Proteomic Analysis of the Chlorophyta Dunaliella New Strain AL-1 Revealed Global Changes of Metabolism during High Carotenoid Production



Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of the Newly Isolated Dunaliella Strain
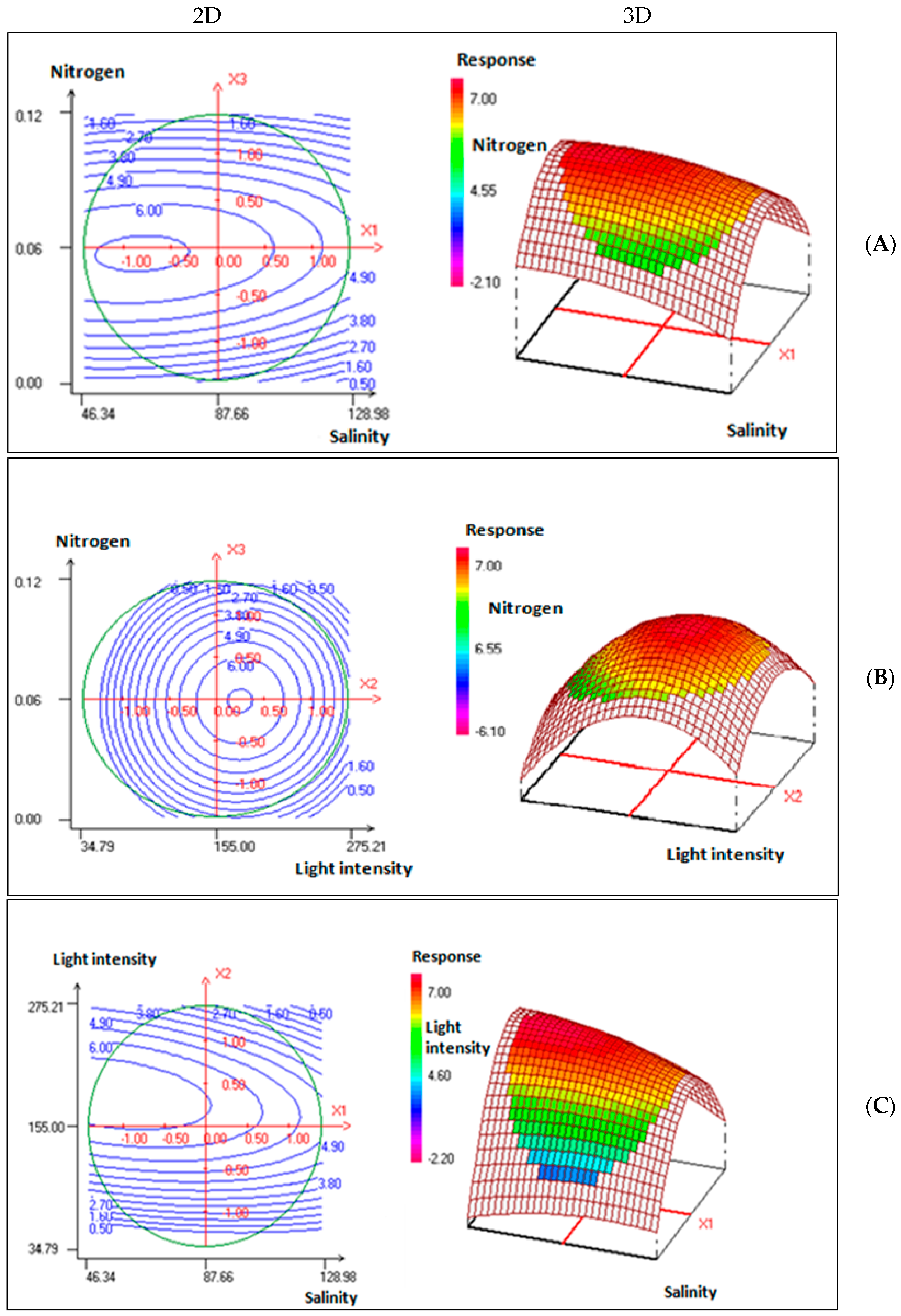
2.2. Experimental Design and Validation of Optimized Growth Conditions
2.3. Proteomic Analysis
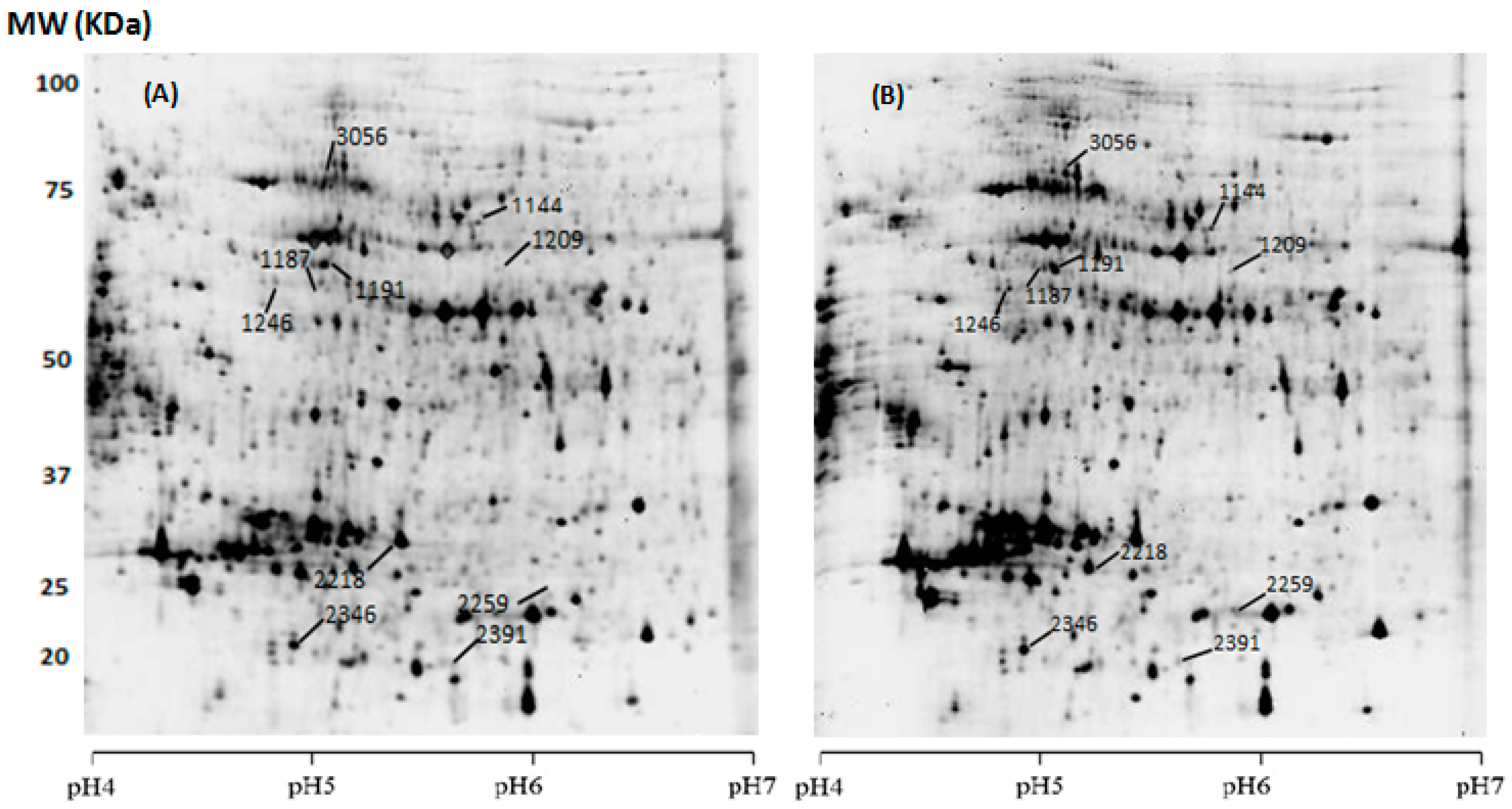
2.3.1. Two-Dimensional Electrophoresis Profiles of Total Proteins in Response to Carotenoid High Production
2.3.2. Spot Identification
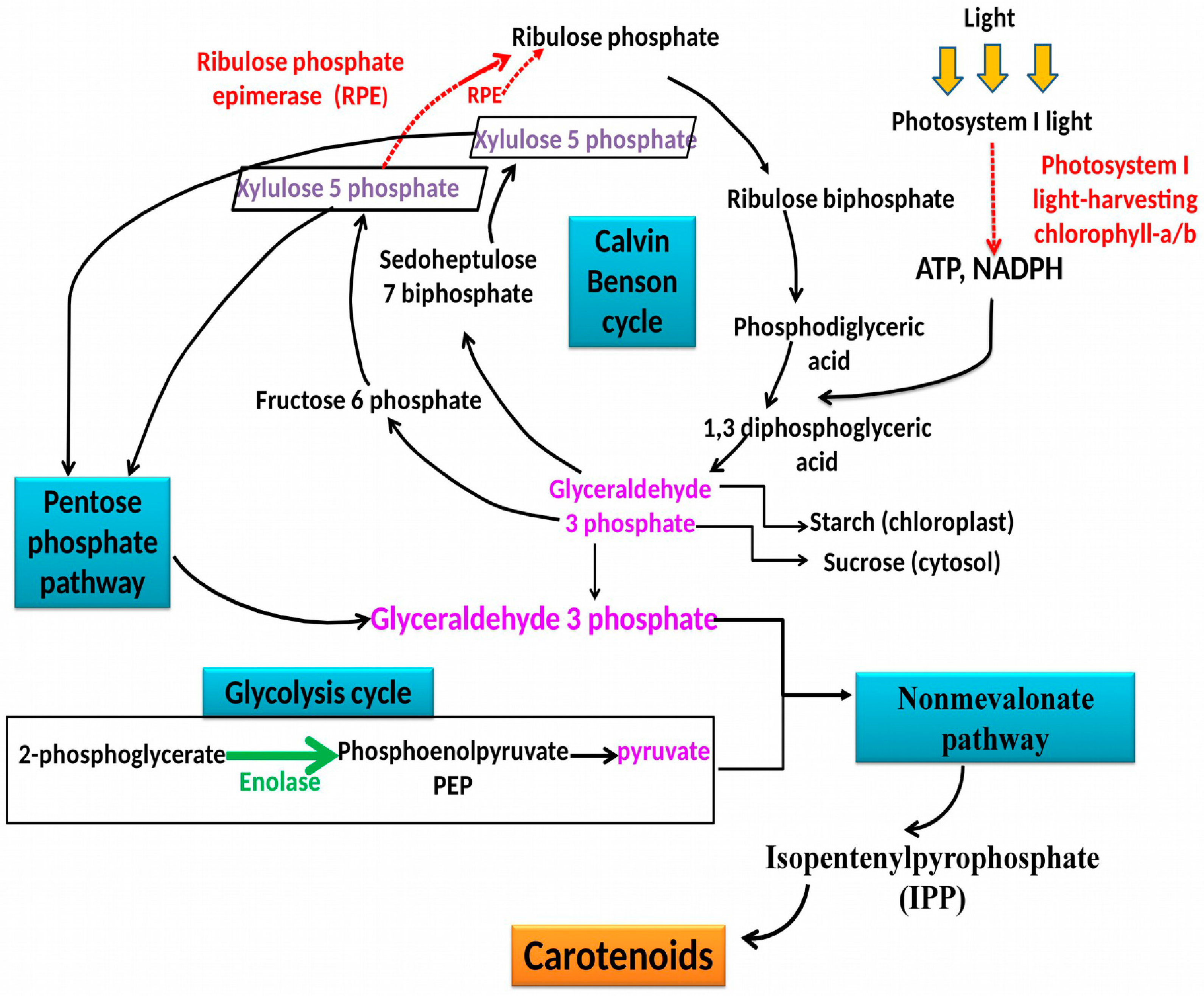
2.3.3. Functional Classification of Identified Proteins
3. Materials and Methods
3.1. Algal Strain, Medium and Culture Conditions
3.2. DNA Extraction, PCR Amplification, Sequencing, and Phylogenetic Analysis
3.3. Carotenoid Analysis
3.4. Experimental Design and Data Analysis
3.5. Statistical Analysis
3.6. Proteomic Analysis
3.6.1. Protein Extraction
3.6.2. Two-Dimensional Gel Electrophoresis (2-DE)
3.6.3. Image Acquisition and Data Analysis
3.6.4. In-Gel Tryptic Digestion and Protein Identification by Mass Spectrometry
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hosseini Tafreshi, A.; Shariati, M. Dunaliella biotechnology: Methods and applications. J. Appl. Microbiol. 2009, 107, 14–35. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.; Chan, K.W.; Ebrahimi, M. Antioxidant capacities of fucoxanthin-producing algae as influenced by their carotenoid and phenolic contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Ben Hsouna, A.; Hamdi, N.; Miladi, R.; Abdelkafi, S. Myrtus communis essential oil: Chemical composition and antimicrobial activities against food spoilage pathogens. Chem. Biodivers. 2014, 11, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, E.; Cifuentes, A. Benefits of using algae as natural sources of functional ingredients. J. Sci. Food Agric. 2013, 93, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Emtyazjoo, M.; Moghadasi, Z.; Rabbani, M.; Emtyazjoo, M.; Samadi, S.; Mossaffa, N. Anticancer effect of Dunaliella salina under stress and normal conditions against skin carcinoma cell line A431 in vitro. Iran. J. Fish. Sci. 2012, 11, 283–293. [Google Scholar]
- Yokthongwattana, K.; Chrost, B.; Behrman, S.; Casper-Lindley, C.; Melis, A. Photosystem II damage and repair cycle in the green alga Dunaliella salina: Involvement of a chloroplast-localized HSP70. Plant Cell Physiol. 2001, 42, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Lao, Y.-M.; Xiao, L.; Luo, L.-X.; Jiang, J.-G. Hypoosmotic expression of Dunaliella bardawil ζ-carotene desaturase is attributed to a hypoosmolarity-responsive element different from other key carotenogenic genes. Plant Physiol. 2014, 165, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amotz, A.; Avron, M. The Role of Glycerol in the osmotic regulation of the halophilic alga Dunaliella parva. Plant Physiol. 1973, 51, 875–878. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, M.R.; Tofighi, H.; Samadi, N.; Jamalifar, H. Effects of salinity on beta-carotene production by Dunaliella tertiolecta DCCBC26 isolated from the Urmia salt lake, north of Iran. Bioresour. Technol. 2006, 97, 2453–2456. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, M.A.; Wijffels, R.H. Effect of light intensity on β-carotene production and extraction by Dunaliella salina in two-phase bioreactors. Biomol. Eng. 2003, 20, 171–175. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Mojaat, M.; Pruvost, J.; Foucault, A.; Legrand, J. Effect of organic carbon sources and Fe2+ ions on growth and β-carotene accumulation by Dunaliella salina. Biochem. Eng. J. 2008, 39, 177–184. [Google Scholar] [CrossRef]
- Avron, M. The osmotic components of halotolerant algae. Trends Biochem. Sci. 1986, 11, 5–6. [Google Scholar] [CrossRef]
- Guarnieri, M.T.; Nag, A.; Smolinski, S.L.; Darzins, A.; Seibert, M.; Pienkos, P.T. Examination of triacylglycerol biosynthetic pathways via de novo transcriptomic and proteomic analyses in an unsequenced microalga. PLoS ONE 2011, 6, e25851. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.; Waridel, P.; Shevchenko, A.; Pick, U. Salt-induced changes in the plasma membrane proteome of the halotolerant alga Dunaliella salina as revealed by blue native gel electrophoresis and nano-LC-MS/MS analysis. Mol. Cell. Proteom. 2007, 6, 1459–1472. [Google Scholar] [CrossRef] [PubMed]
- Juin, C.; Bonnet, A.; Nicolau, E.; Bérard, J.B.; Devillers, R.; Thiéry, V.; Cadoret, J.P.; Picot, L. UPLC-MSE profiling of Phytoplankton metabolites: Application to the identification of pigments and structural analysis of metabolites in Porphyridium purpureum. Mar. Drugs 2015, 13, 2541–2558. [Google Scholar] [CrossRef] [PubMed]
- Brechlin, P.; Jahn, O.; Steinacker, P.; Cepek, L.; Kratzin, H.; Lehnert, S.; Jesse, S.; Mollenhauer, B.; Kretzschmar, H.A.; Wiltfang, J.; et al. Cerebrospinal fluid-optimized two-dimensional difference gel electrophoresis (2-D DIGE) facilitates the differential diagnosis of Creutzfeldt-Jakob disease. Proteomics 2008, 8, 4357–4366. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Hu, L.-S.; Chang, M.; Wu, J.; Winblad, B.; Zhu, J. Proteomic identification of potential protein markers in cerebrospinal fluid of GBS patients. Eur. J. Neurol. 2007, 14, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. A hundred years of Dunaliella research: 1905–2005. Saline Syst. 2005, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chtourou, H.; Dahmen, I.; Jebali, A.; Karray, F.; Hassairi, I.; Abdelkafi, S.; Ayadi, H.; Sayadi, S.; Dhouib, A. Characterization of Amphora sp., a newly isolated diatom wild strain, potentially usable for biodiesel production. Bioprocess Biosyst. Eng. 2015, 38, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, H.; Dahmen, I.; Hassairi, I.; Abdelkafi, S.; Sayadi, S.; Dhouib, A. Dunaliella sp. a wild algal strain isolated from the Sfax-Tunisia solar evaporating salt-ponds, a high potential for biofuel production purposes. J. Biobased Mater. Bioenergy 2014, 8, 27–34. [Google Scholar] [CrossRef]
- Ahmad, A.L.; Wong, S.S.; Teng, T.T.; Zuhairi, A. Optimization of coagulation-flocculation process for pulp and paper mill effluent by response surface methodological analysis. J. Hazard. Mater. 2007, 145, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Liska, A.J.; Shevchenko, A.; Pick, U.; Katz, A. Enhanced photosynthesis and redox energy production contribute to salinity tolerance in Dunaliella as revealed by homology-based proteomics. Plant Physiol. 2004, 136, 2806–2817. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.S.; Ferry, J.G. Prokaryotic carbonic anhydrases. FEMS Microbiol. Rev. 2000, 24, 335–366. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, K.; Miyachi, S. Carbonic anhydrase and CO2 concentrating mechanisms in microalgae and cyanobacteria. FEMS Microbiol. Lett. 1986, 39, 215–233. [Google Scholar] [CrossRef]
- Eriksson, M.; Villand, P.; Gardeström, P.; Samuelsson, G. Induction and regulation of expression of a low-CO2-induced mitochondrial carbonic anhydrase in Chlamydomonas reinhardtii. Plant Physiol. 1998, 116, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Dionisio-Sese, M.L.; Fukuzawa, H.; Miyachi, S. Light-induced carbonic anhydrase axpression in Chlamydomonas reinhardtii. Plant Physiol. 1990, 94, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Youn, S.H.; Ki, J.-S. Heat shock protein 70 and 90 genes in the harmful dinoflagellate Cochlodinium polykrikoides: Genomic structures and transcriptional responses to environmental stresses. Int. J. Genom. 2015, 2015, e484626. [Google Scholar]
- Li, Z.; Srivastava, P. Heat-shock proteins. Curr. Protoc. Immunol. 2004, Appendix 1. Appendix 1T. [Google Scholar] [CrossRef]
- Santoro, M.G. Heat shock factors and the control of the stress response. Biochem. Pharmacol. 2000, 59, 55–63. [Google Scholar] [CrossRef]
- Choi, Y.E.; Kwon, K.W.; Lee, J.C.; Woo, S.Y. Expression of the rice cytoplasmic cysteine synthase gene in tobacco reduces ozone-induced damage. Plant Biotechnol. Rep. 2007, 1, 93–100. [Google Scholar] [CrossRef]
- Hockin, N.L.; Mock, T.; Mulholland, F.; Kopriva, S.; Malin, G. The response of diatom central carbon metabolism to nitrogen starvation is different from that of green algae and higher plants. Plant Physiol. 2012, 158, 299–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, P.F.; Nielsen, E.E.; Koed, A.; Thomsen, D.S.; Olsvik, P.A.; Loeschcke, V. Interpopulation differences in expression of candidate genes for salinity tolerance in winter migrating anadromous brown trout (Salmo trutta L.). BMC Genet. 2008, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tine, M.; Bonhomme, F.; McKenzie, D.J.; Durand, J.-D. Differential expression of the heat shock protein Hsp70 in natural populations of the tilapia, Sarotherodon melanotheron, acclimatised to a range of environmental salinities. BMC Ecol. 2010, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Li, K.-L.; Chen, Y.; Lai, F.-X.; Fu, Q. Identification and function analysis of enolase gene NlEno1 from Nilaparvata lugens (Stål) (Hemiptera: Delphacidae). J. Insect Sci. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Pancholi, V. Multifunctional alpha-enolase: Its role in diseases. Cell. Mol. Life Sci. CMLS 2001, 58, 902–920. [Google Scholar] [CrossRef] [PubMed]
- Aaronson, R.M.; Graven, K.K.; Tucci, M.; McDonald, R.J.; Farber, H.W. Non-neuronal enolase is an endothelial hypoxic stress protein. J. Biol. Chem. 1995, 270, 27752–27757. [Google Scholar] [CrossRef] [PubMed]
- Ruan, K.; Duan, J.; Bai, F.; Lemaire, M.; Ma, X.; Bai, L. Function of Dunaliella salina (Dunaliellaceae) enolase and its expression during stress. Eur. J. Phycol. 2009, 44, 207–214. [Google Scholar] [CrossRef]
- Yang, Z.-K.; Ma, Y.-H.; Zheng, J.-W.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Proteomics to reveal metabolic network shifts towards lipid accumulation following nitrogen deprivation in the diatom Phaeodactylum tricornutum. J. Appl. Phycol. 2013, 26, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Nowitzki, U.; Wyrich, R.; Westhoff, P.; Henze, K.; Schnarrenberger, C.; Martin, W. Cloning of the amphibolic Calvin cycle/OPPP enzyme d-ribulose-5-phosphate 3-epimerase (EC 5.1.3.1) from spinach chloroplasts: Functional and evolutionary aspects. Plant Mol. Biol. 1995, 29, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Mastrobuoni, G.; Irgang, S.; Pietzke, M.; Aßmus, H.E.; Wenzel, M.; Schulze, W.X.; Kempa, S. Proteome dynamics and early salt stress response of the photosynthetic organism Chlamydomonas reinhardtii. BMC Genom. 2012, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.-L.; Chen, H.; Zhang, C.; Gao, L.-J.; Wang, X.-C.; Qiu, L.-L.; Wu, J.-F.; Jia, Y.-L.; Chen, H.; Zhang, C.; et al. Proteomic analysis of halotolerant proteins under high and low salt stress in Dunaliella salina using two-dimensional differential in-gel electrophoresis. Genet. Mol. Biol. 2016, 39, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.M.; Morrissey, P.J.; Guenther, J.E.; Nemson, J.A.; Harrison, M.A.; Allen, J.F.; Melis, A. Response of the photosynthetic apparatus in Dunaliella salina (Green Algae) to irradiance stress. Plant Physiol. 1990, 93, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.L.; Kavallaris, M.; McCarroll, J.A. Microtubules and their role in cellular stress in cancer. Front. Oncol. 2014, 4, 153. [Google Scholar] [CrossRef] [PubMed]
- Willem, S.; Srahna, M.; Devos, N.; Gerday, C.; Loppes, R.; Matagne, R.F. Protein adaptation to low temperatures: A comparative study of α-tubulin sequences in mesophilic and psychrophilic algae. Extremophiles 1999, 3, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Livanos, P.; Galatis, B.; Apostolakos, P. The interplay between ROS and tubulin cytoskeleton in plants. Plant Signal. Behav. 2014, 9, e28069. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Prášil, I.T.; Vítámvás, P. Protein contribution to plant salinity response and tolerance acquisition. Int. J. Mol. Sci. 2013, 14, 6757–6789. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Springer: New York, NY, USA, 1975; pp. 29–60. ISBN 978-1-4615-8716-3. [Google Scholar]
- Dammak, M.; Haase, S.M.; Miladi, R.; Ben Amor, F.; Barkallah, M.; Gosset, D.; Pichon, C.; Huchzermeyer, B.; Fendri, I.; Denis, M.; et al. Enhanced lipid and biomass production by a newly isolated and identified marine microalga. Lipids Health Dis. 2016, 15, 209. [Google Scholar] [CrossRef] [PubMed]
- Fendri, I.; Chaari, A.; Dhouib, A.; Jlassi, B.; Abousalham, A.; Carrière, F.; Sayadi, S.; Abdelkafi, S. Isolation, identification and characterization of a new lipolytic Pseudomonas sp., strain AHD-1, from Tunisian soil. Environ. Technol. 2010, 31, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R.; Lichtenthaler, H. Formulae and program to determine total carotenoids and chlorophylls A and B of leaf extracts in different solvents. In Advances in Agricultural Biotechnology; Sybesma, C., Ed.; Advances in Photosynthesis Research; Springer: Dordrecht, The Netherland, 1984; pp. 9–12. ISBN 978-90-247-2943-2. [Google Scholar]
- Box, G.E.P.; Behnken, D.W. Some new three level designs for the study of quantitative variables. Technometrics 1960, 2, 455–475. [Google Scholar] [CrossRef]
- Mathieu, J.E.; Heffner, T.S.; Goodwin, G.F.; Salas, E.; Cannon-Bowers, J.A. The influence of shared mental models on team process and performance. J. Appl. Psychol. 2000, 85, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.W.-F.; Lo, S.C.-L. The use of Trizol reagent (phenol/guanidine isothiocyanate) for producing high quality two-dimensional gel electrophoretograms (2-DE) of dinoflagellates. J. Microbiol. Methods 2008, 73, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Garnier, M.; Carrier, G.; Rogniaux, H.; Nicolau, E.; Bougaran, G.; Saint-Jean, B.; Cadoret, J.P. Comparative proteomics reveals proteins impacted by nitrogen deprivation in wild-type and high lipid-accumulating mutant strains of Tisochrysis lutea. J. Proteom. 2014, 105, 107–120. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [PubMed]
- Abdelkafi, S.; Abousalham, A.; Fendri, I.; Ogata, H.; Barouh, N.; Fouquet, B.; Scheirlinckx, F.; Villeneuve, P.; Carrière, F. Identification of a new phospholipase D in Carica papaya latex. Gene 2012, 499, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Ben Halima, N.; Khemakhem, B.; Fendri, I.; Ogata, H.; Baril, P.; Pichon, C.; Abdelkafi, S. Identification of new oat β-amylase by functional proteomics. Biochim. Biophys. Acta 2016, 1864, 52–61. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Factor | Coded Symbole | Level | ||
|---|---|---|---|---|
| −1 | 0 | +1 | ||
| Salinity | X1 | 58.44 | 87.66 | 116.88 |
| Light intensity (μmol photons·m−2·s−1) | X2 | 70 | 155 | 240 |
| Nitrogen·(g·L−1) | X3 | 0.02 | 0.06 | 0.1 |
| Coefficient | Value | Student’s t Test | Significance (%) |
|---|---|---|---|
| b0 | 6.421 | 10.77 | 0.0120 a |
| b1 | −0.513 | −1.41 | 21.9 |
| b2 | 1.395 | 3.82 | 1.24 c |
| b3 | −0.125 | −0.34 | 74.6 |
| b11 | −0.327 | −0.61 | 56.9 |
| b22 | −2.781 | −5.17 | 0.355 b |
| b33 | −2.305 | −4.29 | 0.781 b |
| b12 | −0.646 | −1.25 | 26.6 |
| b13 | 0.188 | 0.36 | 73.1 |
| b23 | 0.135 | 0.26 | 80.5 |
| Source of Variation | Sum of Squares | Degrees of Freedom | Mean Square | Ratio | Significance (%) | Significance |
|---|---|---|---|---|---|---|
| Regression | 64.4768 | 9 | 7.1641 | 6.7147 | 2.47 c | Significant |
| Residual | 5.3346 | 5 | 1.0669 | - | - | - |
| Lack of fit | 5.1483 | 3 | 1.7161 | 18.423 | 5.2 | Not significant |
| Error | 0.1863 | 2 | 0.0932 | - | - | - |
| Total | 69.811 | 14 | - | - | - | - |
| Spot Number | Hypothetical Function | ANOVA (p) | Fold | Standard Conditions | Stressful Conditions | Up/Down |
|---|---|---|---|---|---|---|
| 1144 | Enolase | 3.218 × 10−4 | 3.1 | 2.936 × 105 | 8.967 × 105 | up |
| 3056 | Carbonic anhydrase | 5.285 × 10−4 | 3.1 | 1.958 × 106 | 6.082 × 106 | up |
| 1246 | ATP synthase CF1 beta subunit (chloroplast) | 0.001 | 4.1 | 3.382 × 105 | 1.370 × 106 | up |
| 1209 | ATP synthase CF1 alpha subunit (chloroplast) | 0.002 | 3.4 | 1.002 × 105 | 3.448 × 105 | up |
| 1187 | beta-tubulin, partial | 0.003 | 3.1 | 2.854 × 105 | 8.967 × 105 | up |
| 2391 | Photosystem I light-harvesting chlorophyll-a/b protein | 0.005 | 3.5 | 4.321 × 106 | 1.227 × 106 | down |
| 1191 | ATP synthase CF1 beta subunit (chloroplast) | 0.011 | 5.0 | 4.455 × 105 | 2.232 × 106 | up |
| 2259 | Chloroplast ribulose phosphate -3-epimerase | 0.014 | 5.4 | 2.089 × 106 | 3.860 × 105 | down |
| 2218 | ATP synthase CF1 alpha subunit (chloroplast) | 0.021 | 2.7 | 3.416 × 106 | 1.273 × 106 | up |
| 2346 | Photosystem I light-harvesting chlorophyll-a/b protein 2 | 0.086 | 2.1 | 7.220 × 106 | 3.412 × 106 | down |
| 1255 | Heat shock protein HSP70 | 0.021 | 2.9 | 3.357 × 105 | 9.837 × 105 | up |
| Class | Spot Number | MW [kDa]/pI Theoretical | Score | SC [%] | Peptides | Hypothetical Function | Protein Accession Number |
|---|---|---|---|---|---|---|---|
| Carbon and energy metabolism | 3056 | 64.2/4.6 | 130.4 | 4.2 | 4 | Carbonic anhydrase | P54212 |
| 1209 | 54.5/5.2 | 62.4 | 4.8 | 2 | ATP synthase CF1 alpha subunit (chloroplast) | ACS95056 | |
| 2218 | 54.5/5.2 | 100.7 | 5.0 | 2 | ATP synthase CF1 alpha subunit (chloroplast) | ACS95056 | |
| 1187 | 51.8/4.8 | 3017.7 | 89.6 | 82 | ATP synthase CF1 beta subunit (chloroplast) | ACS95063 | |
| 1246 | 51.8/4.8 | 71.7 | 9.8 | 2 | ATP synthase CF1 beta subunit (chloroplast) | ACS95063 | |
| 1191 | 51.8/4.8 | 754.6 | 40.6 | 13 | ATP synthase CF1 beta subunit (chloroplast) | ACS95063 | |
| 2259 | 28.2/9 | 67.9 | 8.3 | 2 | Chloroplast ribulose phosphate-3-epimerase | AEF79975 | |
| Carbohydrate metabolism (Glycolysis/Gluconeogenesis) | 1144 | 52.1/5.3 | 2202.8 | 55.5 | 51 | Enolase | AIJ00881 |
| Protein chaperon (stress) | 1255 | 71.7/5.2 | 459.2 | 11.8 | 13 | Heat shock protein HSP70 | CAB71138 |
| 1246 | 71.7/5.2 | 217.6 | 8.2 | 5 | Heat shock protein HSP70 | CAB71138 | |
| Photosynthesis | 2391 | 24.7/6.4 | 200.0 | 15.4 | 5 | Photosystem I light-harvesting chlorophyll-a/b protein | ACN94453 |
| 2346 | 24.7/6.4 | 161.9 | 16.7 | 4 | harvesting chlorophyll-a/b protein 2 | ACN94453 | |
| Cellular Processes (Cytoskeleton) | 1187 | 39.1/4.7 | 952.7 | 46.1 | 22 | beta-tubulin, partial | AAY84712 |
| 1187 | 49.4/4.9 | 326.5 | 19.7 | 9 | alpha-tubulin protein | AEF79970 | |
| 1191 | 39.1/4.7 | 1683.3 | 57.3 | 47 | beta-tubulin, partial | AAY84712 | |
| 1191 | 49.4/4.9 | 489.9 | 26.6 | 13 | alpha-tubulin protein | AEF79970 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Amor, F.; Elleuch, F.; Ben Hlima, H.; Garnier, M.; Saint-Jean, B.; Barkallah, M.; Pichon, C.; Abdelkafi, S.; Fendri, I. Proteomic Analysis of the Chlorophyta Dunaliella New Strain AL-1 Revealed Global Changes of Metabolism during High Carotenoid Production. Mar. Drugs 2017, 15, 293. https://doi.org/10.3390/md15090293
Ben Amor F, Elleuch F, Ben Hlima H, Garnier M, Saint-Jean B, Barkallah M, Pichon C, Abdelkafi S, Fendri I. Proteomic Analysis of the Chlorophyta Dunaliella New Strain AL-1 Revealed Global Changes of Metabolism during High Carotenoid Production. Marine Drugs. 2017; 15(9):293. https://doi.org/10.3390/md15090293
Chicago/Turabian StyleBen Amor, Faten, Fatma Elleuch, Hajer Ben Hlima, Matthieu Garnier, Bruno Saint-Jean, Mohamed Barkallah, Chantal Pichon, Slim Abdelkafi, and Imen Fendri. 2017. "Proteomic Analysis of the Chlorophyta Dunaliella New Strain AL-1 Revealed Global Changes of Metabolism during High Carotenoid Production" Marine Drugs 15, no. 9: 293. https://doi.org/10.3390/md15090293
APA StyleBen Amor, F., Elleuch, F., Ben Hlima, H., Garnier, M., Saint-Jean, B., Barkallah, M., Pichon, C., Abdelkafi, S., & Fendri, I. (2017). Proteomic Analysis of the Chlorophyta Dunaliella New Strain AL-1 Revealed Global Changes of Metabolism during High Carotenoid Production. Marine Drugs, 15(9), 293. https://doi.org/10.3390/md15090293