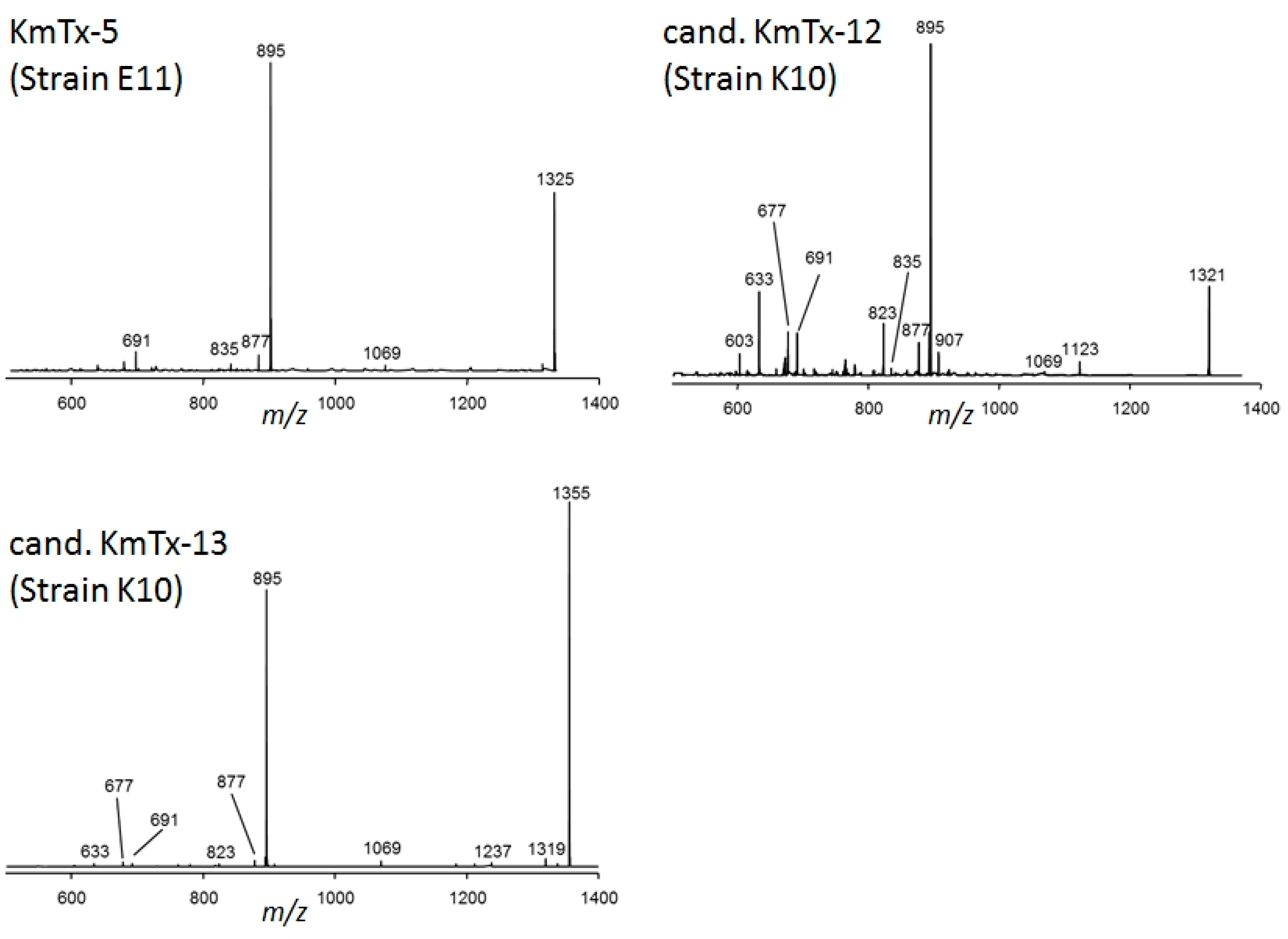
2.2. New Karlotoxin Candidates
2.3. Karlotoxin Profiles
2.4. Collision Induced Dissociation (CID) Spectra of Karlotoxins and Amphidinol-18 (AM-18)
2.5. Selected Reaction Monitoring (SRM) Method for the Detection of Karlotoxins
2.6. Karlotoxin Cell Quota Estimations
2.7. Hemolytic Activity of Cand. Sulfo-KmTx-10
3. Discussion
3.1. Liquid Chromatography-Tandem Mass Spectrometry LC-MS/MS Method
3.2. Quantitation and Method Potential and Limitation
3.3. Characteristics of New Karlotoxins
3.4. Structural Variations of Karlotoxins, Similarities to Amphidinols and Function
4. Materials and Methods
4.1. Isolation and Culturing of Karlodinium Strains
4.2. Molecular Techniques
4.2.1. DNA Extraction for Karlodinium Species Assignment
4.2.2. Polymerase Chain Reaction Template Amplification of Karlodinium spp. from the Mediterranean
4.2.3. Amplicon Preparation and Sequencing
4.2.4. DNA Sequence Analysis and Molecular Taxonomic Assignment
4.3. Chemical Analysis
4.3.1. Harvest of Karlodinium Cells and Extraction of Karlotoxins
4.3.2. Solid Phase Extraction (SPE) Clean-Up of Karlodinium Extracts
4.3.3. LC-MS/MS Precursor Ion Spectra
4.3.4. LC-MS/MS Collision Induced Dissociation (CID) Spectra
4.3.5. LC-MS/MS Selected Reaction Monitoring (SRM)
4.3.6. High Resolution Mass Spectroscopy
4.4. Hemolytic Assay
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Disclaimer
References
- Abbott, B.C.; Ballantine, D. The toxin from Gymnodinium veneficum Ballantine. J. Mar. Biol. Assoc. U. K. 1957, 36, 169–189. [Google Scholar] [CrossRef]
- Place, A.R.; Bowers, H.A.; Bachvaroff, T.R.; Adolf, J.E.; Deeds, J.R.; Sheng, J. Karlodinium veneficum—The little dinoflagellate with a big bite. Harmful Algae 2012, 14, 179–195. [Google Scholar] [CrossRef]
- Fernandez-Tejedor, M.; Soubrier-Pedreno, M.A.; Furones, M.D. Mitigation of lethal effects of Karlodinium veneficum and K. armiger on Sparus aurata: Changes in haematocrit and plasma osmolality. Dis. Aquat. Org. 2007, 77, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Bachvaroff, T.R.; Adolf, J.E.; Squier, A.H.; Harvey, H.R.; Place, A.R. Characterization and quantification of karlotoxins by liquid chromatography-mass spectrometry. Harmful Algae 2008, 7, 473–484. [Google Scholar] [CrossRef]
- Deeds, J.R.; Terlizzi, D.E.; Adolf, J.E.; Stoecker, D.K.; Place, A.R. Toxic activity from cultures of Karlodinium micrum (= Gyrodinium galatheanum) (Dinophyceae)—A dinoflagellate associated with fish mortalities in an estuarine aquaculture facility. Harmful Algae 2002, 1, 169–189. [Google Scholar] [CrossRef]
- Kempton, J.W.; Lewitus, A.J.; Deeds, J.R.; Law, J.M.; Place, A.R. Toxicity of Karlodinium micrum (Dinophyceae) associated with a fish kill in a South Carolina brackish retention pond. Harmful Algae 2002, 1, 233–241. [Google Scholar] [CrossRef]
- Adolf, J.E.; Bachvaroff, T.R.; Place, A.R. Can cryptophyte abundance trigger toxic Karlodinium veneficum blooms in eutrophic estuaries? Harmful Algae 2008, 8, 119–128. [Google Scholar] [CrossRef]
- Rasmussen, S.A.; Binzer, S.B.; Hoeck, K.; Meier, S.; De Medeiros, L.S.; Andersen, N.G.; Place, A.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Karmitoxin: An Amine-Containing Polyhydroxy-Polyene Toxin from the Marine Dinoflagellate Karlodinium armiger. J. Nat. Prod. 2017, 80, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Malkiel, E.; Katz, J.; Adolf, J.E.; Place, A.R. A dinoflagellate exploits toxins to immobilize prey prior to ingestion. Proc. Natl. Acad. Sci. USA 2010, 107, 2082–2087. [Google Scholar] [CrossRef] [PubMed]
- Bachvaroff, T.R.; Adolf, J.E.; Place, A.R. Strain Variation in Karlodinium veneficum (Dinophyceae): Toxin Profiles, Pigments, and Growth Characteristics. J. Phycol. 2009, 45, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Adolf, J.E.; Krupatkina, D.; Bachvaroff, T.; Place, A.R. Karlotoxin mediates grazing by Oxyrrhis marina on strains of Karlodinium veneficum. Harmful Algae 2007, 6, 400–412. [Google Scholar] [CrossRef]
- Waggett, R.J.; Tester, P.A.; Place, A.R. Anti-grazing properties of the toxic dinoflagellate Karlodinium veneficum during predator-prey interactions with the copepod Acartia tonsa. MEPS 2008, 366, 31–42. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Deeds, J.R.; Satake, M.; Ribeiro, A.A.; Place, A.R.; Wright, J.L.C. Isolation and characterization of karlotoxin 1, a new amphipathic toxin from Karlodinium veneficum. Tetrahedron Lett. 2008, 49, 6457–6461. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, R.M.; Deeds, J.R.; Tatters, A.O.; Place, A.R.; Tomas, C.R.; Wright, J.L.C. Structure and Relative Potency of Several Karlotoxins from Karlodinium veneficum. J. Nat. Prod. 2010, 73, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Place, A.R.; Yoshida, W.Y.; Anklin, C.; Hamann, M.T. Structure and Absolute Configuration of Karlotoxin-2, an Ichthyotoxin from the Marine Dinoflagellate Karlodinium veneficum. J. Am. Chem. Soc. 2010, 132, 3277–3279. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Place, A.R. Sterol-specific membrane interactions with the toxins from Karlodinium micrum (Dinophyceae)—A strategy for self-protection? Afr. J. Mar. Sci. 2006, 28, 421–425. [Google Scholar] [CrossRef]
- Busch, J.A.; Andree, K.B.; Diogène, J.; Fernández-Tejedor, M.; Toebe, K.; John, U.; Krock, B.; Tillmann, U.; Cembella, A.D. Toxigenic algae and associated phycotoxins in two coastal embayments in the Ebro Delta (NW Mediterranean). Harmful Algae 2016, 55, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busch, J.A.; Cembella, A.D.; Fernández-Tejedor, M.; Diogène, J.; Zielinski, O. An integrated approach for the assessment of HAB dynamics in two NW Mediterranean bays from a GEOHAB perspective. In Proceedings of the 14th International Conference on Harmful Algae, Hersonissos, Crete, Greek, 1–5 November 2010; International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2012; pp. 84–86. [Google Scholar]
- Fernández-Tejedor, M.; Soubrier-Pedreño, M.Á.; Furones, M.D. Acute LD50 of a Gyrodinium corsicum natural population for Sparus aurata and Dicentrarchus labrax. Harmful Algae 2004, 3, 1–9. [Google Scholar] [CrossRef]
- Fernández-Tejedor, M.; Delgado, M.; Garcés, E.; Camp, J.; Diogène, J. Toxic phytoplankton response to warming in two Mediterranean bays of the Ebro Delta. In Phytoplankton Response to Mediterranean Environmental Changes; Briand, F., Ed.; CIESM Publisher: Monaco, 2010; pp. 83–86. [Google Scholar]
- MacKenzie, L.; Beuzenberg, V.; Holland, P.; McNabb, P.; Selwood, A. Solid phase adsorption toxin tracking (SPATT): A new monitoring tool that simulates the biotoxin contamination of filter feeding bivalves. Toxicon 2004, 44, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, L.A. In situ passive solid-phase adsorption of micro-algal biotoxins as a monitoring tool. Curr. Opin. Biotechnol. 2010, 21, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Curtis, J.M.; Walter, J.A.; Wright, J.L.C. Identification of DTX-4, a new water-soluble phosphatase inhibitor from the toxic dinoflagellate Prorocentrum lima. J. Chem. Soc. Chem. Commun. 1995, 0, 597–599. [Google Scholar] [CrossRef]
- Hu, T.; LeBlanc, P.; Burton, I.W.; Walter, J.A.; McCarron, P.; Melanson, J.E.; Strangman, W.K.; Wright, J.L.C. Sulfated diesters of okadaic acid and DTX-1: Self-protective precursors of diarrhetic shellfish poisoning (DSP) toxins. Harmful Algae, 2017, 63, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Gómez, B.; Souto, M.L.; Norte, M.; Fernández, J.J. Isolation and structural determination of DTX-6, a new okadaic acid derivative. J. Nat. Prod. 2001, 64, 1363–1364. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, G.; Cutignano, A.; Sardo, A.; Fontana, A. Antifungal amphidinol-18 and its 7-sulfate derivative from the marine dinoflagellate Amphidinium carterae. J. Nat. Prod. 2014, 77, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.O.; Samdal, I.A.; Aasen, J.A.B.; Jensen, D.J.; Quilliam, M.A.; Petersen, D.; Briggs, L.M.; Wilkins, A.L.; Rise, F.; Cooney, J.M.; et al. Evidence for numerous analogs of yessotoxin in Protoceratium reticulatum. Harmful Algae 2005, 4, 1075–1091. [Google Scholar] [CrossRef]
- Martens, H.; Tillmann, U.; Harju, K.; Dell’Aversano, C.; Tartaglione, L.; Krock, B. Toxin Variability Estimations of 68 Alexandrium ostenfeldii (Dinophyceae) Strains from The Netherlands Reveal a Novel Abundant Gymnodimine. Microorganisms 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; McCarrron, P.; Krock, B.; Kilcoyne, J.; Miles, C.O. Azaspiracids: Chemistry, biosynthesis, metabolism, and detection. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection, 3rd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 799–821. [Google Scholar]
- Deeds, J.R.; Hoesch, R.E.; Place, A.R.; Kao, J.P.Y. The cytotoxic mechanism of karlotoxin 2 (KmTx 2) from Karlodinium veneficum (Dinophyceae). Aquat. Toxicol. 2015, 159, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Adolf, J.E.; Bachvaroff, T.R.; Bowers, H.A.; Deeds, J.R.; Place, A.R. Ichthyotoxic Karlodinium veneficum in the Upper Swan River estuary (Western Australia): Synergistic effects of karlotoxin and hypoxia leading to a fish kill. Harmful Algae 2015, 48, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Alpermann, T.; John, U.; Cembella, A. Allelochemical interactions and short-term effects of the dinoflagellate Alexandrium on selected photoautotrophic and heterotrophic protists. Harmful Algae 2008, 7, 52–64. [Google Scholar] [CrossRef]
- Tillmann, U.; Hansen, P.J. Allelopathic effects of Alexandrium tamarense on other algae: Evidence from mixed growth experiments. Aquat. Microb. Ecol. 2009, 57, 101–112. [Google Scholar] [CrossRef]
- Ma, H.; Krock, B.; Tillmann, U.; Bickmeyer, U.; Graeve, M.; Cembella, A.D. Mode of action of membrane-disruptive lytic compounds from the marine dinoflagellate Alexandrium tamarense. Toxicon 2011, 58, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Krock, B.; Tillmann, U.; Muck, A.; Wielsch, N.; Svatoš, A.; Cembella, A.D. Isolation of activity and partial characterization of large non-proteinaceous lytic allelochemicals produced by the marine dinoflagellate Alexandrium tamarense. Harmful Algae 2011, 11, 65–72. [Google Scholar] [CrossRef]
- Place, A.R.; Harvey, H.R.; Bai, X.; Coats, D.W. Sneaking under the toxin surveillance radar: Parasitism and Sterol Content. Afr. J. Mar. Sci. 2006, 28, 347–353. [Google Scholar] [CrossRef]
- Calbet, A.; Bertos, M.; Fuentes-Grünewald, C.; Alacid, E.; Figeroa, R.; Renom, B.; Garcés, E. Intraspecific variability in Karlodinium veneficum: Growth rates, mixotrophy, and lipid composition. Harmful Algae 2010, 10, 654–667. [Google Scholar] [CrossRef]
- Tillmann, U.; Alpermann, T.J.; da Purificação, R.C.; Krock, B.; Cembella, A.D. Intra-population clonal variability in allelochemical potency of the toxigenic dinoflagellate Alexandrium tamarense. Harmful Algae 2009, 8, 759–769. [Google Scholar] [CrossRef]
- West, J.A.; McBride, D.L. Long-term and diurnal carpospore discharge patterns in the Ceramiaceae, Rhodomelaceae and Delesseriaceae (Rhodophyta). Hydrobiologia 1999, 398, 101–114. [Google Scholar] [CrossRef]
- Andree, K.B.; Quijano-Scheggia, S.; Fernández, M.; Elandaloussi, L.M.; Garcés, E.; Camp, J.; Diogène, J. Quantitative PCR Coupled with Melt Curve Analysis for Detection of Selected Pseudonitzschia spp. (Bacillariophyceae) from the Northwestern Mediterranean Sea. Environ. Microbiol. 2011, 77, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Scholin, C.A.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). I. RFLP analysis of SSU rRNA genes. J. Phycol. 1994, 30, 744–754. [Google Scholar] [CrossRef]
- Toldrà, A.; Andree, K.B.; Fernández-Tejedor, M.; Diogène, J.; Campàs, M. Dual quantitative PCR assay for identification and enumeration of Karlodinium veneficum and Karlodinium armiger combined with a simple and rapid DNA extraction method. J. Appl. Phycol. Submitted.
- Boldin, I.A.; Nikolaev, E.N. Fourier transform ion cyclotron resonance cell with dynamic harmonization of the electric field in the whole volume by shaping of the excitation and detection electrode assembly. Rapid Commun. Mass Spectrom. 2011, 25, 122–126. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain # | Harvested Cells | Species Identification | Geographical Origin |
|---|---|---|---|
| E11 | 5.10 × 106 | K. veneficum | Fangar Bay, Ebro Delta (2012) |
| K10 | 1.79 × 108 | K. veneficum | Alfacs Bay, Ebro Delta (2007) |
| CCMP2936 | 1.28 × 108 | K. veneficum | Swann Keys, DE, USA (2006) |
| CCMP415 | 1.67 × 109 | K. veneficum | Norway (1985) |
| IRTA-SMM-12-23 | 6.35 × 107 | K. veneficum | Alfacs Bay, Ebro Delta (2012) |
| IRTA-SMM-12-01 | 8.36 × 108 | K. veneficum | Alfacs Bay, Ebro Delta (2012) |
| K0668 | 2.37 × 108 | K. veneficum | Alfacs Bay, Ebro Delta (2002) |
| IRTA-SMM-12-17 | 1.20 × 108 | K. veneficum | Alfacs Bay, Ebro Delta (2012) |
| IRTA-SMM-12-03 | 6.63 × 108 | K. veneficum | Alfacs Bay, Ebro Delta (2012) |
| IRTA-SMM-12-07 | 1.35 × 108 | K. veneficum | Alfacs Bay, Ebro Delta (2012) |
| K. armiger 1 * | 6.24 × 107 | K. armiger | Alfacs Bay, Ebro Delta (2000) |
| K. armiger 2 * | 4.45 × 107 | K. armiger | Alfacs Bay, Ebro Delta (2000) |
| Toxin | m/z Observed | Elemental Composition | ±ppm |
|---|---|---|---|
| cand. KmTx-10 | 1303.733892 | C62H117ClNaO22S | 0.08 |
| cand. KmTx-11 | 1379.748807 | C64H121ClNaO25S | 0.8 |
| cand. sulfo-KmTx-10 | 1423.682504 | C62H118ClNa2O26S2 | 0.4 |
| cand. KmTx-12 | 1321.742175 | C62H119ClNaO23S | 1.0 |
| cand. KmTx-13 | 1355.749870 | C62H121ClNaO25S | 0.2 |
| Strain | KmTx-1 | KmTx-3 | KmTx-5 | Cand. KmTx-10 | Cand. KmTx-12 | Cand. KmTx-13 | Cand. KmTx-11 | Cand. Sulfo-KmTx-10 |
|---|---|---|---|---|---|---|---|---|
| E11 | - | - | 0.21 | - | - | - | - | - |
| K10 | - | - | - | 0.03 | 0.58 | 0.002 | 0.003 | 0.07 |
| CCMP2936 | 7.9 | 2.5 | - | - | - | - | - | - |
| CCMP415 | - | - | - | - | - | - | - | - |
| IRTA-SMM-12-23 | - | - | 0.05 | 0.08 | 1.2 | 0.10 | 0.02 | 0.5 |
| IRTA-SMM-12-01 | - | - | - | - | - | - | - | - |
| K0668 | - | - | 0.004 | 0.006 | 0.16 | 0.003 | 0.009 | 0.09 |
| IRTA-SMM-12-17 | - | - | 0.08 | 0.09 | 2.2 | 0.40 | 0.15 | 0.64 |
| IRTA-SMM-12-03 | - | - | - | - | - | - | - | - |
| IRTA-SMM-12-07 | - | - | 0.57 | 0.006 | 0.28 | 0.01 | 0.02 | 0.03 |
| K. armiger 1 | - | - | - | - | - | - | - | - |
| K. armiger 2 | - | - | - | - | - | - | - | - |
| Toxin | Q1 Mass (m/z) | Q3 Mass (m/z) | Retention Time (min) | Molecular Weight (g mol−1) |
|---|---|---|---|---|
| AM-18 | 1381.8 1381.8 | 1105.6 687.6 | 9.9 | 1358.8 |
| KcTx-1 | 1339.8 1339.8 | 1121.7 719.6 | 9.7 | 1316.8 |
| KmTx-1 | 1361.8 1361.8 | 937.6 877.6 | 10.2 | 1338.8 |
| KmTx-2 | 1367.8 1367.8 | 937.6 877.6 | 10.0 | 1344.8 |
| 44-oxo-KmTx-2 | 1383.8 1383.8 | 937.6 877.6 | 10.0 | 1360.8 |
| KmTx-3 | 1347.8 1347.8 | 937.6 877.6 | 10.3 | 1324.8 |
| KmTx-5 | 1325.8 1325.8 | 895.6 691.5 | 10.1 | 1302.8 |
| cand. KmTx-10 | 1303.8 1303.8 | 877.6 615.4 | 9.6 | 1280.7 |
| cand. KmTx-12 | 1321.8 1321.8 | 895.6 633.4 | 9.8 | 1298.7 |
| cand. KmTx-13 | 1355.8 1355.8 | 895.7 677.5 | 9.1 | 1332.7 |
| cand. KmTx-11 | 1379.8 1379.8 | 877.6 615.4 | 9.9 | 1356.7 |
| cand. sulfo-KmTx-10 | 1423.8 1423.8 | 1303.8 877.6 | 9.8 | 1400.7 |
| Parameter | Precursor Ion Scan | Collision Induced Dissociation (CID) | Selected Reaction Monitoring (SRM) |
|---|---|---|---|
| Curtain gas | 20 psi | 20 psi | 20 psi |
| Collision activated dissociation (CAD) | high | high | high |
| Ion spray voltage | 5500 V | 5500 V | 5500 V |
| Temperature | 550 °C | 550 °C | 550 °C |
| Nebulizer gas | 30 psi | 30 psi | 30 psi |
| Auxiliary gas | 60 psi | 60 psi | 60 psi |
| Interface heater | on | on | on |
| Declustering potential | 151 V | 151 V | 151 V |
| Entrance potential | 10 V | - | 10 V |
| Collision energy | 80 V | 100 V | 100 V |
| Collision energy spread | - | 10 V | - |
| Exit potential | 26 V | - | 26 V |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krock, B.; Busch, J.A.; Tillmann, U.; García-Camacho, F.; Sánchez-Mirón, A.; Gallardo-Rodríguez, J.J.; López-Rosales, L.; Andree, K.B.; Fernández-Tejedor, M.; Witt, M.; et al. LC-MS/MS Detection of Karlotoxins Reveals New Variants in Strains of the Marine Dinoflagellate Karlodinium veneficum from the Ebro Delta (NW Mediterranean). Mar. Drugs 2017, 15, 391. https://doi.org/10.3390/md15120391
Krock B, Busch JA, Tillmann U, García-Camacho F, Sánchez-Mirón A, Gallardo-Rodríguez JJ, López-Rosales L, Andree KB, Fernández-Tejedor M, Witt M, et al. LC-MS/MS Detection of Karlotoxins Reveals New Variants in Strains of the Marine Dinoflagellate Karlodinium veneficum from the Ebro Delta (NW Mediterranean). Marine Drugs. 2017; 15(12):391. https://doi.org/10.3390/md15120391
Chicago/Turabian StyleKrock, Bernd, Julia A. Busch, Urban Tillmann, Francisco García-Camacho, Asterio Sánchez-Mirón, Juan J. Gallardo-Rodríguez, Lorenzo López-Rosales, Karl B. Andree, Margarita Fernández-Tejedor, Matthias Witt, and et al. 2017. "LC-MS/MS Detection of Karlotoxins Reveals New Variants in Strains of the Marine Dinoflagellate Karlodinium veneficum from the Ebro Delta (NW Mediterranean)" Marine Drugs 15, no. 12: 391. https://doi.org/10.3390/md15120391
APA StyleKrock, B., Busch, J. A., Tillmann, U., García-Camacho, F., Sánchez-Mirón, A., Gallardo-Rodríguez, J. J., López-Rosales, L., Andree, K. B., Fernández-Tejedor, M., Witt, M., Cembella, A. D., & Place, A. R. (2017). LC-MS/MS Detection of Karlotoxins Reveals New Variants in Strains of the Marine Dinoflagellate Karlodinium veneficum from the Ebro Delta (NW Mediterranean). Marine Drugs, 15(12), 391. https://doi.org/10.3390/md15120391