
Higher Anti-Liver Fibrosis Effect of Cordyceps militaris-Fermented Product Cultured with Deep Ocean Water via Inhibiting Proinflammatory Factors and Fibrosis-Related Factors Expressions



Abstract
:1. Introduction
2. Results
2.1. Effect of DOW on the Cordycepin and Adenosine Production of CM
2.2. Effect of the Individual Metal Waters on the Production of Monascus-Fermented Pigments
2.3. Effect on Serum AST, ALT, ALP, and TBIL
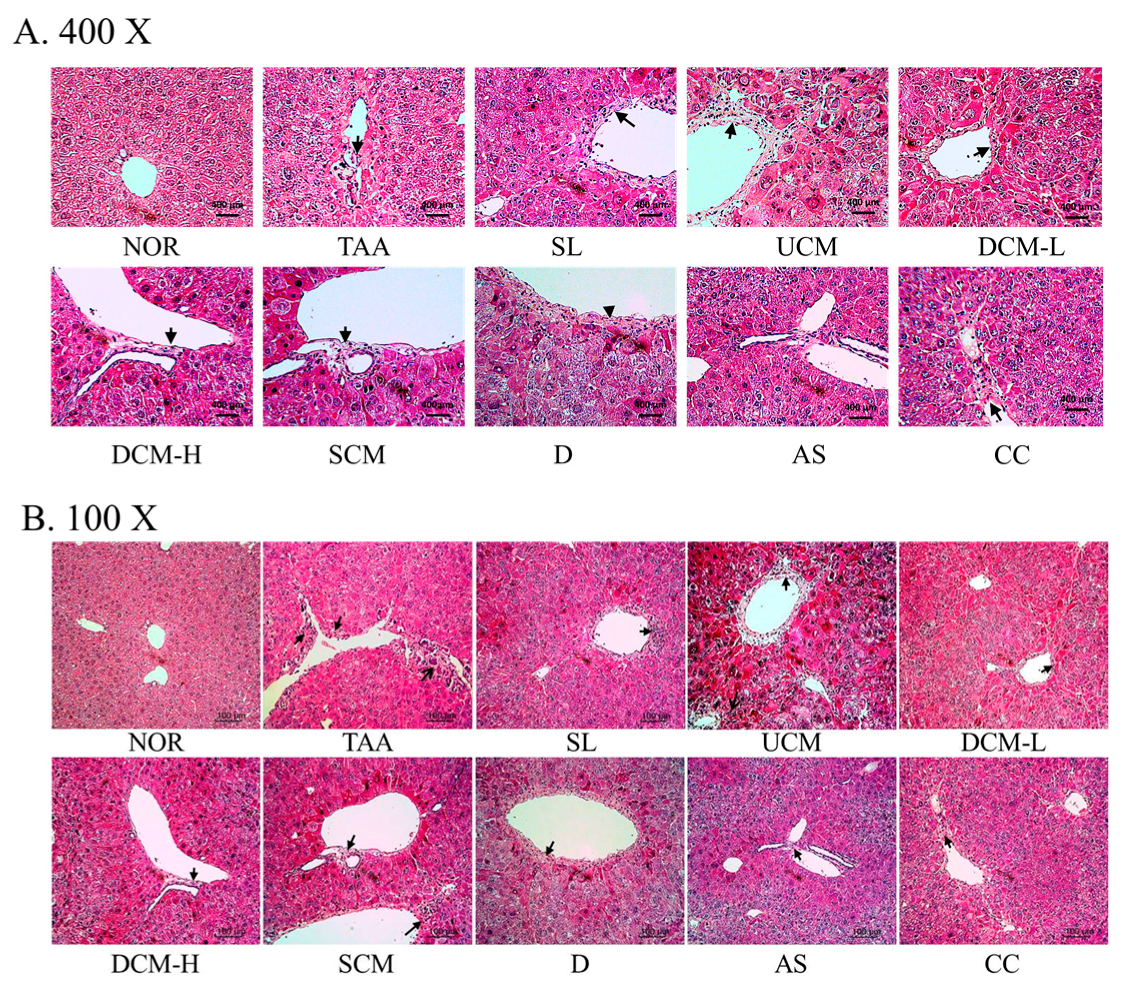
2.4. Effect on the Liver HE Stain and Collagen Stain
2.5. Effect on the Liver Proinflammatory Factors and PPAR-γ Protein Expression
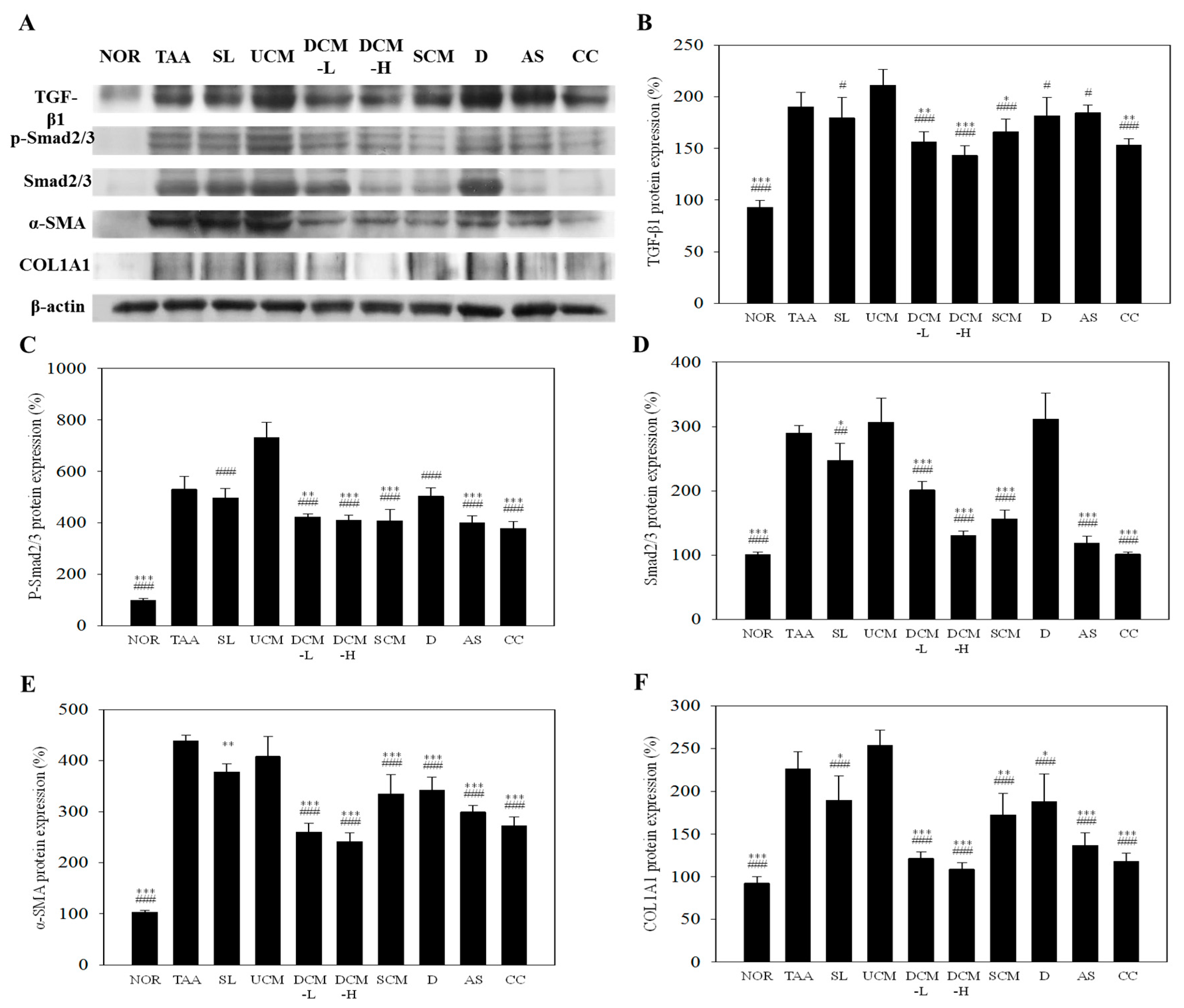
2.6. Effect on the Liver Fibrosis-Related Factors Expression
3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. The Source of DOW
4.3. Microorganism and Seed Cultures
4.4. Solid Fermentation of C. militaris in DOW, SW, or UPW
4.5. Cordycepin and Adenosine Analysis
4.6. Animal Experiments
4.7. Serum Biochemical and Fibrosis Markers Analyses
4.8. Immunoblotting
4.9. Histological Analysis and Collagen Staining
4.10. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Friedman, S.L. Hepatic stellate cells: Protean, multifunctional, and enigmatic cells of the liver. Physiol. Rev. 2008, 88, 125–172. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Hasegawa, G.; Ebe, Y.; Yamamoto, T. Differentiation and function of Kupffer cells. Med. Electron. Microsc. 2004, 37, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Ceni, E.; Salzano, R.; Biondi, P.; Parola, M.; Galli, A.; Foschi, M.; Caligiuri, A.; Pinzani, M.; Surrenti, C. Neutrophil-derived superoxide anion induces lipid peroxidation and stimulates collagen synthesis in human hepatic stellate cells: Role of nitric oxide. Hepatology 1997, 25, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Shek, F.W.; Benyon, R.C. How can transforming growth factor β be targeted usefully to combat liver fibrosis? Eur. J. Gastroenterol. Hepatol. 2004, 16, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Olaso, E.; Friedman, S.L. Molecular regulation of hepatic fibrogenesis. J. Hepatol. 1998, 29, 836–847. [Google Scholar] [CrossRef]
- Hataguchi, Y.; Tai, H.; Nakajima, H.; Kimata, H. Drinking deep-sea water restores mineral imbalance in atopic eczema/dermatitis syndrome. Eur. J. Clin. Nutr. 2005, 59, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Kim, H.A.; Lee, S.H.; Yun, J.W. Anti-obesity and antidiabetic effects of deep sea water on ob/ob mice. Mar. Biotechnol. 2009, 11, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Kung, Y.H.; Wang, J.J.; Lung, T.Y.; Pan, T.M. Enhanced hypolipidemic effect and safety of red mold dioscorea cultured in deep ocean water. J. Agric. Food Chem. 2011, 59, 8199–8207. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Kuo, I.U.; Tsai, T.Y.; Lee, C.L. Antrodia camphorata-fermented product cultured in deep ocean water has more liver protection against thioacetamide-induced fibrosis. Appl. Microbiol. Biotechnol. 2013, 97, 9955–9967. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Lung, T.Y.; Kung, Y.H.; Wang, J.J.; Tsai, T.Y.; Wei, B.L.; Pan, T.M.; Lee, C.L. Enhanced anti-obesity activities of red mold dioscorea when fermented using deep ocean water as the culture water. Mar. Drugs 2013, 11, 3902–3925. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Naura, A.S.; Errami, Y.; Ju, J.H.; Boulares, A.H. Cordycepin Blocks Lung Injury-Associated Inflammation and Promotes BRCA1-Deficient Breast Cancer Cell Killing by Effectively Inhibiting PARP. Mol. Med. 2011, 17, 893–900. [Google Scholar] [PubMed]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H.I.; et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Johno, H.; Nakajima, S.; Kato, H.; Takahashi, S.; Katoh, R.; Kitamura, M. Blockade of Smad signaling by 3′-deoxyadenosine: A mechanism for its anti-fibrotic potential. Lab. Investig. 2013, 93, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L. The advantages of deep ocean water for the development of functional fermentation food. Appl. Microbiol. Biotechnol. 2015, 99, 2523–2531. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, S.K. A toxicologist guide to the diagnostic interpretation of hepatic biochemical parameters. Food Chem. Toxicol. 2007, 45, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Marra, F. Chemokines in liver inflammation and fibrosis. Front. Biosci. 2002, 7, d1899–d1914. [Google Scholar] [CrossRef] [PubMed]
- Bonniaud, P.; Margetts, P.J.; Ask, K.; Flanders, K.; Gauldie, J.; Kolb, M. TGF-β and Smad3 signaling link inflammation to chronic fibrogenesis. J. Immunol. 2005, 175, 5390–5395. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Chiu, Y.T.; Cheng, C.C.; Wu, C.F.; Lin, Y.L.; Huang, Y.T. Antifibrotic effects of tetrandrine on hepatic stellate cells and rats with liver fibrosis. J. Gastroenterol. Hepatol. 2007, 22, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; Schoonhoven, R.; Tuvia, S.; Brenner, D.A.; Rippe, R.A. Nuclear factor κB in proliferation, activation, and apoptosis in rat hepatic stellate cells. J. Hepatol. 2000, 33, 49–58. [Google Scholar] [CrossRef]
- Castrillo, A.; Tontonoz, P. Nuclear receptors in macrophage biology: At the crossroads of lipid metabolism and inflammation. Annu. Rev. Cell Dev. Biol. 2004, 20, 455–480. [Google Scholar] [CrossRef] [PubMed]
- Marra, F.; Efsen, E.; Romanelli, R.G.; Caligiuri, A.; Pastacaldi, S.; Batignani, G.; Bonacchi, A.; Caporale, R.; Laffi, G.; Pinzani, M.; et al. Ligands of peroxisome proliferator-activated receptor γ modulate profibrogenic and proinflammatory actions in hepatic stellate cells. Gastroenterology 2000, 119, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Crabb, D.W.; Ceni, E.; Salzano, R.; Mello, T.; Svegliati-Baroni, G.; Ridolfi, F.; Trozzi, L.; Surrenti, C.; Casini, A. Antidiabetic thiazolidinediones inhibit collagen synthesis and hepatic stellate cell activation in vivo and in vitro. Gastroenterology 2002, 122, 1924–1940. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; de Crombrugghe, B. Conservation of binding sites for regulatory factors in the coordinately expressed α1 (I) and α2 (I) collagen promoters. Biochem. Biophys. Res. Commun. 1991, 177, 538–544. [Google Scholar] [CrossRef]
- Cutroneo, K.R.; White, S.L.; Phan, S.H.; Ehrlich, H.P. Therapies for bleomycin induced lung fibrosis through regulation of TGF-β1 induced collagen gene expression. J. Cell Physiol. 2007, 211, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Benjamin, I.S.; Alexander, B. Reproducible production of thioacetamide-induced macronodular cirrhosis in the rat with no mortality. J. Hepatol. 2002, 36, 488–493. [Google Scholar] [CrossRef]
- Torres, M.I.; Fernandez, M.I.; Gil, A.; Rios, A. Dietary nucleotides have cytoprotective properties in rat liver damaged by thioacetamide. Life Sci. 1998, 62, 13–22. [Google Scholar] [CrossRef]
- Liang, T.J.; Yuan, J.H.; Tan, Y.R.; Ren, W.H.; Han, G.Q.; Zhang, J.; Wang, L.C.; Qin, C.Y. Effect of ursodeoxycholic acid on TGF β1/Smad signaling pathway in rat hepatic stellate cells. Chin. Med. J. 2009, 122, 1209–1213. [Google Scholar] [PubMed]
- Cha, J.Y.; Ahn, H.Y.; Cho, Y.S.; Je, J.Y. Protective effect of cordycepin-enriched Cordyceps militaris on alcoholic hepatotoxicity in Sprague-Dawley rats. Food Chem. Toxicol. 2013, 60, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.H.; Wing, E.J. Neutrophil-Kupffer-cell interaction in host defenses to systemic infections. Immunol. Today 1998, 19, 507–510. [Google Scholar] [CrossRef]
- Kang, J.W.; Yoon, S.J.; Sung, Y.K.; Lee, S.M. Magnesium chenoursodeoxycholic acid ameliorates carbon tetrachloride-induced liver fibrosis in rats. Exp. Biol. Med. 2012, 237, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, R.X.; Wang, F.G.; Zhou, H.P.; Shi, T.T.; Liu, S.R. The protective effect of N-acetylcysteine magnesium against liver cirrhosis with portal hypertension in rat. Chin. J. Exp. Clin. Virol. 2012, 26, 366–369. [Google Scholar]
- Mogna, L.; Nicola, S.; Pane, M.; Lorenzini, P.; Strozzi, G.; Mogna, G. Selenium and zinc internalized by Lactobacillus buchneri Lb26 (DSM 16341) and Bifidobacterium lactis Bb1 (DSM 17850): Improved bioavailability using a new biological approach. J. Clin. Gastroenterol. 2012, 46, S41–S45. [Google Scholar] [CrossRef] [PubMed]
- Kwak, K.G.; Wang, J.H.; Shin, J.W.; Lee, D.S.; Son, C.G. A traditional formula, Chunggan extract, attenuates thioacetamide-induced hepatofibrosis via GSH system in rats. Hum. Exp. Toxicol. 2011, 30, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E. The Growth of the Surface Area of Human Body; University of Minnesota Press: Minneapolis, MN, USA, 1935. [Google Scholar]
- Wang, L.C.; Wang, S.E.; Wang, J.J.; Tsai, T.Y.; Lin, C.H.; Pan, T.M.; Lee, C.L. In vitro and in vivo comparisons of the effects of the fruiting body and mycelium of Antrodia camphorata against amyloid β-protein-induced neurotoxicity and memory impairment. Appl. Microbiol. Biotechnol. 2012, 94, 1505–1519. [Google Scholar] [CrossRef] [PubMed]
- Bihaqi, S.W.; Singh, A.P.; Tiwari, M. Supplementation of Convolvulus pluricaulis attenuates scopolamine-induced increased tau and Amyloid precursor protein (AβPP) expression in rat brain. Indian J. Pharmacol. 2012, 44, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Kuo, T.F.; Wu, C.L.; Wang, J.J.; Pan, T.M. Red mold rice promotes neuroprotective sAPPalpha secretion instead of Alzheimer’s risk factors and amyloid beta expression in hyperlipidemic Aβ40-infused rats. J. Agric. Food Chem. 2010, 58, 2230–2238. [Google Scholar] [CrossRef] [PubMed]
- Phadnis, S.V.; Atilade, A.; Bowring, J.; Kyrgiou, M.; Young, M.P.; Evans, H.; Paraskevaidis, E.; Walker, P. Regeneration of cervix after excisional treatment for cervical intraepithelial neoplasia: A study of collagen distribution. BJOG 2011, 118, 1585–1591. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Cordycepin Concentration (mg/g) | Adenosine Concentration (mg/g) |
|---|---|---|
| UCM | 505.12 ± 16.91 *** | 235.18 ± 77.26 *** |
| DCM | 1020.45 ± 76.42 | 544.30 ± 24.45 |
| SCM | 820.60 ± 75.25 * | 413.08 ± 10.17 * |
| Groups | Liver Weight Gain (g) | Body Weight (g) | Liver Weight/Body Weight Ratio (%) |
|---|---|---|---|
| NOR | 1.24 ± 0.06 ***, ### | 29.36 ± 1.17 ***, ### | 4.51 ± 0.14 ***, ### |
| TAA | 2.07 ± 0.09 ### | 23.34 ± 1.05 ### | 7.88 ± 0.19 ### |
| SL | 1.95 ± 0.07 *** | 27.35 ± 1.03 *** | 7.15 ± 0.24 *** |
| UCM | 1.91 ± 0.08 *** | 27.31 ± 0.46 *** | 7.20 ± 0.24 *** |
| DCM-L | 1.90 ± 0.06 *** | 27.88 ± 0.59 *** | 6.98 ± 0.36 *** |
| DCM-H | 1.93 ± 0.04 *** | 27.26 ± 1.15 *** | 7.09 ± 0.31 *** |
| SCM | 1.81 ± 0.04 ***, ## | 25.79 ± 0.55 *** | 7.10 ± 0.18 *** |
| DOW | 1.89 ± 0.06 *** | 26.96 ± 0.82 *** | 7.17 ± 0.15 *** |
| AS | 1.69 ± 0.08 ***, ### | 25.76 ± 0.93 *** | 7.10 ± 0.22 *** |
| CC | 1.75 ± 0.05 ***, ### | 25.01 ± 0.87 *** | 6.95 ± 0.24 *** |
| Groups | AST (U/L) | ALT (U/L) | ALP(U/L) | TBIL (mg/dL) |
|---|---|---|---|---|
| NOR | 80.63 ± 3.89 ***, ## | 43.25 ± 2.55 ***, ### | 97.63 ± 4.93 ***, ### | 0.043 ± 0.007 ***, ### |
| TAA | 124.50 ± 11.12 ### | 138.75 ± 10.12 | 202.63 ± 7.48 ### | 0.109 ± 0.010 ## |
| SL | 94.38 ± 4.84 *** | 114.75 ± 5.68 * | 175.00 ± 16.29 *** | 0.091 ± 0.010 *** |
| UCM | 92.25 ± 3.45 *** | 126.63 ± 12.23 | 170.63 ± 8.90 *** | 0.094 ± 0.005 ** |
| DCM-L | 89.25 ± 5.57 ***, # | 128.88 ± 10.79 | 174.63 ± 13.71 *** | 0.086 ± 0.009 *** |
| DCM-H | 90.75 ± 6.92 *** | 119.63 ± 7.44 * | 171.63 ± 14.09 *** | 0.086 ± 0.009 *** |
| SCM | 97.63 ± 9.02 *** | 129.88 ± 14.51 | 176.13 ± 5.30 *** | 0.086 ± 0.012 *** |
| D | 96.25 ± 8.97 *** | 128.63 ± 13.90 | 177.13 ± 9.43 *** | 0.095 ± 0.009 ** |
| AS | 99.00 ± 4.78 *** | 125.38 ± 17.73 * | 161.75 ± 11.52 *** | 0.071 ± 0.011 ***, ### |
| CC | 97.13 ± 10.59 *** | 106.75 ± 10.50 *, ## | 176.38 ± 10.01 *** | 0.080 ± 0.011 ***, # |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, Y.-P.; Lee, C.-L. Higher Anti-Liver Fibrosis Effect of Cordyceps militaris-Fermented Product Cultured with Deep Ocean Water via Inhibiting Proinflammatory Factors and Fibrosis-Related Factors Expressions. Mar. Drugs 2017, 15, 168. https://doi.org/10.3390/md15060168
Hung Y-P, Lee C-L. Higher Anti-Liver Fibrosis Effect of Cordyceps militaris-Fermented Product Cultured with Deep Ocean Water via Inhibiting Proinflammatory Factors and Fibrosis-Related Factors Expressions. Marine Drugs. 2017; 15(6):168. https://doi.org/10.3390/md15060168
Chicago/Turabian StyleHung, Yu-Ping, and Chun-Lin Lee. 2017. "Higher Anti-Liver Fibrosis Effect of Cordyceps militaris-Fermented Product Cultured with Deep Ocean Water via Inhibiting Proinflammatory Factors and Fibrosis-Related Factors Expressions" Marine Drugs 15, no. 6: 168. https://doi.org/10.3390/md15060168
APA StyleHung, Y. -P., & Lee, C. -L. (2017). Higher Anti-Liver Fibrosis Effect of Cordyceps militaris-Fermented Product Cultured with Deep Ocean Water via Inhibiting Proinflammatory Factors and Fibrosis-Related Factors Expressions. Marine Drugs, 15(6), 168. https://doi.org/10.3390/md15060168