Physiological and Biochemical Changes Reveal Differential Patterns of Docosahexaenoic Acid Partitioning in Two Marine Algal Strains of Isochrysis



Abstract
:1. Introduction
2. Results and Discussion
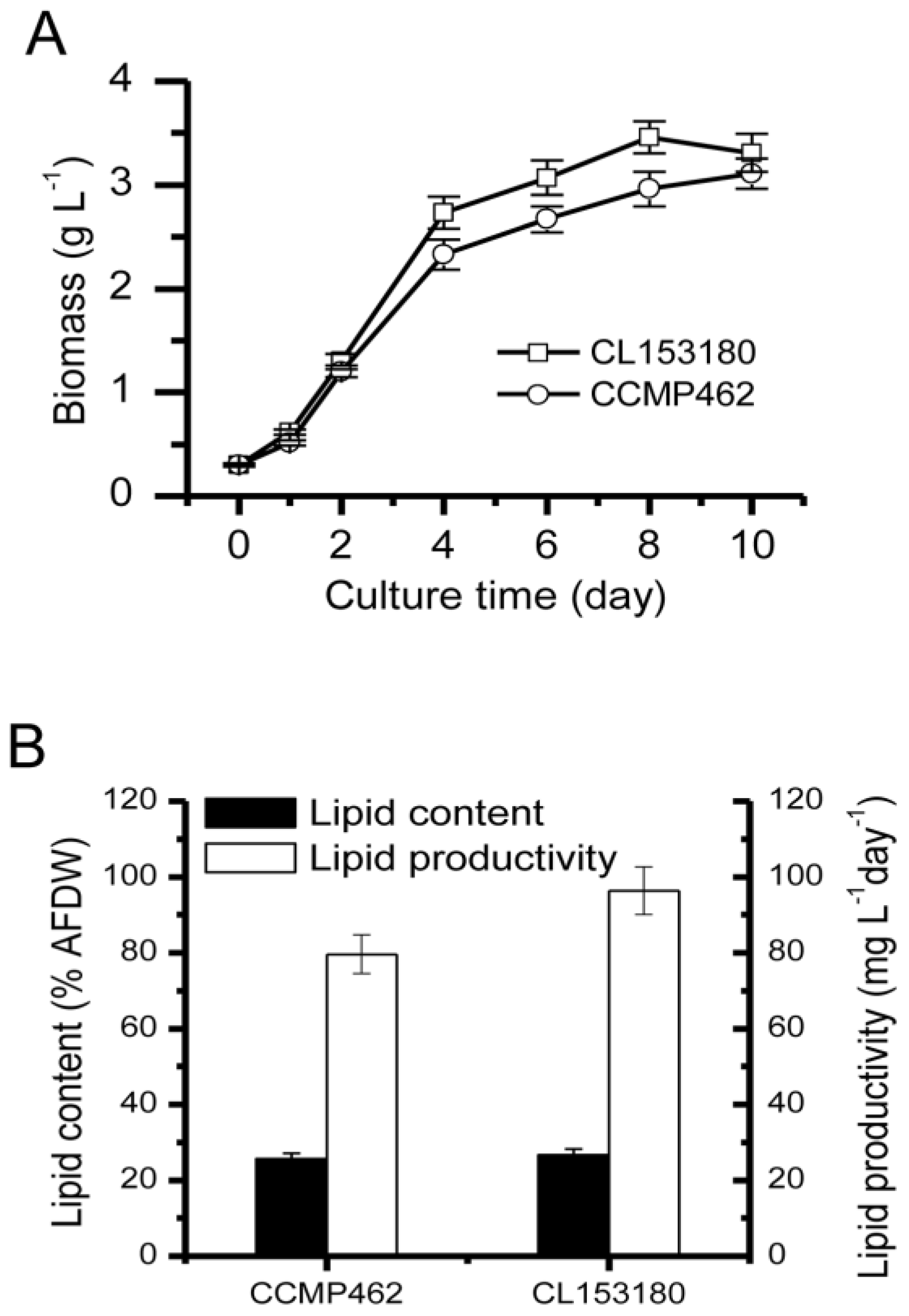
2.1. Biomass and Lipid Accumulation of Two Microalgae
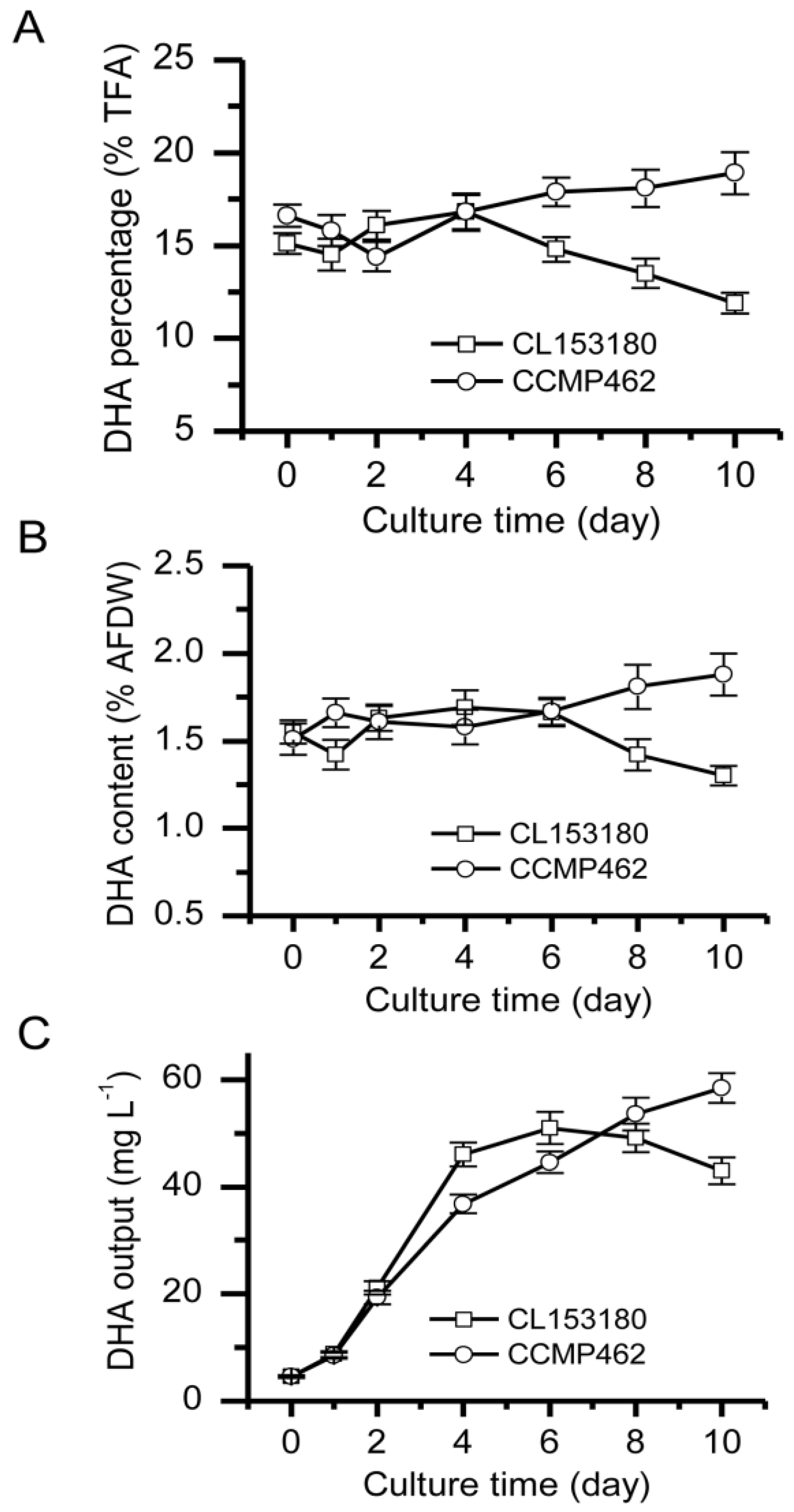
2.2. Batch Culture for DHA Production
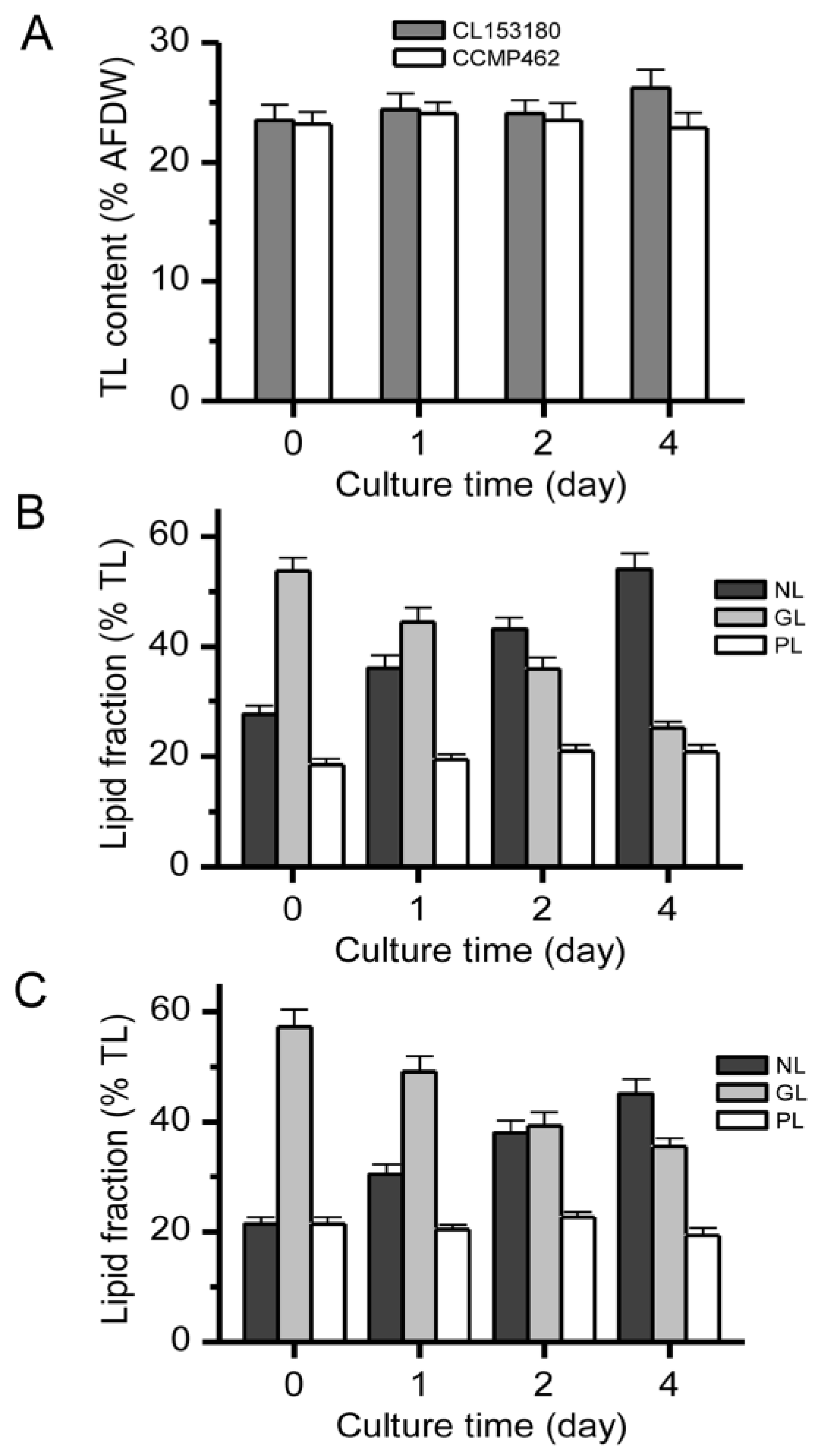
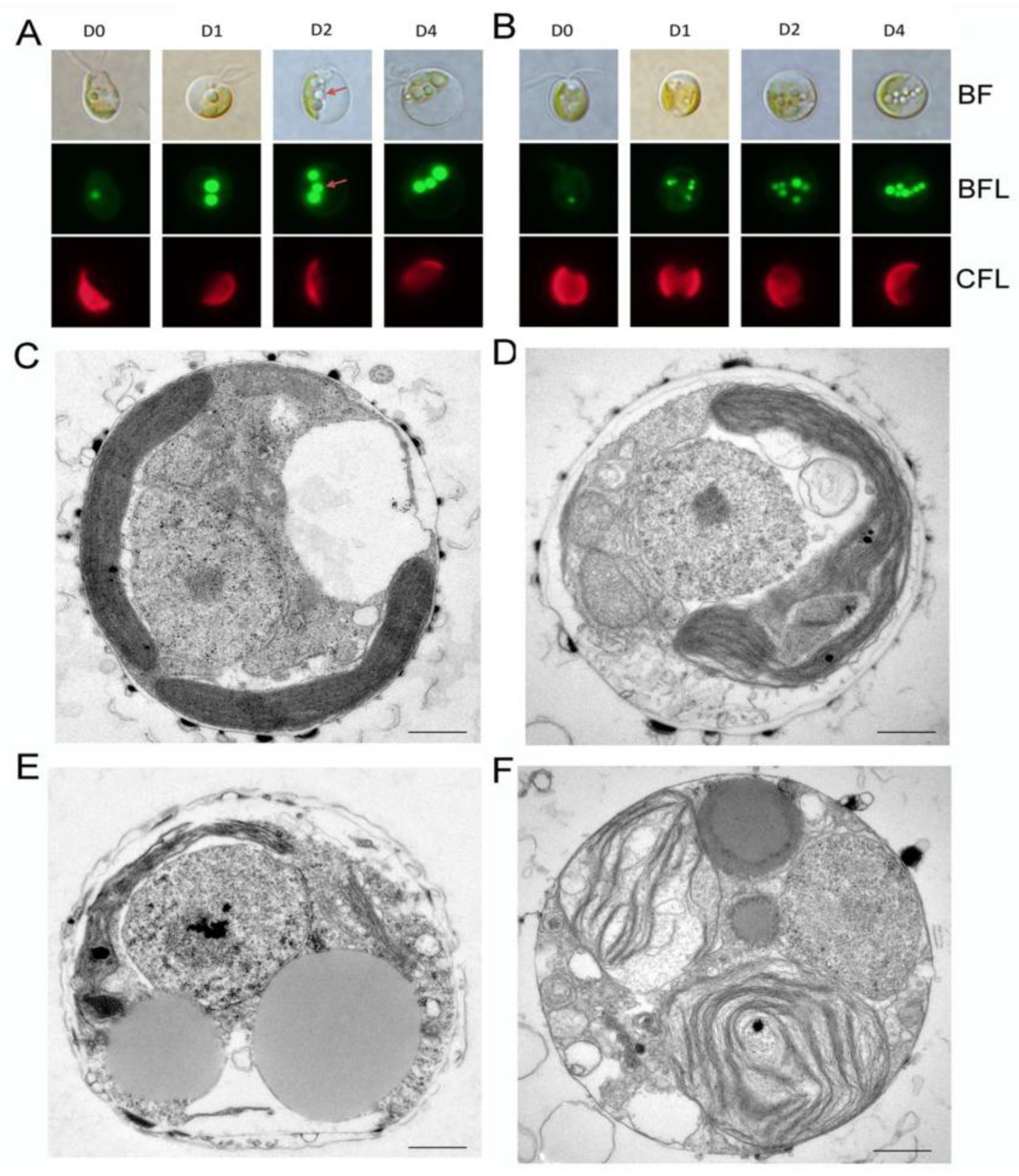
2.3. Impact of ND on Lipid Compositions and Cell Structures
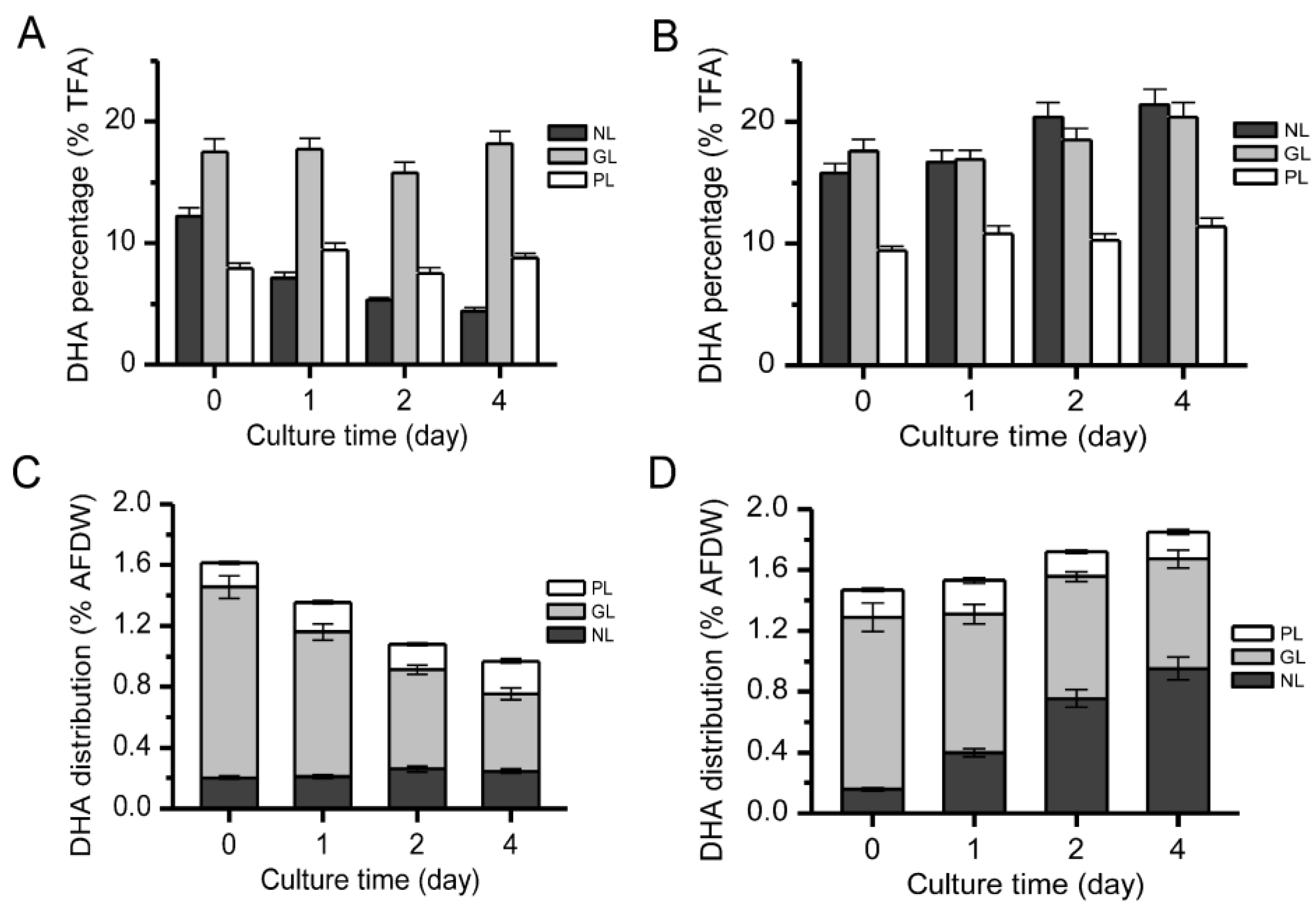
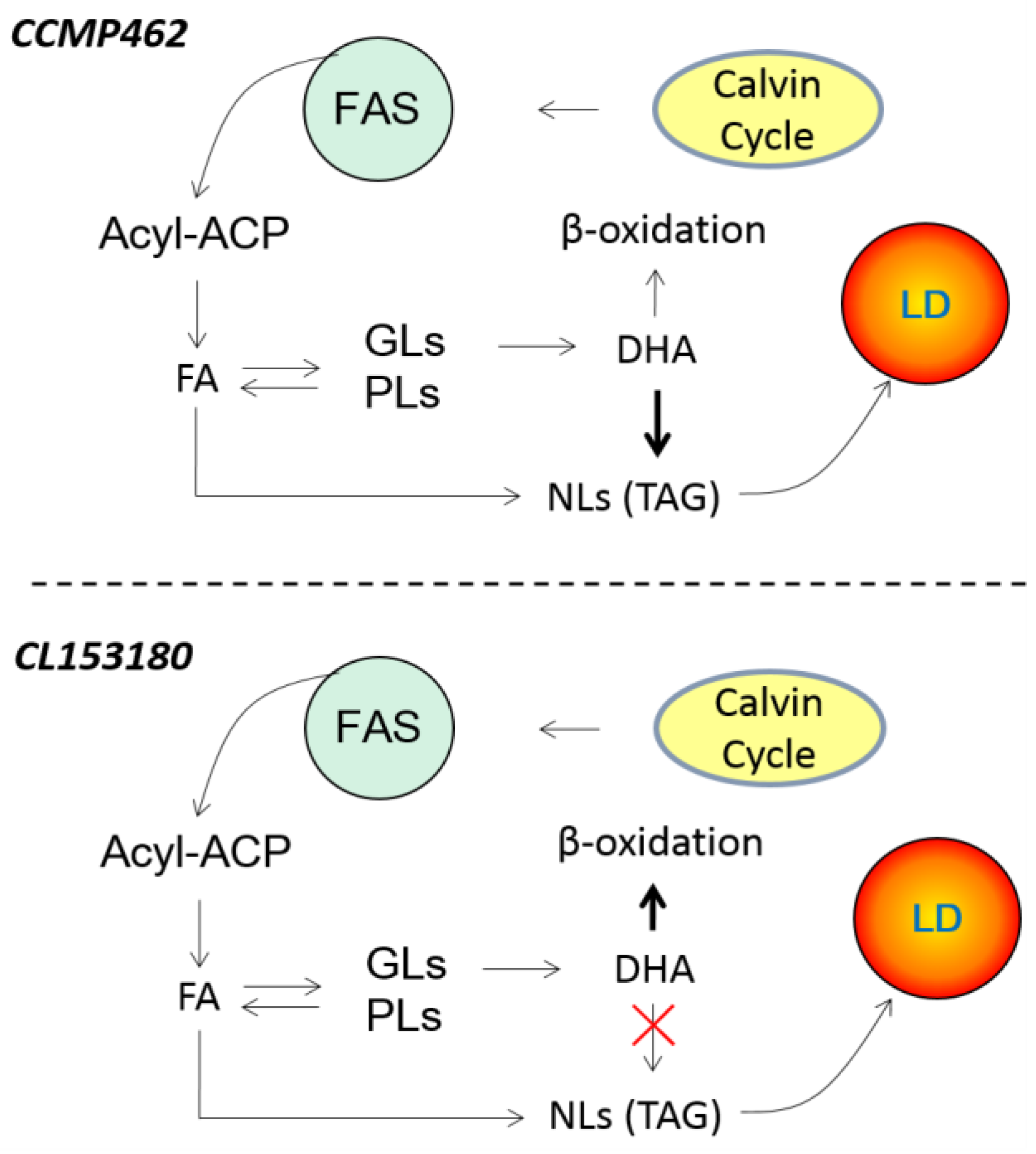
2.4. Distribution of DHA in Different Lipid Components
2.5. Effects of ND on Fatty Acid Profiles
3. Materials and Methods
3.1. Algal Strains and Maintenance
3.2. Algal Cultivation
3.3. Growth Measurements
3.4. Lipid Extraction and Analysis
3.5. Bodipy Staining
3.6. Ultrastructural Observation
3.7. Fatty Acid Analysis
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Das, U.N.; Fams, M.D. Long-chain polyunsaturated fatty acids in the growth and development of the brain and memory. Nutrition 2003, 19, 62–65. [Google Scholar] [CrossRef]
- Bakker, E.C.; Hornstra, G.; Blanco, C.E.; Vles, J.S.H. Relationship between long-chain polyunsaturated fatty acids at birth and motor function at 7 years of age. Eur. J. Clin. Nutr. 2009, 63, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Guesnet, P.; Alessandri, J.M. Docosahexaenoic acid (DHA) and the developing central nervous system (CNS)-implications for dietary recommendations. Biochimie 2011, 93, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, L.; Hansen, H.S.; Jorgensen, M.H.; Michaelsen, K.F. The essentiality of long chain omega-3 fatty acids in relation to development and function of the brain and retina. Prog. Lipid Res. 2001, 40, 1–94. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology. Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Fan, K.W.; Chen, F.; Jones, E.B.G.; Vrijmoed, L.L.P. Eicosapentaenoic and docosahexaenoic acids production by and okara-utilizing potential of Thraustochytrids. J. Ind. Microbiol. Biotechnol. 2001, 27, 199–202. [Google Scholar] [CrossRef]
- Ethier, S.; Woisard, K.; Vaughan, D.; Wen, Z. Continuous culture of the microalgae Schizochytrium limacinum on biodiesel-derived crude glycerol for producing docosahexaenoic acid. Bioresour. Technol. 2011, 102, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.K.; Rairakhwada, D.; Seo, P.S.; Park, S.Y.; Hur, B.K.; Kim, C.H.; Seo, J.W. Production of lipids containing high levels of docosahexaenoic acid by a newly isolated microalga, Aurantiochytrium sp. KRS101. Appl. Biochem. Biotechnol. 2011, 164, 1468–1480. [Google Scholar] [CrossRef] [PubMed]
- Pleissner, D.; Eriksen, N.T. Effects of phosphorous, nitrogen, and carbon limitation on biomass composition in batch and continuous flow cultures of the heterotrophic dinoflagellate Crypthecodinium cohnii. Biotechnol. Bioeng. 2012, 109, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.L.; Reis, A. The use of multi-parameter flow cytometry to study the impact of n-dodecane additions to marine dinoflagellate microalga Crypthecodinium cohnii batch fermentations and DHA production. J. Ind. Microbiol. Biotechnol. 2008, 35, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, F. Effects of medium glucose concentration and pH on docosahexaenoic acid content of heterotrophic Crypthecodinium cohnii. Process Biochem. 2000, 35, 1205–1209. [Google Scholar] [CrossRef]
- Liu, J.; Sommerfeld, M.; Hu, Q. Screening and characterization of Isochrysis strains and optimization of culture conditions for docosahexaenoic acid production. Appl. Microbiol. Biotechnol. 2013, 10, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Devos, M.; Poisson, L.; Ergan, F.; Pencreac’h, G. Enzymatic hydrolysis of phospholipids from Isochrysis galbana for docosahexaenoic acid enrichment. Enzyme Microb. Technol. 2006, 39, 548–554. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chang, F.L.; Tsao, C.Y.; Leu, J.Y. Influence of growth phase and nutrient source on fatty acid composition of Isochrysis galbana CCMP 1324 in a batch photoreactor. Biol. Eng. J. 2007, 37, 166–176. [Google Scholar] [CrossRef]
- Cai, S.Q.; Hu, C.Q.; Du, S.B. Comparisons of growth and biochemical composition between mixed culture of alga and yeast and monocultures. J. Biosci. Bioeng. 2007, 5, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Yago, T.; Yoshie-Stark, Y.; Arakawa, H.; Morinaga, T. Effect of high frequency of intermittent light on the growth and fatty acid profile of Isochrysis galbana. Aquaculture 2012, 338, 111–117. [Google Scholar] [CrossRef]
- Balduyck, L.; Bijttebier, S.; Bruneel, C.; Jacobs, G.; Voorspoels, S.; Durme, J.V. Lipolysis in T-Isochrysis lutea during wet storage at different temperatures. Algal Res. 2016, 18, 281–287. [Google Scholar] [CrossRef]
- Huerlimann, R.; Nys, R.; Heimann, K. Growth, lipid content, productivity, and fatty acid composition of tropical microalgae for scale-up production. Biotechnol. Bioeng. 2010, 10, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.J.; Lee, Y.K.; Chao, T.M. Effects of temperature and growth phase on lipid and biochemical composition of Isochrysis galbana TK1. J. Appl. Phycol. 1997, 9, 451–457. [Google Scholar] [CrossRef]
- Freddy, G.; Dagmar, B.S. Interactive effects of light and temperature on pigments and omega-3 LC-PUFA-enriched oil accumulation in batch-cultivated Pavlova lutheri using high-bicarbonate supply. Algal Res. 2017, 23, 113–125. [Google Scholar]
- Simionato, D.; Block, M.A.; Rocca, N.L. The response of Nannochloropsis gaditana to nitrogen starvation includes de novo biosynthesis of triacylglycerols, a decrease of chloroplast galactolipids, and reorganization of the photosynthetic apparatus. Eukaryot. Cell 2013, 12, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Berges, J.A.; Charlebois, D.O.; Mauzerall, D.C.; Falkowski, P.G. Differential effects of nitrogen limitation on photosynthetic efficiency of photosystems I and II in microalgae. Plant Physiol. 1996, 110, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.; Berg, R.B.; Schachtman, D.P. Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Molina, G.E.; Perez, J.A.S.; Camacho, F.G.; Sevilla, J.M.F.; Fernández, F.G.A. Effect of growth rate on the eicosapentaenoic acid and docosahexaenoic acid content of Isochrysis galbana in chemostat culture. Appl. Microbiol. Biotechnol. 1994, 41, 23–27. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B. Acyl-lipid metabolism. In The Arabidopsis Book; Beisson, F., Andersson, M.X., Arondel, V., Bates, P.D., Baud, S., Bird, D., Bono, A.D., Durrett, T.P., Franke, R.B., Graham, I.A., et al., Eds.; The American Society of Plant Biologists: Washington, DC, USA, 2010; Volume 133, pp. 1–65. ISBN 1543-8120. [Google Scholar]
- Petrie, J.R.; Vanhercke, T.; Shrestha, P.; Tahchy, A.E.; White, A.; Zhou, X.R.; Liu, Q.; Mansour, M.P.; Nichols, P.D.; Singh, S.P. Recruiting a new substrate for triacylglycerol synthesis in plants: The monoacylglycerol acyltransferase pathway. PLoS ONE 2012, 7, e35214. [Google Scholar] [CrossRef]
- Li, X.; Moellering, E.R.; Liu, B.; Johnny, C.; Fedewa, M.; Sears, B.B.; Kuo, M.H.; Benning, C. A galactoglycerolipid lipase is required for triacylglycerol accumulation and survival following nitrogen deprivation in Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2012, 13, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, D.; Yoon, K.; Hu, Q.; Li, Y. Characterization of type 2 diacylglycerol acyltransferases in Chlamydomonas reinhardtii reveals their distinct substrate specificities and functions in triacylglycerol biosynthesis. Plant J. 2016, 86, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Shi, Y.; Ma, X.; Pan, Y.; Hu, H.; Li, Y.; Luo, M.; Gerken, H.; Liu, J. A type-I diacylglycerol acyltransferase modulates triacylglycerol biosynthesis and fatty acid composition in the oleaginous microalga, Nannochloropsis oceanica. Biotechnol. Biofuels 2017, 10, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Lee, Y.Y.; Wei, L.; Jia, J.; Wang, Q.; Wang, D.; Bai, F.; Hu, H.; Hu, Q.; Liu, J.; et al. Producing designer oils in industrial microalgae by rational modulation of co-evolving type-2 diacylglycerol acyltransferases. Mol. Plant. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Yu, S.Y.; Ouyang, L.L.; Zhou, Z.Z. Comparison of increased arachidonic acid content in Myrmecia incisa cultured during the course of nitrogen or phosphorus starvation. J. Fish. China 2011, 1000, 5–11. [Google Scholar]
- Huerlimann, R.; Steinig, E.J.; Loxton, H.; Zenger, K.R.; Jerry, D.R. Effects of growth phase and nitrogen starvation on expression of fatty acid desaturases and fatty acid composition of Isochrysis aff. galbana (TISO). Gene 2014, 545, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.N.; Chen, Z.A.; Xue, S.; Zhang, W. Increased lipid production of the marine oleaginous microalgae Isochrysis zhangjiangensis (Chrysophyta) by nitrogen supplement. Bioresour. Technol. 2011, 102, 6710–6716. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, H.; Li, C.Y.; Ouyang, L.L.; Zhou, Z.G. Characterization of fatty acidd esaturase (FAD) genes in Myrmecia incisa and the effect of nitrogen stravtion on their transcription. J. Fish. Sci. China 2012, 19, 729–740. [Google Scholar]
- Blight, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Liu, J.; Huang, J.; Fan, K.W.; Jiang, Y.; Zhong, Y.; Sun, Z.; Chen, F. Production potential of Chlorella zofingienesis as a feedstock for biodiesel. Bioresour. Technol. 2010, 101, 8658–8663. [Google Scholar] [CrossRef] [PubMed]
- Christie, W.W.B. Lipid Analysis: Isolation, Separation, Identification, and Structural Analysis of Lipids, 3rd ed.; The Oily Press: Bridgwater, UK, 2003. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids (% TFA) | CL153180 | CCMP462 | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 4 | 0 | 1 | 2 | 4 | |
| c14:0 | 19.4 | 17.4 | 16.7 | 16.7 | 18.9 | 15.9 | 14.1 | 15.5 |
| c16:0 | 14.9 | 18.5 | 19.2 | 20.4 | 11.9 | 13.4 | 13.7 | 14.0 |
| c16:1 | 4.2 | 2.2 | 2.2 | 2.5 | 5.1 | 4.0 | 3.7 | 3.5 |
| c18:0 | 1.0 | 0.9 | 0.7 | 0.2 | 0.2 | 0.3 | 0.3 | 0.3 |
| c18:1 | 14.5 | 26.5 | 30.9 | 33.3 | 17.4 | 19.9 | 20.1 | 21.9 |
| c18:2 | 8.2 | 6.5 | 5.3 | 4.8 | 5.1 | 4.9 | 3.7 | 2.0 |
| c18:3 | 9.1 | 6.2 | 4.4 | 3.8 | 9.5 | 7.6 | 5.9 | 4.1 |
| c18:4 | 12.1 | 9.4 | 9.3 | 9.4 | 14.1 | 16.3 | 17.7 | 18.1 |
| c22:0 | 2.4 | 2.6 | 2.3 | 1.0 | 3.3 | 3.0 | 2.9 | 1.6 |
| c22:6 (DHA) | 14.2 | 9.9 | 8.9 | 7.8 | 14.5 | 14.8 | 17.8 | 19.1 |
| MFA 1 | 18.7 | 28.7 | 33.1 | 35.8 | 22.5 | 23.8 | 23.8 | 25.4 |
| PUFA 2 | 43.5 | 31.9 | 28.0 | 25.8 | 43.2 | 43.7 | 45.1 | 43.2 |
| SFA 3 | 37.7 | 39.4 | 38.9 | 38.4 | 34.3 | 32.5 | 31.0 | 31.4 |
| TFA 4 | 11.9 | 12.9 | 12.5 | 13.2 | 10.9 | 11.2 | 10.3 | 10.1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Chen, Y.; Mao, X.; Liu, J. Physiological and Biochemical Changes Reveal Differential Patterns of Docosahexaenoic Acid Partitioning in Two Marine Algal Strains of Isochrysis. Mar. Drugs 2017, 15, 357. https://doi.org/10.3390/md15110357
Sun Z, Chen Y, Mao X, Liu J. Physiological and Biochemical Changes Reveal Differential Patterns of Docosahexaenoic Acid Partitioning in Two Marine Algal Strains of Isochrysis. Marine Drugs. 2017; 15(11):357. https://doi.org/10.3390/md15110357
Chicago/Turabian StyleSun, Zheng, Yong Chen, Xuemei Mao, and Jin Liu. 2017. "Physiological and Biochemical Changes Reveal Differential Patterns of Docosahexaenoic Acid Partitioning in Two Marine Algal Strains of Isochrysis" Marine Drugs 15, no. 11: 357. https://doi.org/10.3390/md15110357
APA StyleSun, Z., Chen, Y., Mao, X., & Liu, J. (2017). Physiological and Biochemical Changes Reveal Differential Patterns of Docosahexaenoic Acid Partitioning in Two Marine Algal Strains of Isochrysis. Marine Drugs, 15(11), 357. https://doi.org/10.3390/md15110357