Antibacterial Activities of Bacteria Isolated from the Marine Sponges Isodictya compressa and Higginsia bidentifera Collected from Algoa Bay, South Africa

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Terminal Restriction Fragment Length Polymorphism (T-RFLP) Analysis
2.3. Bacterial Culturing
2.4. Antimicrobial Screening
2.5. DNA Sequencing and Phylogenetic Analysis
2.6. Genome Sequencing
3. Results and Discussion
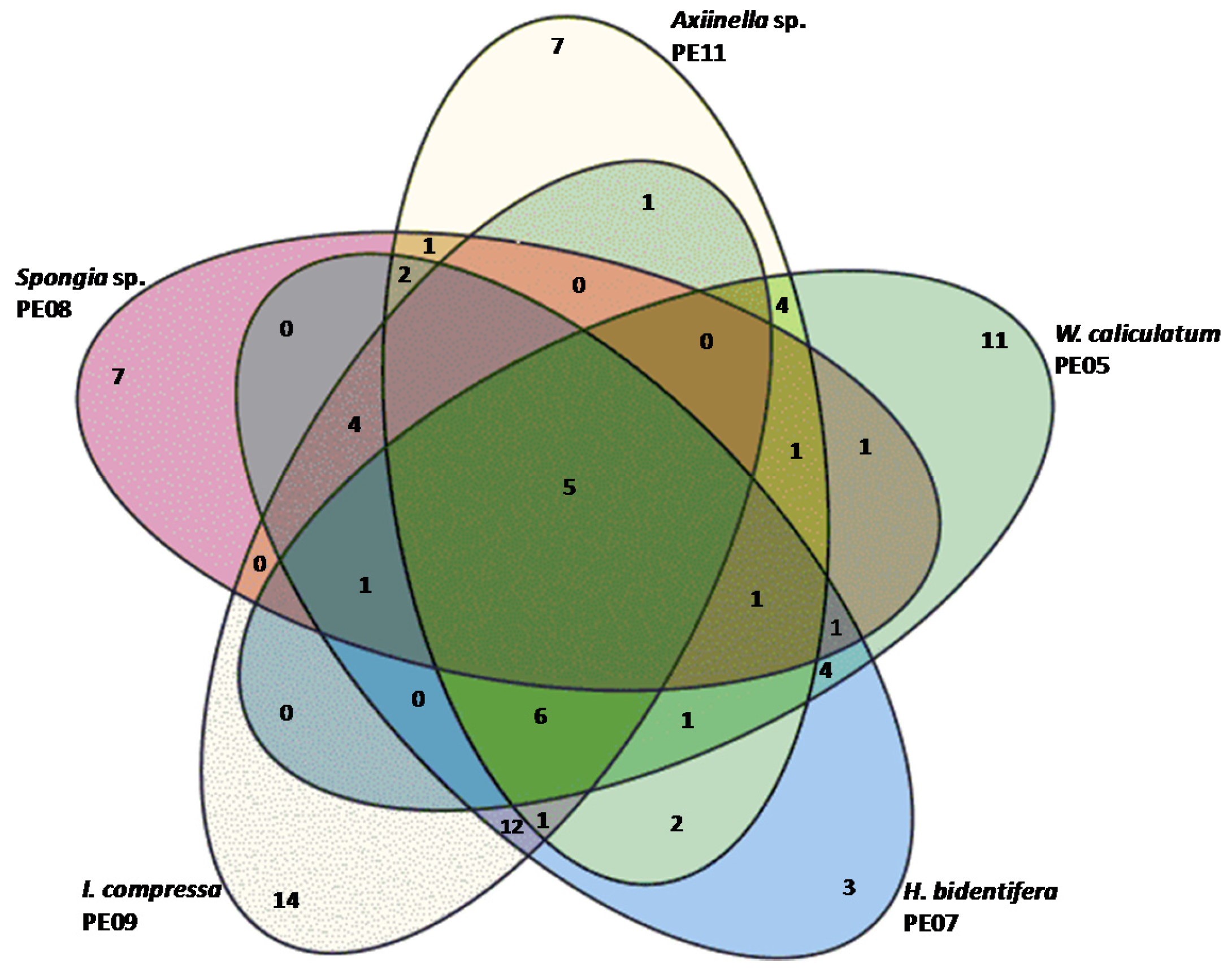
3.1. Bacterial Diversity Analysis and Culturing of Marine Sponge-Associated Bacteria
3.2. Antimicrobial Screening and Induction
3.3. 16S rRNA Gene Sequence Analysis of Bacteria Producing Antimicrobial Activity
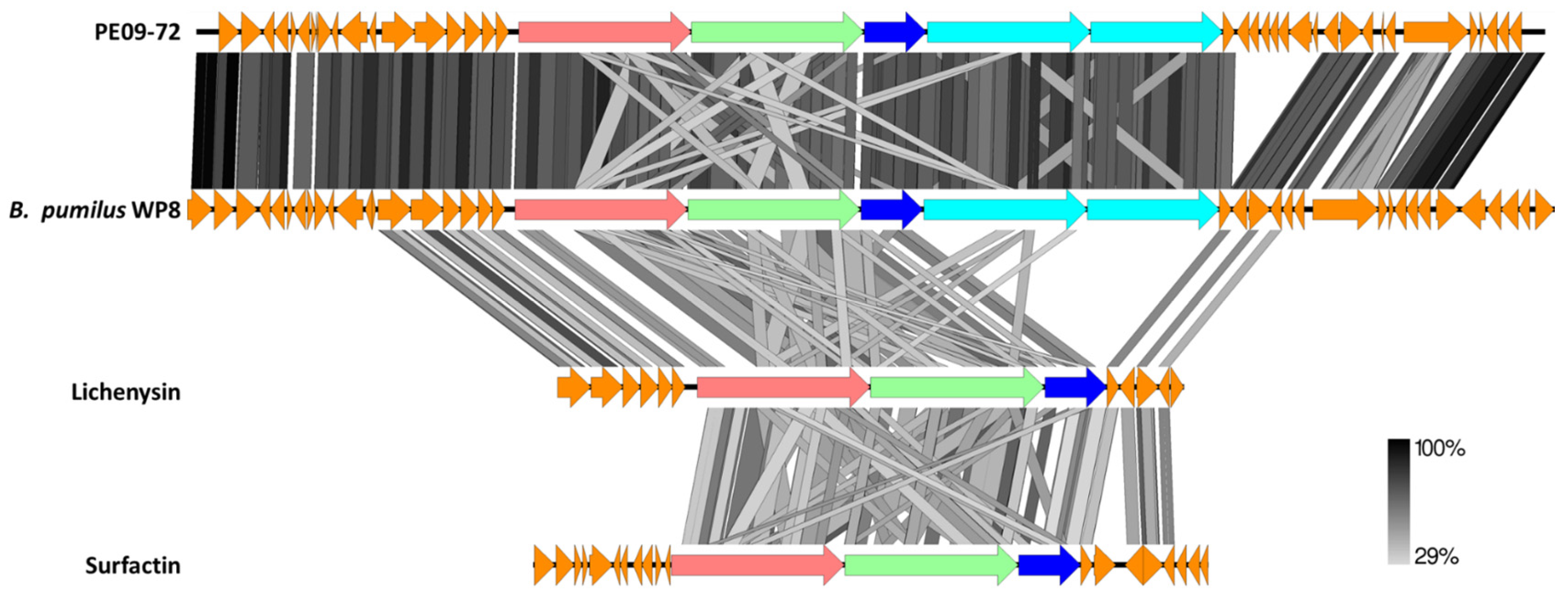
3.4. PE09-72 and PE09-221 Genome Sequence and Secondary Metabolism
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Piel, J. Bacterial symbionts: Prospects for the sustainable production of invertebrate-derived pharmaceuticals. Curr. Med. Chem. 2006, 13, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, C.C.P.; Costa, R.; Araújo, F.V.; Hajdu, E.; Peixoto, R.; Lins, U.; Rosado, A.S.; van Elsas, J.D. Diversity of bacteria in the marine sponge Aplysina fulva in Brazilian coastal waters. Appl. Environ. Microbiol. 2009, 75, 3331–3343. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, R.W.M.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; De Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.A. Global diversity of sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, S.; Weisz, J.B.; Lindquist, N.; Hentschel, U. Vertical transmission of a phylogenetically complex microbial consortium in the viviparous sponge Ircinia felix. Appl. Environ. Microbiol. 2007, 73, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Bayer, K.; Scheuermayer, M.; Fieseler, L.; Hentschel, U. Genomic mining for novel FADH2-dependent halogenases in marine sponge-associated microbial consortia. Mar. Biotechnol. 2013, 15, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhao, J.; Ke, C.; Wang, D. Antimicrobial activities of novel cultivable bacteria isolated from marine sponge Tedania anhelans. Chin. J. Oceanol. Limnol. 2013, 31, 581–590. [Google Scholar] [CrossRef]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, T.A.; Matcher, G.F.; Zhang, F.; Hill, R.T.; Davies-Coleman, M.T.; Dorrington, R.A. Diversity of bacterial communities associated with the Indian Ocean sponge Tsitsikamma favus that contains the bioactive pyrroloiminoquinones, tsitsikammamine A and B. Mar. Biotechnol. 2012, 14, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Ruan, J.; Huang, Y. Diversity and biosynthetic potential of culturable actinomycetes associated with marine sponges in the China seas. Int. J. Mol. Sci. 2012, 13, 5917–5932. [Google Scholar] [CrossRef] [PubMed]
- Fieseler, L.; Horn, M.; Wagner, M.; Hentschel, U. Discovery of the novel candidate phylum Poribacteria in marine sponges. Appl. Environ. Microbiol. 2004, 70, 3724–3732. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Taylor, M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Fenner, A.M. Drug Discovery from Marine Microbes. Microb. Ecol. 2013, 65, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Codling, C.E.; Jones, B.V.; Dobson, A.D.; Marchesi, J.R. Diversity of microbes associated with the marine sponge, Haliclona simulans, isolated from Irish waters and identification of polyketide synthase genes from the sponge metagenome. Environ. Microbiol. 2008, 10, 1888–1902. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Antifungal peptides in marine invertebrates. Invert. Surv. J. 2010, 7, 53–66. [Google Scholar]
- Phelan, R.W.; O’Halloran, J.A.; Kennedy, J.; Morrissey, J.P.; Dobson, A.D.; O’Gara, F.; Barbosa, T.M. Diversity and bioactive potential of endospore-forming bacteria cultured from the marine sponge Haliclona simulans. J. Appl. Microbiol. 2012, 112, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Rao, A.G.; Patil, M.R.; Saikia, B.; Bharadwaj, V.K.N.; Rao, J.G.S. Antimicrobial potential of metabolites extracted from bacterial symbionts associated with marine sponges in coastal area of Gulf of Mannar Biosphere, India. Lett. Appl. Microbiol. 2014, 58, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, I.; Kim, S.K. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, J.N.; Fan, L.; Brown, M.V.; Thomas, T.; Neilan, B.A. Deep sequencing of non-ribosomal peptide synthetases and polyketide synthases from the microbiomes of Australian marine sponges. ISME J. 2013, 7, 1842–1851. [Google Scholar] [CrossRef] [PubMed]
- Bergman, O.; Haber, M.; Mayzel, B.; Anderson, M.A.; Shpigel, M.; Hill, R.T.; Ilan, M. Marine-based cultivation of diacarnus sponges and the bacterial community composition of wild and maricultured sponges and their larvae. Mar. Biotechnol. 2011, 13, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Steinert, G.; Whitfield, S.; Taylor, M.W.; Thoms, C.; Schupp, P.J. Application of diffusion growth chambers for the cultivation of marine sponge-associated bacteria. Mar. Biotechnol. 2014, 16, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Oh, H.M.; Cho, J.C. Improved culturability of SAR11 strains in dilution-to-extinction culturing from the East Sea, West Pacific Ocean. FEMS Microbiol. Lett. 2009, 295, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, A.; Lewis, K.; Epstein, S.S. Incubation of environmental samples in a diffusion chamber increases the diversity of recovered isolates. Appl. Environ. Microbiol. 2007, 73, 6386–6390. [Google Scholar] [CrossRef] [PubMed]
- Joint, I.; Mühling, M.; Querellou, J. Culturing marine bacteria—An essential prerequisite for biodiscovery. Microb. Biotechnol. 2010, 3, 564–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branch, G.M.; Griffiths, C.L.; Branch, M.L.; Beckley, L.E. Two Oceans. A Guide to the Marine Life of Southern Africa; Struik Nature: Cape Town, South Africa, 2010. [Google Scholar]
- Griffiths, C.L.; Robinson, T.B.; Lange, L.; Mead, A. Marine biodiversity in South Africa: An evaluation of current states of knowledge. PLoS ONE 2010, 5, e12008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, E.M.; Beukes, D.R.; Kelly, M.; Samaai, T.; Barrows, L.R.; Marshall, K.M.; Sincich, C.; Davies-Coleman, M.T. Cytotoxic pyrroloiminoquinones from four new species of South African Latrunculid sponges. J. Nat. Prod. 2004, 67, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Samaai, T.; Gibbons, M.J.; Kerwath, S.; Yemane, D.; Sink, K. Sponge richness along a bathymetric gradient within the Isimangaliso Wetland Park, South Africa. Mar. Biodivers. 2010, 40, 205–217. [Google Scholar] [CrossRef]
- Higgin, T. Description of some Sponges obtained during a Cruise of the Steam-Yacht ‘Argo’ in the Caribbean and neighbouring Seas. J. Nat. Hist. 1877, 19, 291–299. [Google Scholar] [CrossRef]
- Ridley, S.O.; Dendy, A. Preliminary Report on the Monaxonida collected by H.M.S. ‘Challenger’. Ann. Mag. Nat. Hist. 1886, 18, 325–351, 470–493. [Google Scholar] [CrossRef]
- Esper, E.C.J. Die Pflanzenthiere in Abbildungen Nach der Natur Mit Farben Erleuchtet, Nebst Beschreibungen, Zweyter Theil; Raspe: Nürnberg, Germany, 1794; pp. 1–303. [Google Scholar]
- Topsent, E. Éponges de Lamarck conservées au Muséum de Paris. Archives du Muséum National D’histoire Naturelle 1931, 5, 1–56. [Google Scholar]
- Kirkpatrick, R. Descriptions of South African Sponges. Part III. Mar. Investig. S. Afr. 1903, 2, 233–264. [Google Scholar]
- Vimala, A.; Xavier Innocent, B.; Huxley, V.A.J. Phylogenetic investigations on the endosymbiotic bacteria of Axinella donnani. J. Microb. Biochem. Technol. 2015, 7, 318–322. [Google Scholar]
- Haber, M.; Ilan, M. Diversity and antibacterial activity of bacteria cultured from Mediterranean Axinella spp. sponges. J. Appl. Microbiol. 2014, 116, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Bauvais, C.; Zirah, S.; Piette, L.; Chaspoul, F.; Domart-Coulon, I.; Chapon, V.; Gallice, P.; Rebuffat, S.; Perez, T.; Bourguet-Kondracki, M.-L. Sponging up metals: Bacteria associated with the marine sponge Spongia officinalis. Mar. Environ. Res. 2015, 104, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Noyer, C.; Casamayor, E.O.; Becerro, M.A. Environmental heterogeneity and microbial inheritance influence sponge-associated bacterial composition of Spongia lamella. Microb. Ecol. 2014, 68, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liles, M.R.; Williamson, L.L.; Rodbumrer, J.; Torsvik, V.; Goodman, R.M.; Handelsman, J. Recovery, purification, and cloning of high-molecular-weight DNA from soil microorganisms. Appl. Environ. Microbiol. 2008, 74, 3302–3305. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.C.; Tolker-Nelson, T.; Givskov, M.; Molin, S. Biased 16S rDNA PCR amplification caused by interference from DNA flanking template region. FEMS Microbiol. Ecol. 1998, 15, 25–36. [Google Scholar] [CrossRef]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Meth. 2003, 55, 541–555. [Google Scholar] [CrossRef]
- Culman, S.W.; Bukowski, R.; Gauch, H.G.; Cadillo-Quiroz, H.; Buckley, D.H. T-REX: Software for the processing and analysis of T-RFLP data. BMC Bioinform. 2009, 10, 171. Available online: http://trex.biohpc.org/ (accessed on 3 February 2017). [Google Scholar] [CrossRef] [PubMed]
- Blackwood, C.B.; Hudleston, D.; Zak, D.R.; Buyer, J.S. Interpreting ecological diversity indices applied to terminal restriction fragment length polymorphism data: Insights from simulated microbial communities. Appl. Environ. Microbiol. 2007, 73, 5276–5283. [Google Scholar] [CrossRef] [PubMed]
- Nocker, A.; Burr, M.; Camper, A.K. Genotypic microbial community profiling: A critical technical review. Microb. Ecol. 2007, 54, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Schütte, U.M.E.; Abdo, Z.; Bent, S.J.; Shyu, C.; Williams, C.J.; Pierson, J.D.; Forney, L.J. Advances in the use of terminal restriction fragment length polymorphism (T-RFLP) analysis of 16S rRNA genes to characterize microbial communities. Appl. Microbiol. Biotechnol. 2008, 80, 365–380. [Google Scholar]
- Ramond, J.-B.; Tshabuse, F.; Bopda, C.W.; Cowan, D.A.; Tuffin, M.I. Evidence of variability in the structure and recruitment of rhizospheric and endophytic bacterial communities associated with arable sweet sorghum (Sorghum bicolor (L) Moench). Plant Soil 2013, 372, 265–278. [Google Scholar] [CrossRef]
- Lavy, A.; Keren, R.; Haber, M.; Schwartz, I.; Ilan, M. Implementing sponge physiological and genomic information to enhance the diversity of its culturable associated bacteria. FEMS Microbiol. Ecol. 2014, 87, 486–502. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; CRC Press: Florence, KY, USA, 2010; pp. 1–2040. [Google Scholar]
- Yang, Y.; Zhang, J.; Wu, H.; Gong, M. Genetic diversity analysis of Azotobacter in cotton soil of Yangtze River Basin. J. Chem. Pharm. Res. 2013, 5, 54–58. [Google Scholar]
- Sipkema, D.; Schippers, K.; Maalcke, W.J.; Yang, Y.; Salim, S.; Blanch, H.W. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (Gellius) sp. Appl. Environ. Microbiol. 2011, 77, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Intra, B.; Mungsuntisuk, I.; Nihira, T.; Igarashi, Y.; Panbangred, W. Identification of actinomycetes from plant rhizospheric soils with inhibitory activity against Colletotrichum spp., the causative agent of anthracnose disease. BMC Res. Notes 2011, 4, 98. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.F.; Platas, G.; Fillola, A.; Jiménez, M.R.; Collado, J.; Vicente, F.; Martín, J.; González, A.; Bur-Zimmermann, J.; Tormo, J.R.; et al. Enhancement of antibiotic and secondary metabolite detection from filamentous fungi by growth on nutritional arrays. J. Appl. Microbiol. 2008, 104, 1644–1658. [Google Scholar] [CrossRef] [PubMed]
- Nakata, K.; Yoshimoto, A.; Yamada, Y. Promotion of antibiotic production by high ethanol, high NaCl concentration, or heat shock in Pseudomonas fluorescens S272. Biosci. Biotechnol. Biochem. 1999, 63, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. Available online: http://www.megasoftware.net/ (accessed on 8 February 2017). [Google Scholar] [CrossRef] [PubMed]
- Illumina Nextera® XT DNA Sample Preparation Guide, Part # 15031942 Rev. C, 2012. Available online: https://tinyurl.com/jfd867j (accessed on 8 February 2017).
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, 7–19. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 8 February 2017). [Google Scholar]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N.; Perez, T.; Rodrigo, A.; Schupp, P.J.; Vacelet, J.; et al. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012, 6, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Tsai, P.; Simister, R.L.; Deines, P.; Botte, E.; Ericson, G.; Schmitt, S.; Webster, N.S. “Sponge-specific” bacteria are widespread (but rare) in diverse marine environments. ISME J. 2013, 7, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Simister, R.L.; Taylor, M.W.; Tsai, P.; Fan, L.; Bruxner, T.J.; Crowe, M.L.; Webster, N. Thermal stress responses in the bacterial biosphere of the great barrier reef sponge, Rhopaloeides odorabile. Environ. Microbiol. 2012, 14, 3232–3246. [Google Scholar] [CrossRef] [PubMed]
- Simister, R.L.; Deines, P.; Botté, E.S.; Webster, N.S.; Taylor, M.W. Sponge-specific clusters revisited: A comprehensive phylogeny of sponge-associated microorganisms. Environ. Microbiol. 2012, 14, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Erwin, P.M.; Olson, J.B.; Thacker, R.W. Phylogenetic diversity, host-specificity and community profiling of sponge-associated bacteria in the northern Gulf of Mexico. PLoS ONE 2011, 6, e26806. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Demain, A.L. Metabolic regulation of fermentation processes. Enzyme Microb. Technol. 2002, 31, 895–906. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Harrison, W.T.; Deng, H.; Okoro, C.K.; Asenjo, J.A.; Andrews, B.A.; Bull, A.T.; Goodfellow, M.; Ebel, R.; et al. Diverse metabolic profiles of a Streptomyces strain isolated from a hyper-arid environment. J. Nat. Prod. 2011, 74, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Dai, S.; Jiang, S.; Wang, G.; Liu, G.; Wu, H.; Li, X. Culture-dependent and culture-independent diversity of Actinobacteria associated with the marine sponge Hymeniacidon perleve from the South China Sea. Antonie van Leeuwenhoek 2010, 98, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Wu, H.; Xie, L.; Wang, G.; Dai, S.; Chen, M.; Yang, K.; Li, X. A method to type the potential angucycline producers in actinomycetes isolated from marine sponges. Antonie van Leeuwenhoek 2011, 99, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Vynne, N.G.; Mansson, M.; Gram, L. Gene sequence based clustering assists in dereplication of Pseudoalteromonas luteoviolacea strains with identical inhibitory activity and antibiotic production. Mar. Drugs 2012, 10, 1729–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, K.R.; Haltli, B.; Gill, K.A.; Correa, H.; Berrué, F.; Kerr, R.G. Exploring the diversity and metabolic potential of actinomycetes from temperate marine sediments from Newfoundland, Canada. J. Ind. Microbiol. Biotechnol. 2015, 42, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lai, Q.; Du, J.; Shao, Z. Bacillus zhangzhouensis sp. nov. and Bacillus australimaris sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.L.; O’Sullivan, L.; Tan, S.P.; McLoughlin, P.; Hughes, H.; O’Connor, P.M.; Cotter, P.D.; Lawlor, P.G.; Gardiner, G.E. Assessment of the bacteriocinogenic potential of marine bacteria reveals lichenicidin production by seaweed-derived Bacillus spp. Mar. Drugs 2012, 10, 2280–2299. [Google Scholar] [CrossRef]
- Sumi, C.D.; Yang, B.W.; Yeo, I.-C.; Hahm, Y.T. Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Can. J. Microbiol. 2014, 61, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Berrue, F.; Ibrahim, A.; Boland, P.; Kerr, R.G. Newly isolated marine Bacillus pumilus (SP21): A source of novel lipoamides and other antimicrobial agents. Pure Appl. Chem. 2009, 81, 1027–1031. [Google Scholar] [CrossRef]
- Ouoba, L.I.I.; Diawara, B.; Jespersen, L.; Jakobsen, M. Antimicrobial activity of Bacillus subtilis and Bacillus pumilus during the fermentation of African locust bean (Parkia biglobosa) for Soumbala production. J. Appl. Microbiol. 2007, 102, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.J.; Langley, D.; Bushell, M.E. Oxygen limitation can induce microbial secondary metabolite formation: Investigations with miniature electrodes in shaker and bioreactor culture. Microbiology 1995, 141, 663–669. [Google Scholar] [CrossRef]
- Jami, M.; Ghanbari, M.; Kneifel, W.; Domig, K.J. Phylogenetic diversity and biological activity of culturable Actinobacteria isolated from freshwater fish gut microbiota. Microbiol. Res. 2015, 175, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kim, I.G.; Kang, K.H.; Oh, T.K.; Park, Y.H. Bacillus marisflavi sp. nov. and Bacillus aquimaris sp. nov., isolated from sea water of a tidal flat of the Yellow Sea in Korea. Int. J. Syst. Evol. Microbiol. 2003, 53, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Yeo, S.H.; Oh, T.K.; Park, Y.H. Psychrobacter alimentarius sp. nov., isolated from squid jeotgal, a traditional Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2005, 55, 171. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Xiong, M.; Tang, T.; Song, L.; Hu, X.; Zhao, M.; Zhang, K. Aeromicrobium camelliae sp. nov., isolated from Pu’er tea. Int. J. Syst. Evol. Microbiol. 2015, 65, 4369–4373. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.-F.; Lin, P.; Chu, X.; Kook, M.C.; Li, C.-T.; Yi, T.-H. Aeromicrobium halotolerans sp. nov., isolated from desert soil sample. Arch. Microbiol. 2016, 198, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Magarvey, N.A.; Ehling-Schulz, M.; Walsh, C.T. Characterization of the cereulide NRPS α-hydroxy acid specifying modules: Activation of α-keto Acids and chiral reduction on the assembly line. J. Am. Chem. Soc. 2006, 128, 10698–10699. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Li, L.; Brock, N.L.; Deng, Z.; Lin, S. Functional characterization of PyrG, an unusual nonribosomal peptide synthetase module from the pyridomycin biosynthetic pathway. Chem. Biol. Chem. 2016. [Google Scholar] [CrossRef] [PubMed]
- Heravi, K.M.; Shali, A.; Naghibzadeh, N.; Ahmadian, G. Characterization of cis-acting elements residing in the chitinase promoter of Bacillus pumilus SG2. World J. Microbiol. Biotechnol. 2014, 30, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Ozcengiz, G.; Alaeddinoglu, N.G.; Demain, A.L. Regulation of biosynthesis of bacilysin by Bacillus subtilis. J. Ind. Microbiol. 1990, 6, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Molohon, K.J.; Melby, J.O.; Lee, J.; Evans, B.S.; Dunbar, K.L.; Bumpus, S.B.; Kelleher, N.L.; Mitchell, D.A. Structure determination and interception of biosynthetic intermediates for the plantazolicin class of highly discriminating antibiotics. ACS Chem. Biol. 2011, 6, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Lee, S.D. Aeromicrobium ponti sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, D.; Kokcha, S.; Lagier, J.-C.; Nguyen, T.-T.; Raoult, D.; Fournier, P.-E. Genome sequence and description of Aeromicrobium massiliense sp. nov. Stand. Genom. Sci. 2012, 7, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Lightfield, J.; Fram, N.R.; Ely, B. Across bacterial phyla, distantly-related genomes with similar genomic GC content have similar patterns of amino acid usage. PLoS ONE 2011, 6, e17677. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.; Lin, L. Gene content dissimilarity for subclassification of highly similar microbial strains. BMC Genom. 2016, 17, 647. [Google Scholar] [CrossRef] [PubMed]





| Sample ID | Marine Sponge ID | Species Specific OTUs | S | 1 − λ |
|---|---|---|---|---|
| PE05 | W. caliculatum | 11 | 36 | 0.6923 |
| PE07 | H. bidentifera | 3 | 43 | 0.9388 |
| PE08 | Spongia (spongia) sp. 001RSASPN | 7 | 24 | 0.8514 |
| PE09 | I. compressa | 14 | 44 | 0.9306 |
| PE11 | Axiinella sp. 007RSASPN | 7 | 32 | 0.7706 |
| Isolate | Test Strains | ||||
|---|---|---|---|---|---|
| Gram Negative | Gram Positive | ||||
| E. coli 1699 | P. putida | M. smegmatis | B. cereus | S. epidermidis | |
| PE07-5 | GYM | GYM | ZBA | ||
| PE07-7 | TSA, ZBA | TSA, ZBA | |||
| PE07-86 | TSA | ||||
| PE07-105 | GYM | ||||
| PE07-133 | GYM | ||||
| PE07-143 | TSA, ZBA | ||||
| PE07-144 | GYM | GYM | |||
| PE07-172 | TSA | ||||
| PE07-200 | GYM | ||||
| PE07-201 | GYM | ||||
| PE07-204 | GYM | ||||
| PE07-207 | GYM | GYM | |||
| PE09-7 | TSA | ||||
| PE09-73 | ACM | ||||
| PE09-87 | GYM | GYM | GYM | ||
| PE09-100 | ZBA | ||||
| PE09-105 | GYM, TSA | TSA, GYM | TSA | ||
| PE09-108 | GYM | ||||
| PE09-110 | ZBA | ||||
| PE09-119 | ZBA, GYM | TSA, ZBA | ACM, TSA | ||
| PE09-124 | ACM | ||||
| PE09-140 | ACM | ||||
| PE09-142 | ACM | ||||
| PE09-168 | ACM | ||||
| PE09-197 | ACM | ||||
| PE09-210 | ACM | ||||
| PE09-213 | ACM | ||||
| PE09-221 | ZBA | ||||
| PE09-228 | ACM | ||||
| PE09-229 | ACM | ||||
| PE09-235 | ACM | ||||
| PE09-266 | ZBA | ||||
| Total hits | 7 | 5 | 8 | 17 | 6 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum/Class | Closest Relative (as Present in EzTaxon) | Identity (%) | Isolate | Isolation Medium |
|---|---|---|---|---|
| Actinobacteria | Aeromicrobium erythreum NRRL B-3381 | 96.5 | PE09-221 | GYM |
| Arthrobacter citreus DSM 20133 | 99.9 | PE09-119 | ACM | |
| Brevibacterium luteolum CF87 | 100 | PE09-168 | GYM | |
| Citricoccus nitrophenolicus PNP1 | 99.7 | PE09-100 | SCN | |
| Citricoccus nitrophenolicus PNP1 | 99.8 | PE09-110 | 172 | |
| Curtobacterium oceanosedimentum ATCC 31317 | 99.3 | PE09-7 | GYM | |
| Micrococcus yunnanensis YIM 65004 | 99.7 | PE07-133 | R2A | |
| Firmicutes | Bacillus kochii WCC 4582 | 99.5 | PE07-103 | MAA |
| Bacillus marisflavi TF-11 | 100 | PE07-7 | ZBA | |
| Bacillus vietnamesis B-23890 | 99.7 | PE07-5 | ZBA | |
| Bacillus zhangzhouensis DW5-4 | 99.9 | PE09-72 | ZBA | |
| Bacillus vietnamesis B-23890 | 99.7 | PE07-144 | ZBA | |
| Oceanobacillus picturae LMG 19492 | 99.5 | PE07-105 | MAA | |
| Oceanobacillus picturae LMG 19492 | 100 | PE09-108 | ZBA | |
| Sporosarcina aquimarina SW28 | 98.8 | PE07-172 | 172 | |
| Staphylococcus cohnii ATCC 29974 | 100 | PE07-204 | ZBA | |
| Staphylococcus epidermidis ATCA 1490 | 99.9 | PE07-201 | TSA | |
| Staphylococcus warneri ATCC 27836 | 99.9 | PE07-200 | TSA | |
| Gammaproteobacteria | Halomonas titanicae | 99.9 | PE09-210 | CCA |
| Halomonas titanicae | 99.9 | PE09-228 | SEM | |
| Halomonas titanicae | 99.9 | PE09-229 | SEM | |
| Kushneria pakistanensis NCCP-934 | 98.9 | PE09-73 | PMM | |
| Kushneria pakistanensis NCCP-934 | 98.9 | PE09-124 | ACM | |
| Kushneria pakistanensis NCCP-934 | 98.9 | PE09-266 | ANFA | |
| Pseudomonas fulva NBRC 16637 | 100 | PE09-140 | SNA | |
| Pseudomonas fulva NBRC 16637 | 100 | PE09-197 | ANFA | |
| Psychrobacter alimentarius JG-100 | 99.9 | PE09-87 | ACM | |
| Alphaproteobacteria | Pseudovibrio ascidiaceicola DSM 16392 | 100 | PE09-55 | TSA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matobole, R.M.; Van Zyl, L.J.; Parker‐Nance, S.; Davies‐Coleman, M.T.; Trindade, M. Antibacterial Activities of Bacteria Isolated from the Marine Sponges Isodictya compressa and Higginsia bidentifera Collected from Algoa Bay, South Africa. Mar. Drugs 2017, 15, 47. https://doi.org/10.3390/md15020047
Matobole RM, Van Zyl LJ, Parker‐Nance S, Davies‐Coleman MT, Trindade M. Antibacterial Activities of Bacteria Isolated from the Marine Sponges Isodictya compressa and Higginsia bidentifera Collected from Algoa Bay, South Africa. Marine Drugs. 2017; 15(2):47. https://doi.org/10.3390/md15020047
Chicago/Turabian StyleMatobole, Relebohile Matthew, Leonardo Joaquim Van Zyl, Shirley Parker‐Nance, Michael T. Davies‐Coleman, and Marla Trindade. 2017. "Antibacterial Activities of Bacteria Isolated from the Marine Sponges Isodictya compressa and Higginsia bidentifera Collected from Algoa Bay, South Africa" Marine Drugs 15, no. 2: 47. https://doi.org/10.3390/md15020047
APA StyleMatobole, R. M., Van Zyl, L. J., Parker‐Nance, S., Davies‐Coleman, M. T., & Trindade, M. (2017). Antibacterial Activities of Bacteria Isolated from the Marine Sponges Isodictya compressa and Higginsia bidentifera Collected from Algoa Bay, South Africa. Marine Drugs, 15(2), 47. https://doi.org/10.3390/md15020047