Anti-Obesity and Anti-Diabetic Effect of Neoagarooligosaccharides on High-Fat Diet-Induced Obesity in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of Streptomyces coelicolor β-Agarase DagA
2.2. Preparation of Neoagarooligosaccharides (NAOs)
2.3. Effects of NAOs Intake in High-Fat Diet (HFD)-Induced Obese Mice
2.4. Histology of Mice Adipose and Liver Tissues
2.5. Biochemical Parameter Analysis
2.6. Oral Glucose Tolerance Tests (OGTT)
2.7. Statistical Analysis
3. Results
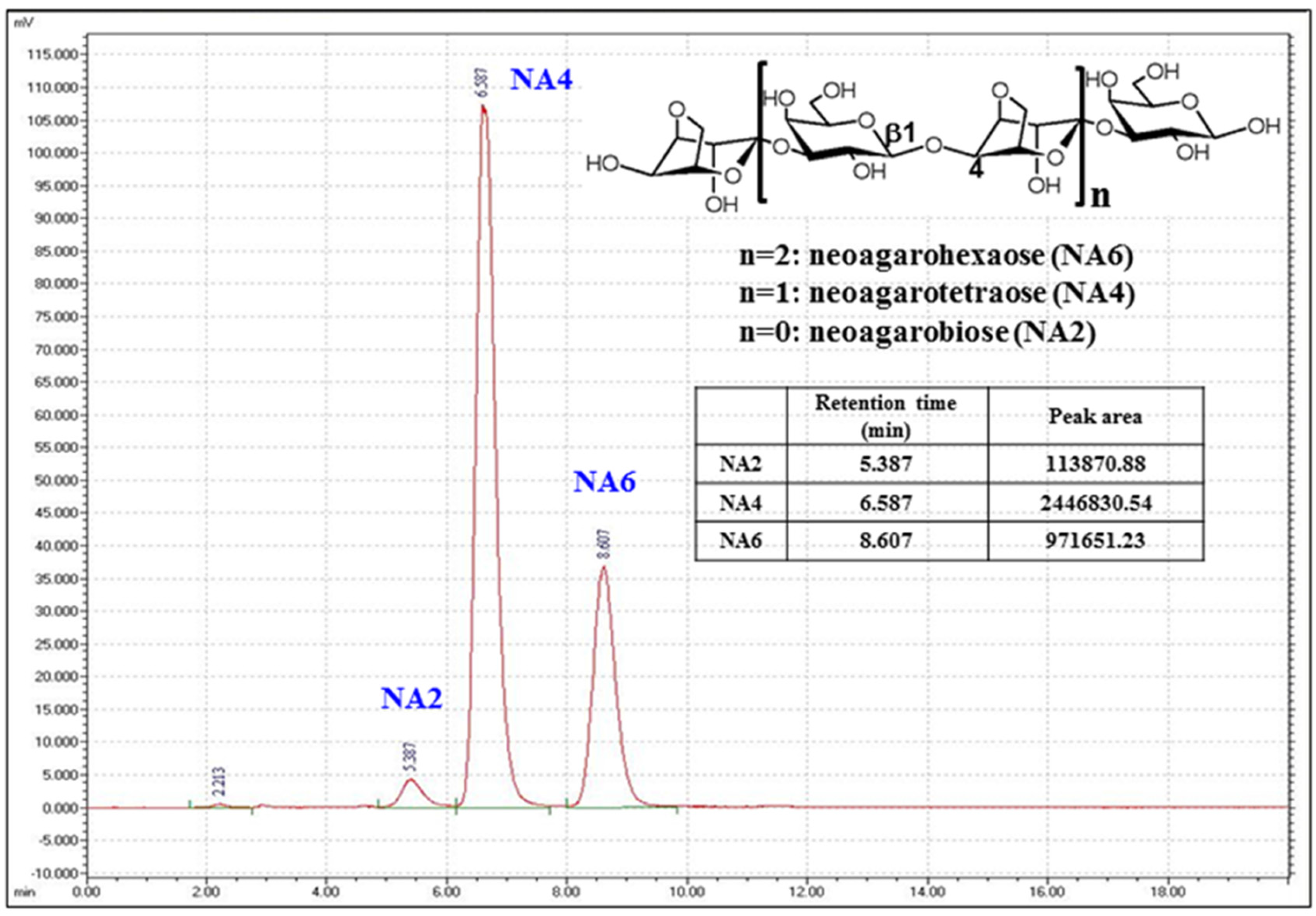
3.1. Analysis of the Composition of NAOs Produced by β-Agarase DagA
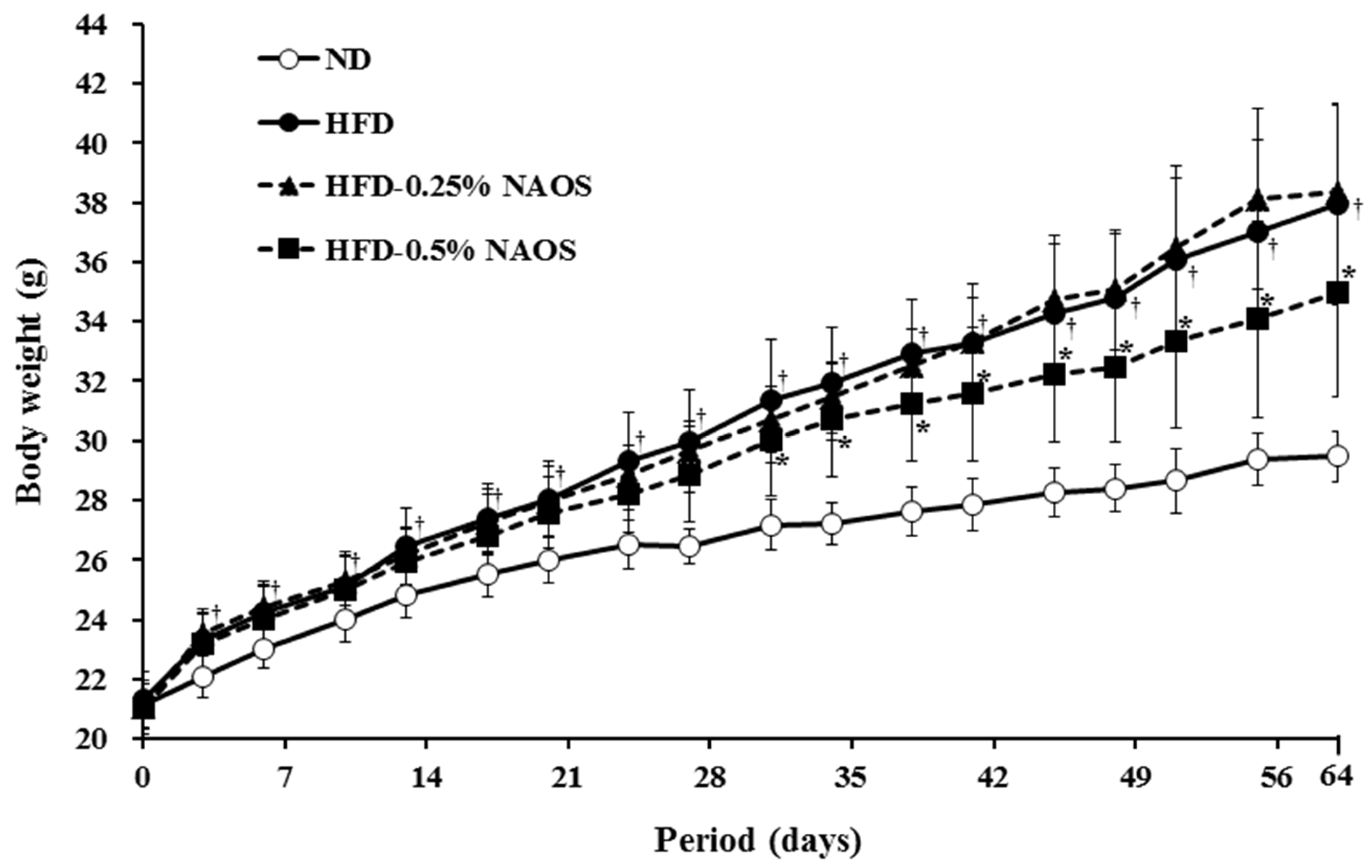
3.2. Effects of NAOs on Body Weight and Food Intake
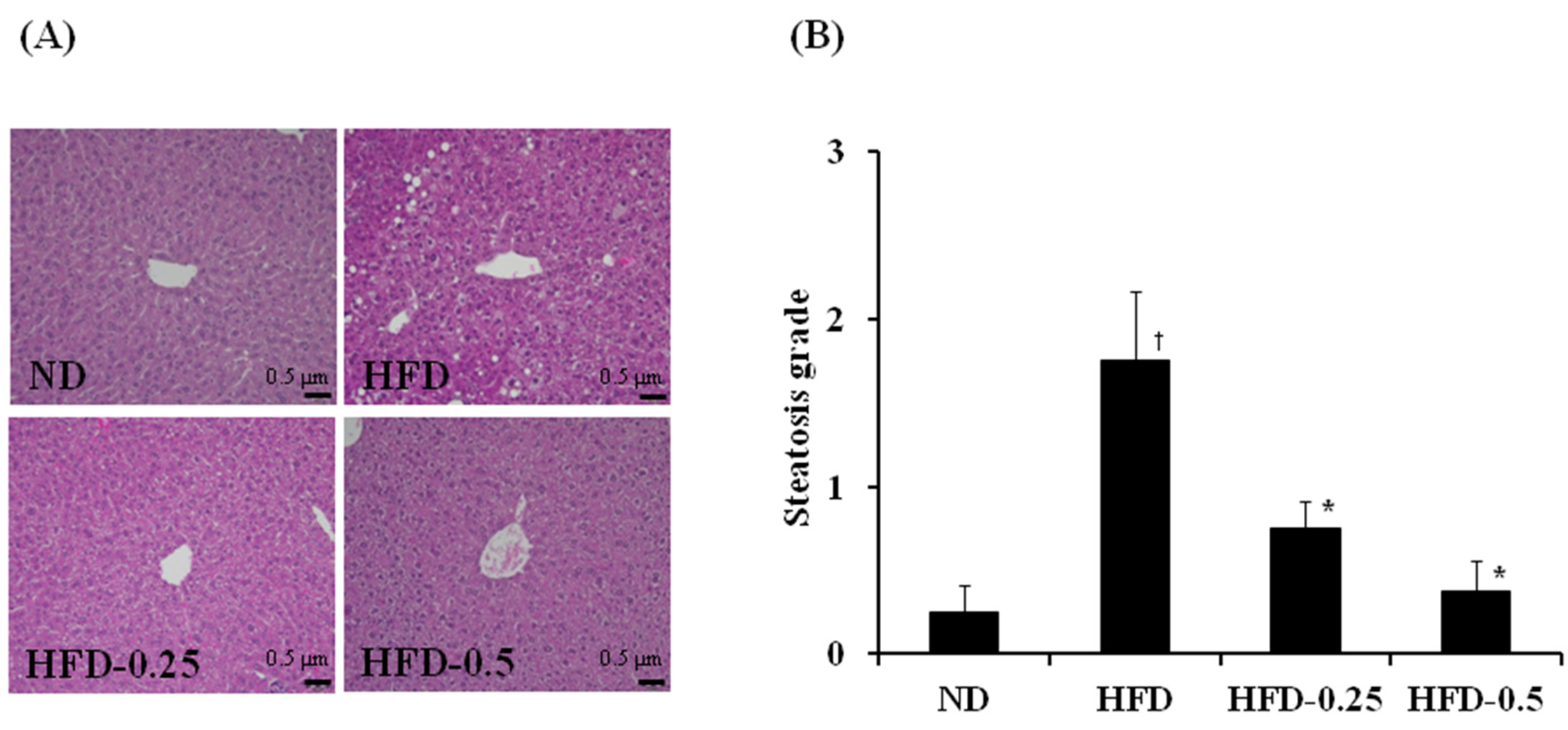
3.3. Effects of NAOs on HFD-Induced Fatty Liver
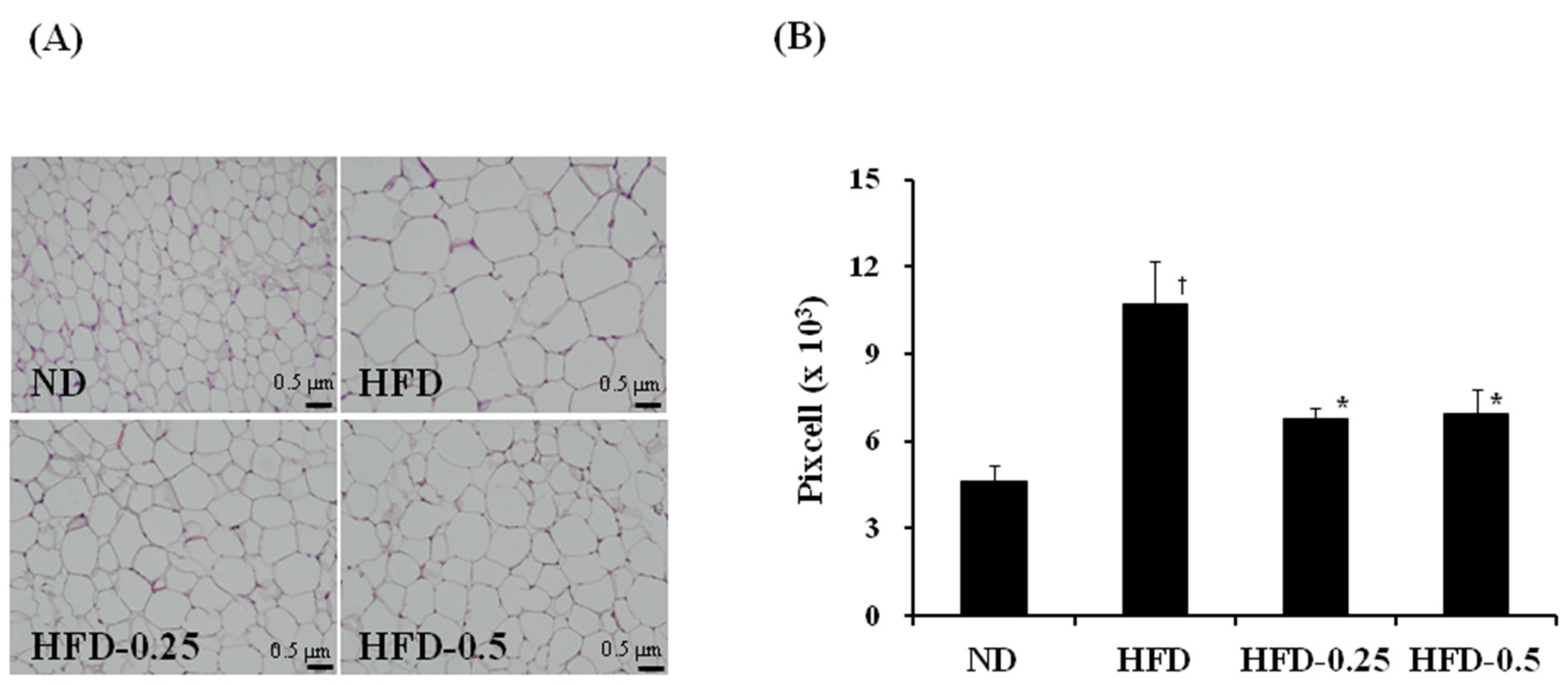
3.4. Effects of NAOs on Development of the Perirenal and Epididymal Adipose Tissues
3.5. Effects of NAOs on Serum Lipid Levels
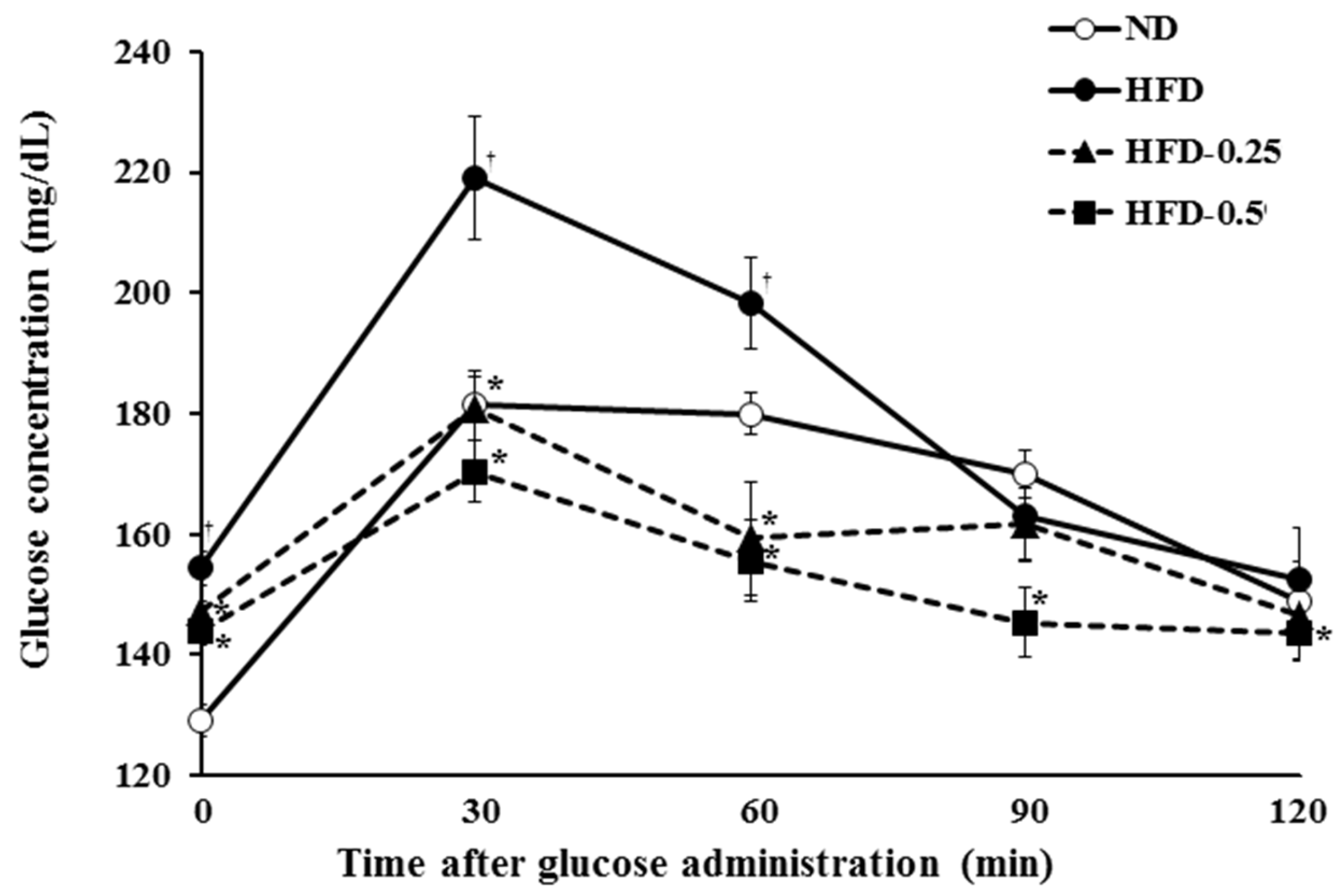
3.6. Effects of NAOs on Insulin Resistance
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Araki, C.H. Acetylation of agar like substance of Gelidium amansii. J. Chem. Soc. 1937, 58, 1338–1350. [Google Scholar]
- Giordano, A.; Andreotti, G.; Tramice, A.; Trincone, A. Marine glycosyl hydrolases in the hydrolysis and synthesis of oligosaccharides. Biotechnol. J. 2006, 1, 511–530. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.J.; Chang, Y.K.; Hong, S.K. Agar degradation by microorganisms and agar-degrading enzymes. Appl. Microbiol. Biotechnol. 2012, 94, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Potin, P.; Richard, C.; Rochas, C.; Kloareg, B. Purification and characterization of the α-agarase from Alteromonas agarlyticus (Cataldi) comb. nov., strain GJ1B. Eur. J. Biochem. 1993, 214, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Masuda, N.; Iwasaki, Y.; Nakagawa, H.; Kobayashi, R.; Usami, S. Purification and characterization of a novel β-agarase from an alkalophilic bacterium, Alteromonas sp. E-1. J. Biosci. Bioeng. 1999, 87, 436–441. [Google Scholar] [CrossRef]
- Park, J.; Hong, S.K.; Chang, Y.K. Production of DagA, a β-agarase, by Streptomyces lividans in glucose medium or mixed-sugar medium simulating microalgae hydrolysate. J. Microbiol. Biotechnol. 2014, 24, 1622–1628. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Kim, S.M. Agarase: Review of major sources, categories, purification method, enzyme characteristics and applications. Mar. Drugs 2010, 8, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Enoki, T.; Tominaga, T.; Takashima, F.; Ohnogi, H.; Sagawa, H.; Kato, I. Anti-tumor-promoting activities of agaro-oligosaccharides on two-stage mouse skin carcinogenesis. Biol. Pharm. Bull. 2012, 35, 1145–1149. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yan, X.; Zhu, P.; Lin, J. Antioxidant activity and hepatoprotective potential of agaro-oligosaccharides in vitro and in vivo. Nutr. J. 2006, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, R.; Takisada, M.; Suzuki, T.; Kirimura, K.; Usami, S. Neoagarobiose as a novel moisturizer with whitening effect. Biosci. Biotechnol. Biochem. 1997, 61, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Hatada, Y.; Nogi, Y.; Li, Z.; Ito, S.; Horikoshi, K. Cloning, expression, and characterization of a glycoside hydrolase family 86 β-agarase from a deep-sea Microbulbifer-like isolate. Appl. Microbiol. Biotechnol. 2004, 66, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Barness, L.A.; Opitz, J.M.; Gilbert-Barness, E. Obesity: Genetic, molecular, and environmental aspects. Am. J. Med. Genet. Part A 2007, 143, 3016–3034. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.G.; Apovian, C.M.; Aronne, L.J. New drug targets for the treatment of obesity. Clin. Pharmacol. Ther. 2011, 90, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.W.; Kim, Y.T.; Jang, Y.J.; Kim, Y.O.; Han, D.S. Anti-obesity effect of Artemisia capillaris extracts in high-fat diet-induced obese rats. Molecules 2013, 18, 9241–9252. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef] [PubMed]
- Temuujin, U.; Lee, S.Y.; Chang, Y.K.; Chi, W.J.; Hong, S.K. Overexpression and biochemical characterization of DagA from Streptomyces coelicolor A3(2): An endo-type β-agarase producing neoagarotetraose and neoagarohexaose. Appl. Microbiol. Biotechnol. 2011, 92, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Kieser, H.; Bibb, M.J.; Buttner, M.J.; Chater, F.K.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Kleiner, D.E.; Brunt, E.M.; Natta, M.V.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W.S. ImageJ; U.S. National Institutes of Health: Bethesda, MD, USA, 1997–2016. Available online: https://imagej.nih.gov/ij/ (accessed on 23 March 2017).
- Nolan, J.J.; Ludvik, B.; Baloga, J.; Reichart, D.; Olefsky, J.M. Mechanisms of the kinetic defect in insulin action in obesity and NIDDM. Diabetes 1997, 46, 994–1000. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Kim, S.K. Potential immune-stimulating effect of antitumoral fraction of chitosan oligosaccharides. J. Chitin Chitosan Sci. 2001, 6, 163–167. [Google Scholar]
- Kim, M.H.; Kim, H.Y.; Kim, W.K.; Kim, J.Y.; Kim, S.H. Effects of soyoligosaccharides on blood glucose and lipid metabolism in streptozotocin-induced diabetic rats. J. Korean Nutr. Soc. 2001, 34, 3–13. [Google Scholar]
- Hsieh, P.C.; Huang, G.J.; Ho, Y.L.; Lin, Y.H.; Huang, S.S.; Chiang, Y.C.; Tseng, M.C.; Chang, Y.S. Activities of antioxidants, α-glucosidase inhibitors and aldose reductase inhibitors of the aqueous extracts of four Flemingia species in Taiwan. Bot. Stud. 2010, 51, 293–302. [Google Scholar]
- Hong, S.J.; Lee, J.-H.; Kim, E.J.; Yang, H.J.; Chang, Y.-K.; Park, J.-S.; Hong, S.-K. In vitro and in vivo investigation for biological activities of neoagarooligosaccharides prepared by hydrolysis agar with β-agarase. Biotechnol. Bioprocess Eng. 2017. in revision. [Google Scholar]
- Haque, M.; Sanyal, A.J. The metabolic abnormalities associated with non-alcoholic fatty liver disease. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 709–731. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, D.; Borkan, G.A.; Gerzof, S.G. Relationship of fat distribution to glucose tolerance. Diabetes 1986, 35, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M.S.; Ahrén, B. The high-fat diet-fed mouse a model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53, S215–S219. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Seeley, R.J.; Rushing, P.A.; D’Alessio, D.; Tso, P. A controlled high-fat diet induces an obese syndrome in rats. J. Nutr. 2003, 133, 1081–1087. [Google Scholar] [PubMed]
- Reddy, J.K.; Rao, M.S. Lipid metabolism and liver inflammation. II. Fatty liver disease and fatty acid oxidation. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G852–G858. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Wang, Y.; Keshaw, H.; Xu, L.Y.; Lam, K.S.; Cooper, G.J. The fat-derived hormone adiponectin alleviates alcoholic and nonalcoholic fatty liver diseases in mice. J. Clin. Investig. 2003, 112, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M.; Langin, D. Lipolysis and lipid mobilization in human adipose tissue. Prog. Lipid Res. 2009, 48, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Loncar, D.; Afzelius, B.A.; Cannon, B. Epididymal white adipose tissue after cold stress in rats. I. Nonmitochondrial changes. J. Ultrastruct. Mol. Struct. Res. 1988, 101, 109–122. [Google Scholar] [CrossRef]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or hyperplasia: Dynamics of adipose tissue growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef] [PubMed]
- Díez, J.J.; Iglesias, P. The role of the novel adipocyte-derived hormone adiponectin in human disease. Eur. J. Endocrinol. 2003, 148, 293–300. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg) | ND | HFD | HFD-0.25 | HFD-0.5 |
|---|---|---|---|---|
| Casein | 210.0 | 230.0 | 230.0 | 230.0 |
| l-Cystine | 3.0 | 3.5 | 3.5 | 3.5 |
| Maltodextrin | 50.0 | 160.0 | 160.0 | 160.0 |
| Sucrose | 325.0 | 90.0 | 90.0 | 90.0 |
| Lard | 20.0 | 310.0 | 310.0 | 310.0 |
| Soybean oil | 20.0 | 30.0 | 30.0 | 30.0 |
| Cellulose | 37.15 | 65.5 | 65.5 | 65.5 |
| Corn starch | 280.0 | - | - | - |
| Mineral mix | 35.0 | 48.0 | 48.0 | 48.0 |
| CaH(PO4)2 | 2.0 | 3.4 | 3.4 | 3.4 |
| Vitamin mix | 15.0 | 21.0 | 21.0 | 21.0 |
| Choline bitartrate | 2.75 | 3.0 | 3.0 | 3.0 |
| Blue food color | 0.1 | 0.1 | 0.1 | 0.1 |
| NAOs | - | - | 2.5 | 5 |
| Nutrition facts: % g (w/w) (% kcal) | ||||
| Protein | 18.8 (20.1) | 23.5 (18.4) | 23.5 (18.4) | 23.5 (18.4) |
| Carbohydrate | 64.7 (69.8) | 27.3 (21.3) | 27.3 (21.3) | 27.3 (21.3) |
| Fat | 4.2 (10.2) | 34.3 (60.3) | 34.3 (60.3) | 34.3 (60.3) |
| Kcal/g | 3.6 | 5.1 | 5.1 | 5.1 |
| Group | Body Weight (g) | Body Weight Gain (g/64 Days) | Food Intake (g/Day) | FER 2 | |
|---|---|---|---|---|---|
| Initial | Final 1 | ||||
| ND | 21.13 ± 0.27 | 29.35 ± 0.27 | 8.22 ± 0.19 | 4.15 ± 0.09 | 2.02 ± 0.07 |
| HFD | 21.30 ± 0.30 | 37.78 ± 1.04 a | 16.48 ± 0.85 a | 2.74 ± 0.05 a | 6.40 ± 0.51 a |
| HFD-0.25 | 21.00 ± 0.25 | 37.59 ± 0.63 | 16.59 ± 0.58 | 2.56 ± 0.10 | 6.45 ± 0.36 |
| HFD-0.5 | 20.89 ± 0.27 | 34.37 ± 0.97 b | 13.48 ± 0.94 b | 2.77 ± 0.01 | 4.82 ± 0.36 b |
| Group | Adipocyte Size (Pixel × 103) | Perirenal Adipose Tissue (g) | Epididymal Adipose Tissue (g) |
|---|---|---|---|
| ND | 4.64 ± 0.54 | 0.215 ± 0.035 | 0.222 ± 0.031 |
| HFD | 10.74 ± 1.44 a | 0.920 ± 0.329 a | 0.902 ± 0.312 a |
| HFD-0.25 | 6.74 ± 0.38 b | 0.929 ± 0.149 | 0.923 ± 0.201 |
| HFD-0.5 | 6.95 ± 0.82 b | 0.824 ± 0.413 | 0.797 ± 0.370 |
| Group | Total Cholesterol (mg/dL) | Triglyceride (mg/dL) | Free Fatty Acid (mEq/L 1) |
|---|---|---|---|
| ND | 140.1 ± 5.2 | 48.7 ± 3.2 | 1.28 ± 0.07 |
| HFD | 182.0 ± 8.2 a | 64.8 ± 4.0 a | 1.52 ± 0.11 a |
| HFD-0.25 | 173.0 ± 4.3 | 56.7 ± 4.4 | 1.30 ± 0.05 b |
| HFD-0.5 | 159.6 ± 5.6 b | 47.3 ± 2.9 b | 1.22 ± 0.06 b |
| Group | Area under Curve (mg, 0–120 min) | Insulin (ng/mL) | Glucose (mg/dL) | Adiponectin (ng/mL) |
|---|---|---|---|---|
| ND | 332.3 ± 6.8 | 0.72 ± 0.12 | 115.4 ± 7.3 | 5.01 ± 0.79 |
| HFD | 373.1 ± 12.4 a | 1.35 ± 0.14 a | 153.3 ± 7.3 a | 4.81 ± 1.11 |
| HFD-0.25 | 322.4 ± 11.6 b | 0.92 ± 0.13 b | 159.4 ± 4.0 | 5.63 ± 0.81 |
| HFD-0.5 | 308.0 ± 10.6 b | 0.94 ± 0.14 | 156.8 ± 19.6 | 6.13 ± 1.25 b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.J.; Lee, J.-H.; Kim, E.J.; Yang, H.J.; Park, J.-S.; Hong, S.-K. Anti-Obesity and Anti-Diabetic Effect of Neoagarooligosaccharides on High-Fat Diet-Induced Obesity in Mice. Mar. Drugs 2017, 15, 90. https://doi.org/10.3390/md15040090
Hong SJ, Lee J-H, Kim EJ, Yang HJ, Park J-S, Hong S-K. Anti-Obesity and Anti-Diabetic Effect of Neoagarooligosaccharides on High-Fat Diet-Induced Obesity in Mice. Marine Drugs. 2017; 15(4):90. https://doi.org/10.3390/md15040090
Chicago/Turabian StyleHong, Sun Joo, Je-Hyeon Lee, Eun Joo Kim, Hea Jung Yang, Jae-Seon Park, and Soon-Kwang Hong. 2017. "Anti-Obesity and Anti-Diabetic Effect of Neoagarooligosaccharides on High-Fat Diet-Induced Obesity in Mice" Marine Drugs 15, no. 4: 90. https://doi.org/10.3390/md15040090
APA StyleHong, S. J., Lee, J. -H., Kim, E. J., Yang, H. J., Park, J. -S., & Hong, S. -K. (2017). Anti-Obesity and Anti-Diabetic Effect of Neoagarooligosaccharides on High-Fat Diet-Induced Obesity in Mice. Marine Drugs, 15(4), 90. https://doi.org/10.3390/md15040090