Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus

 ,
,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
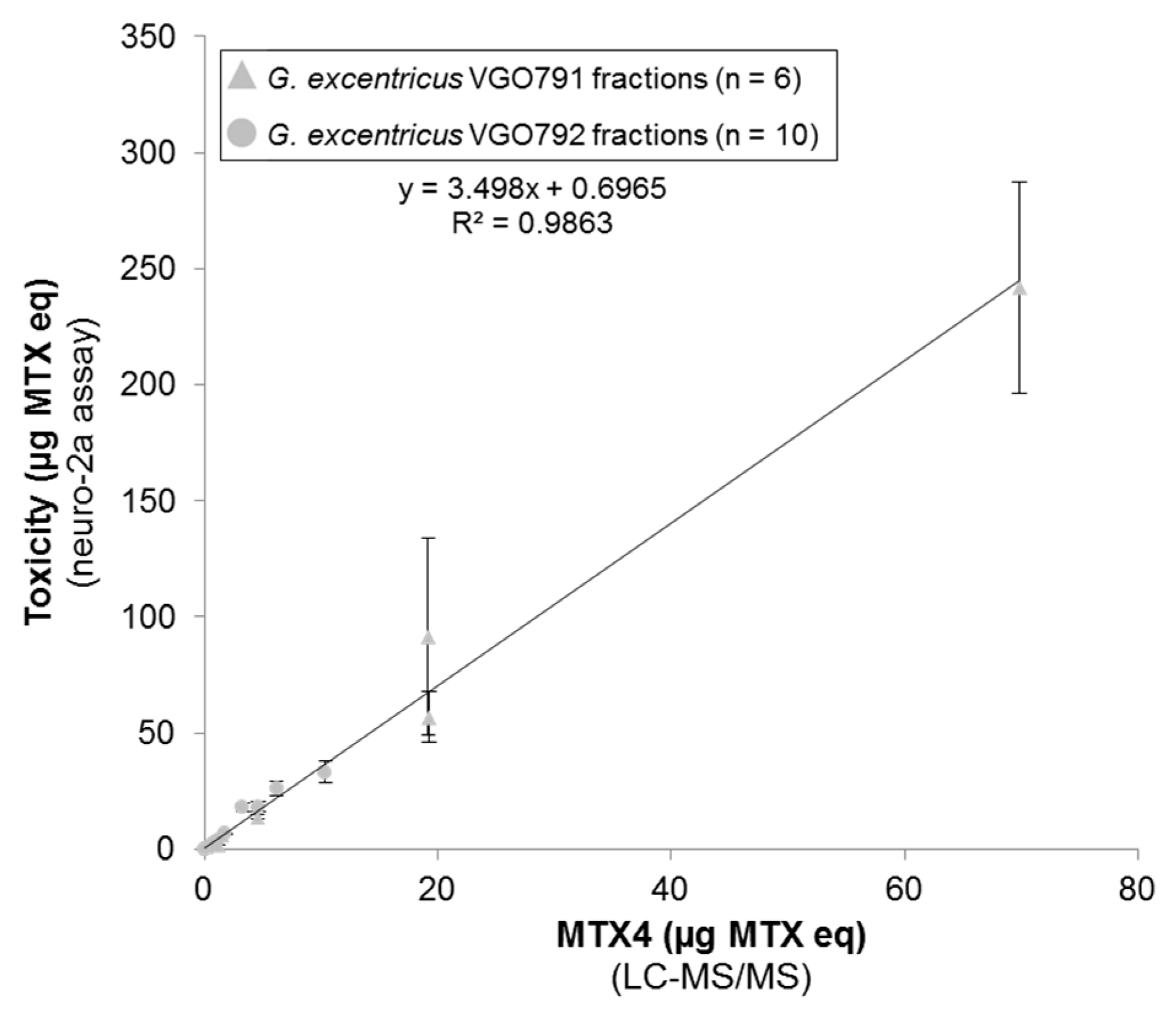
2.1. Toxicity of the Aqueous Methanol Fractions (MSFs) from G. excentricus Strains VGO791 and VGO792
2.2. Screening of Fractionated G. excentricus Extracts Using Neuroblastoma N2a Assays
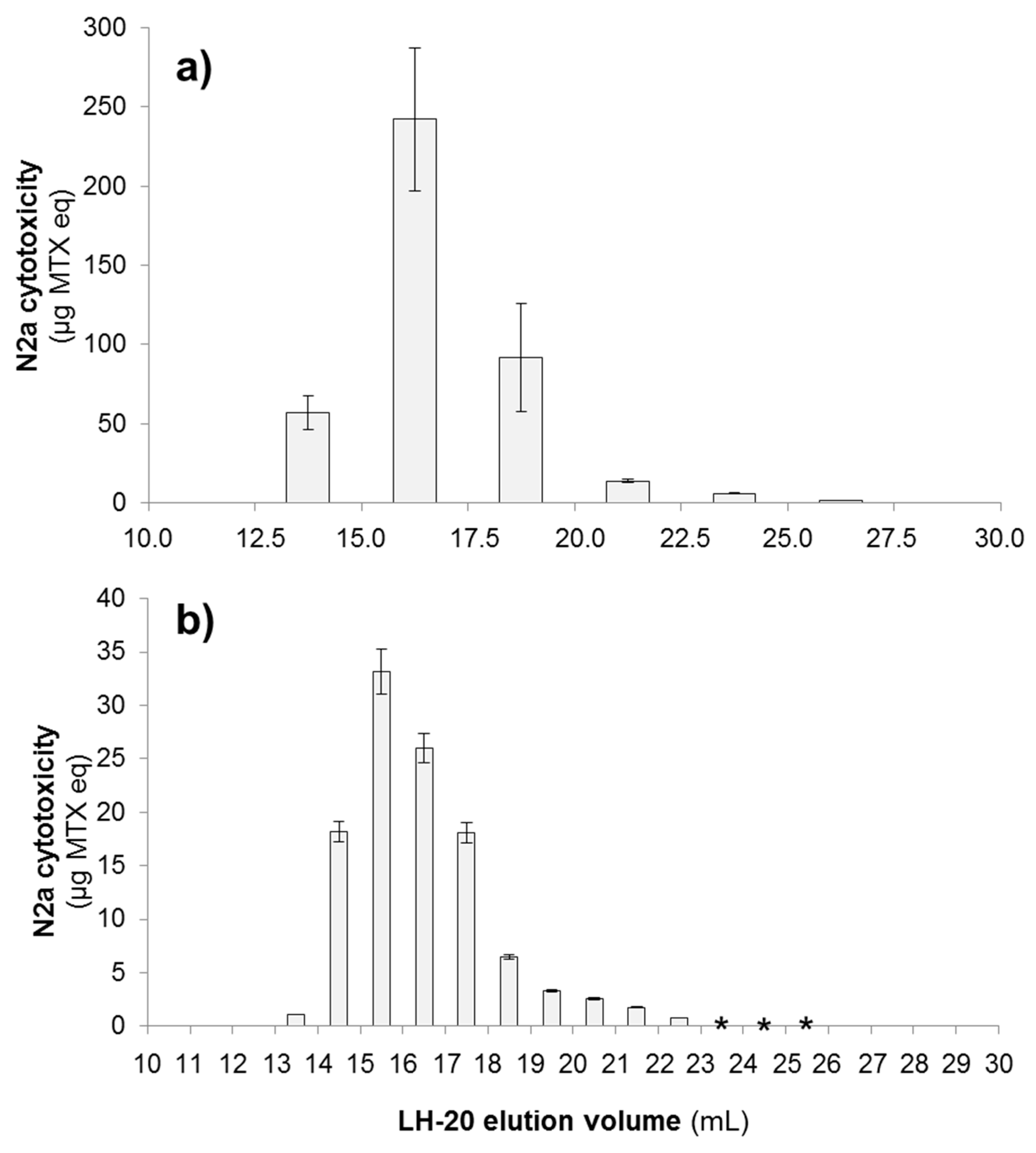
2.2.1. N2a Cytotoxicity Assay
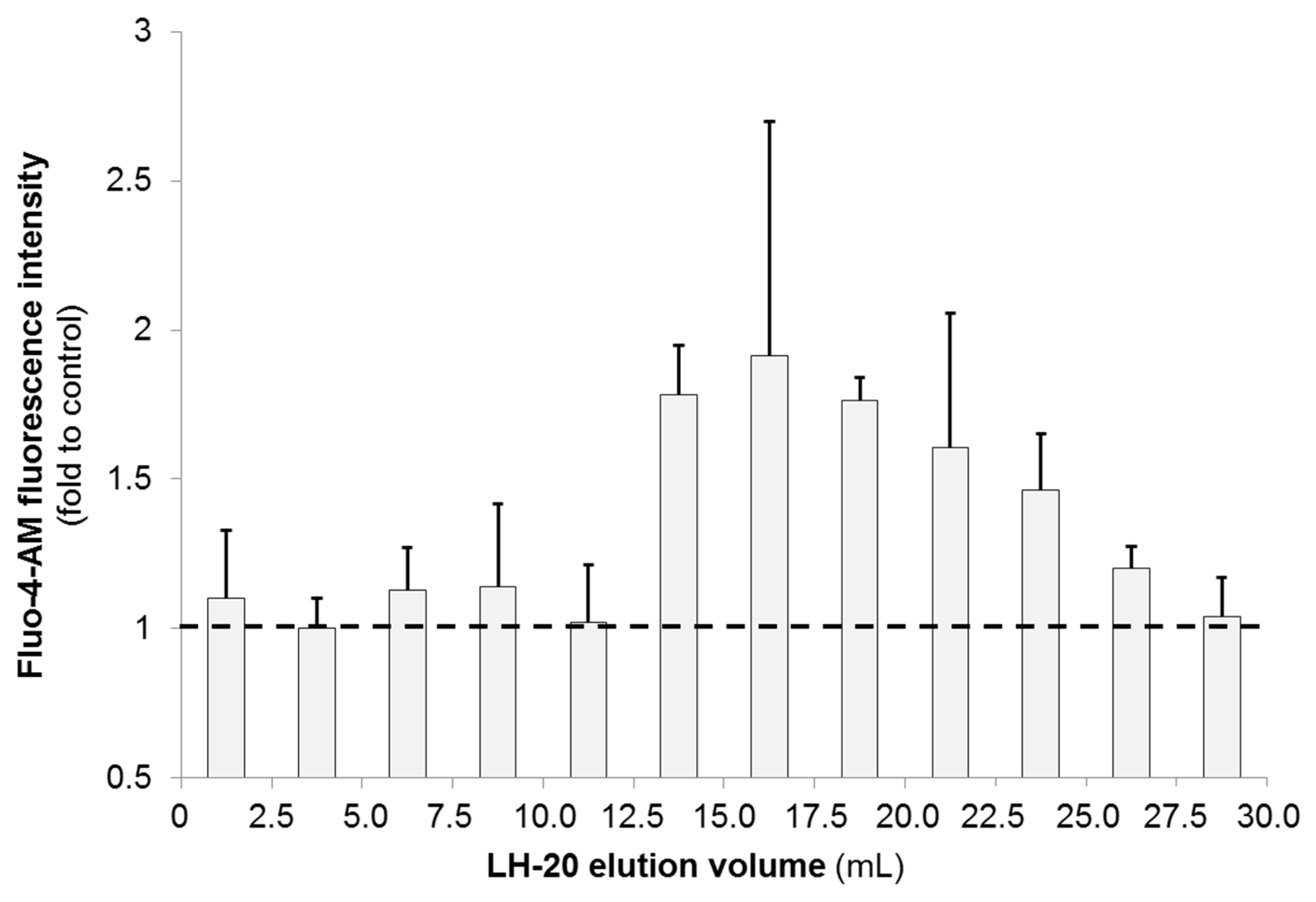
2.2.2. N2a Calcium Flux Assay
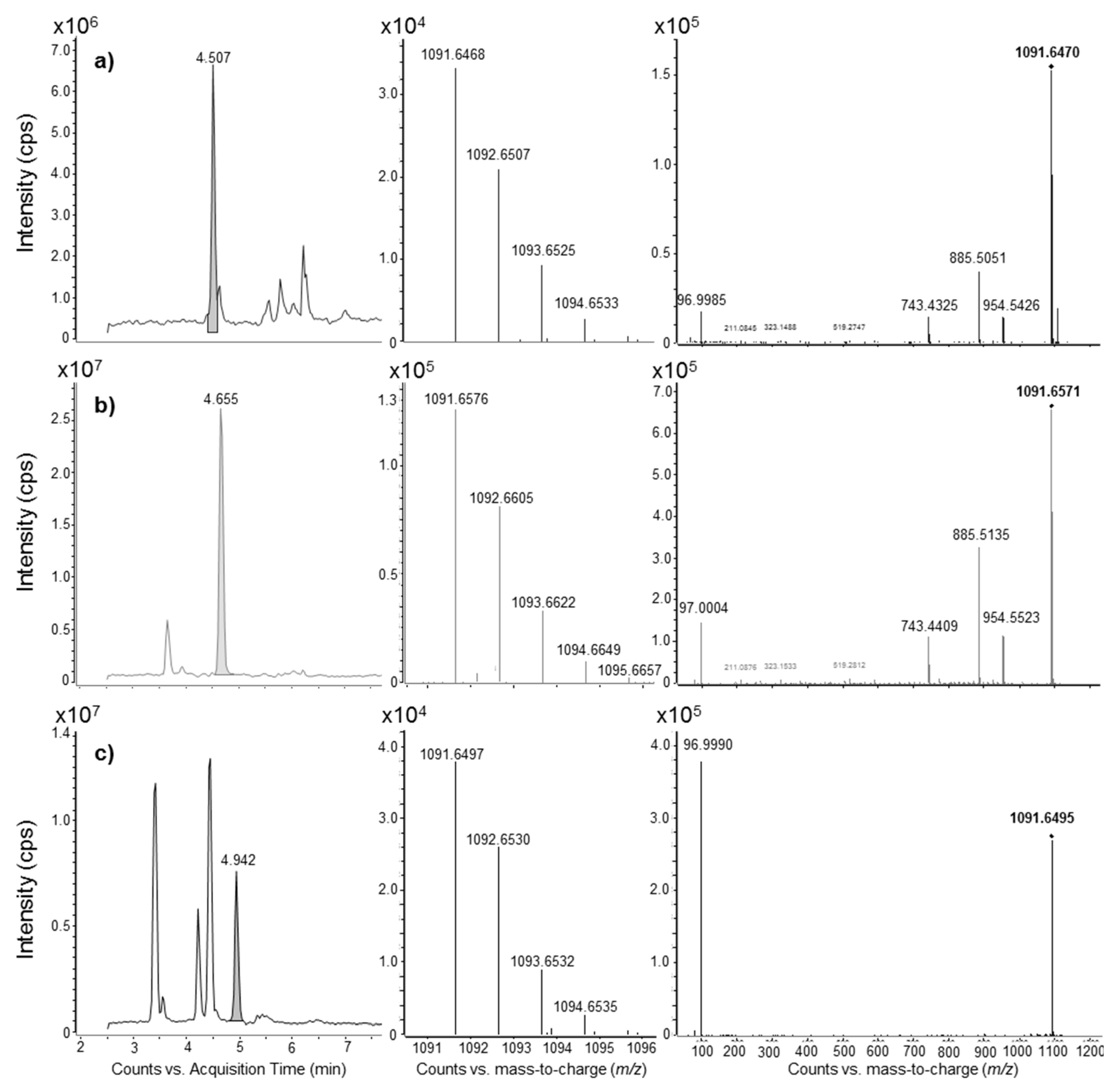
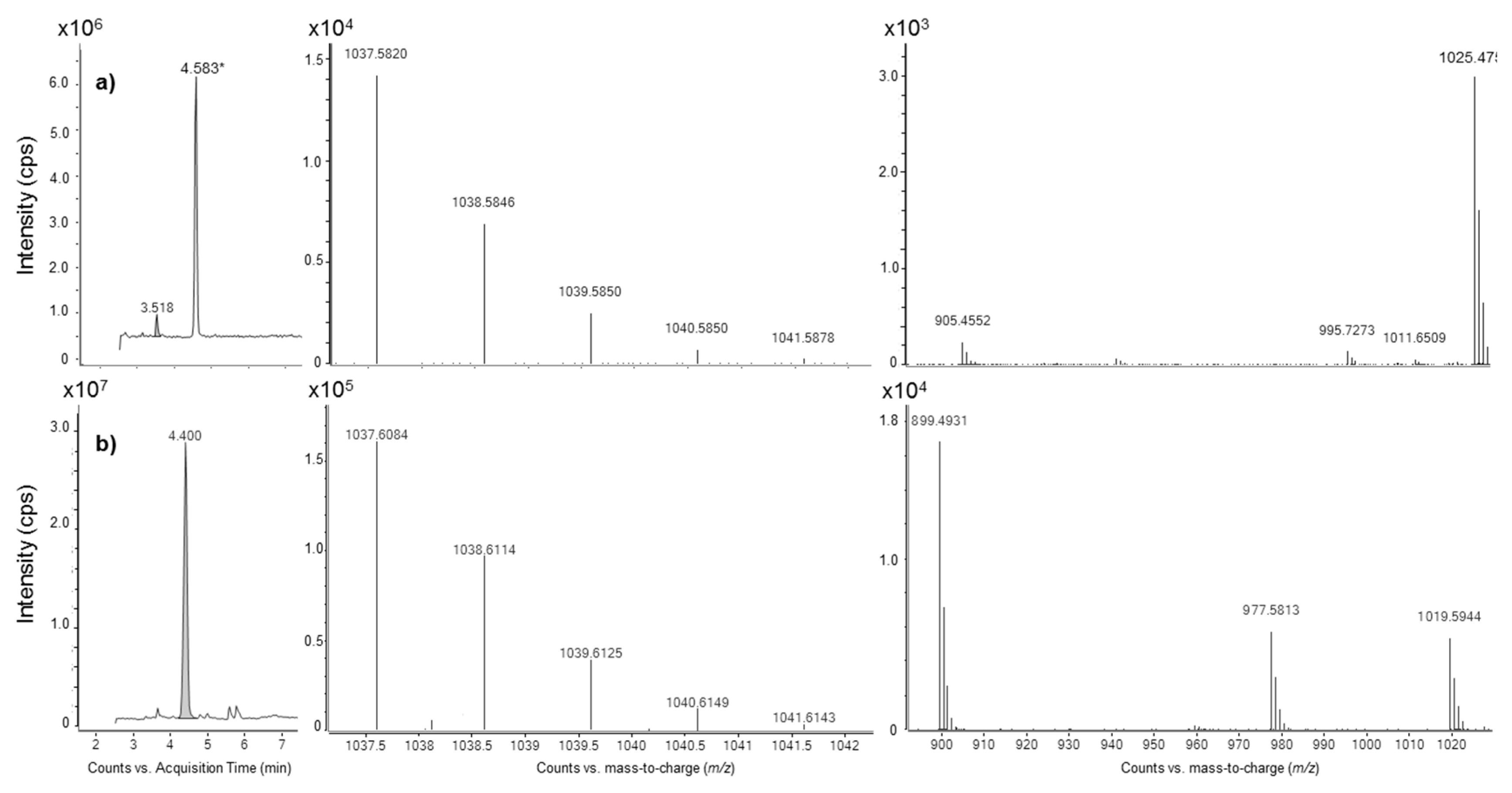
2.3. Liquid Chromatography Coupled to Full-Scan High Resolution Mass Spectrometry and Discovery of a New Maitotoxin Analog
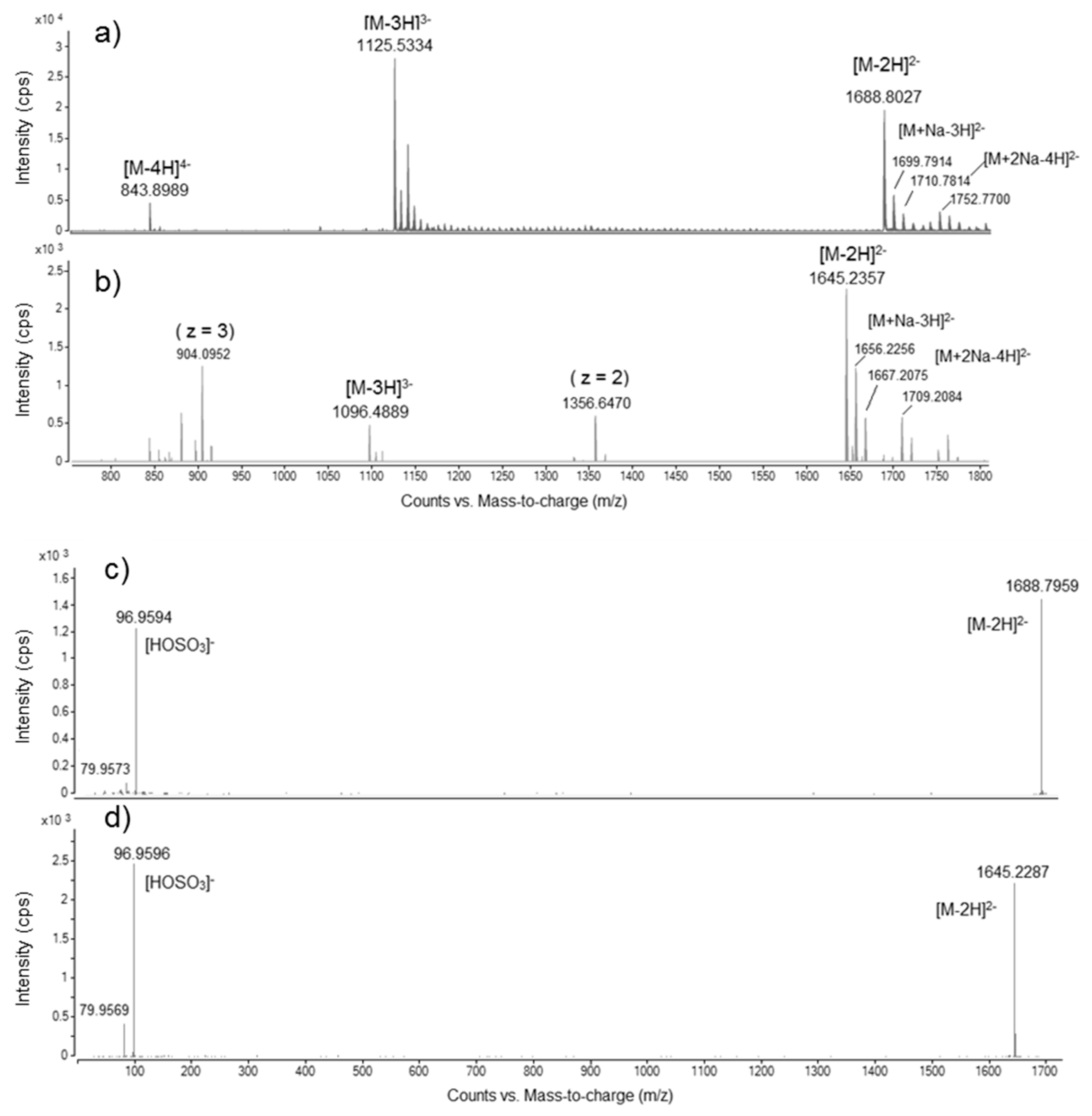
2.4. Mass Spectral Comparison of Maitotoxin-4 with Maitotoxin
2.4.1. Negative Mode HRMS Spectra
2.4.2. Negative Mode HRMS/MS Spectra
2.5. Quantification of MTX4 via LC-LRMS/MS
2.6. Relationship between N2a Cytotoxicity and MTX4 Content
2.7. Diversity of MTX Analogs among Gambierdiscus and Fukuyoa Strains
3. Discussion
3.1. Gambierdiscus excentricus and the Discovery of Maitotoxin-4
3.2. Screening for the Presence of Other MTX Analogs
4. Materials and Methods
4.1. Reference Toxins and Chemicals
4.2. Gambierdiscus and Fukuyoa Strains Examined in This Study
4.3. Cell Pellet Extraction
4.4. Fractionation of Gambierdiscus excentricus VGO791 and VGO792
4.4.1. Liquid-Liquid Partitioning
4.4.2. Size-Exclusion Chromatography (SEC): Sephadex™ LH-20
4.5. Neuroblastoma Neuro-2a (N2a) Assays
4.5.1. N2a Cytotoxicity Assay
4.5.2. N2a Calcium Flux Assay
4.6. LC-MS Analyses
4.6.1. LC-HRMS and HRMS/MS (Q-Tof 6550 iFunnel)
4.6.2. LC-LRMS/MS (API 4000 QTrap)
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Kelley, B.A.; Jollow, D.J.; Felton, E.T.; Voegtline, M.S.; Higerd, T.B. Response of mice to Gambierdiscus toxicus toxin. Mar. Fish. Rev. 1986, 48, 35–37. [Google Scholar]
- Kohli, G.S.; Papiol, G.G.; Rhodes, L.L.; Harwood, D.T.; Selwood, A.; Jerrett, A.; Murray, S.A.; Neilan, B.A. A feeding study to probe the uptake of maitotoxin by snapper (Pagrus auratus). Harmful Algae 2014, 37, 125–132. [Google Scholar] [CrossRef]
- Yasumoto, T.; Bagnis, R.; Vernoux, J. Toxicity of the surgeonfishes II. Properties of the principal water-soluble toxin. Bull. Jpn. Soc. Sci. Fish 1976, 42, 359–365. [Google Scholar] [CrossRef]
- Yasumoto, T.; Hashimoto, Y.; Bagnis, R.; Randall, J.E.; Banner, A.H. Toxicity of the surgeonfishes. Nippon Suisan Gakkaishi 1971, 37, 724–734. [Google Scholar] [CrossRef]
- Yasumoto, T.; Seino, N.; Murakami, Y.; Murata, M. Toxins produced by benthic dinoflagellates. Biol. Bull. 1987, 172, 128–131. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Matsunaga, S.; Satake, M.; Yasumoto, T. Structure and partial stereochemical assignments for maitotoxin, the most toxic and largest natural non-biopolymer. J. Am. Chem. Soc. 1994, 116, 7098–7107. [Google Scholar] [CrossRef]
- Murata, M.; Yasumoto, T. The structure elucidation and biological activities of high molecular weight algal toxins: Maitotoxin, prymnesins and zooxanthellatoxins. Nat. Prod. Rep. 2000, 17, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Murata, M.; Oshima, Y.; Iwashita, T.; Yasumoto, T. Some chemical properties of maitotoxin, a putative calcium channel agonist isolated from a marine dinoflagellate. J. Biochem. 1988, 104, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Gusovsky, F.; Daly, J.W. Maitotoxin: A unique pharmacological tool for research on calcium-dependent mechanisms. Biochem. Pharmacol. 1990, 39, 1633–1639. [Google Scholar] [CrossRef]
- Reyes, J.G.; Sánchez-Cárdenas, C.; Acevedo-Castillo, W.; Leyton, P.; López-González, I.; Felix, R.; Gandini, M.A.; Treviño, M.B.; Treviño, C.L. Maitotoxin: An enigmatic toxic molecule with useful applications in the biomedical sciences. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Botana, L.M., Ed.; CRC Press (Taylor & Francis Group): Boca Raton, FL, USA, 2014; pp. 677–694. [Google Scholar]
- Ogura, A.; Ohizumi, Y.; Yasumoto, T. Calcium-dependent depolarization induced by a marine toxin, maitotoxin, in a neuronal cell. Jpn. J. Pharmacol. 1984, 36, P315. [Google Scholar]
- Gusovsky, F.; Bitran, J.A.; Yasumoto, T.; Daly, J.W. Mechanism of maitotoxin-stimulated phosphoinositide breakdown in HL-60 cells. J. Pharmacol. Exp. Ther. 1990, 252, 466–473. [Google Scholar] [PubMed]
- Ohizumi, Y.; Yasumoto, T. Contraction and increase in tissue calcium content induced by maitotoxin, the most potent known marine toxin, in intestinal smooth muscle. Br. J. Pharmacol. 1983, 79, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Ohizumi, Y.; Yasumoto, T. Contractile response of the rabbit aorta to maitotoxin, the most potent marine toxin. J. Physiol. Lond. 1983, 337, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Ohizumi, Y.; Kajiwara, A.; Yasumoto, T. Excitatory effect of the most potent marine toxin, maitotoxin, on the guinea-pig vas deferens. J. Pharmacol. Exp. Ther. 1983, 227, 199–204. [Google Scholar] [PubMed]
- Treviño, C.L.; De la Vega-Beltrán, J.L.; Nishigaki, T.; Felix, R.; Darszon, A. Maitotoxin potently promotes Ca2+ influx in mouse spermatogenic cells and sperm, and induces the acrosome reaction. J. Cell. Physiol. 2006, 206, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, I.; Matsui, T.; Yasumoto, T.; Oshio, S.; Hoshi, M. Maitotoxin, a presumed calcium-channel activator, induces the acrosome reaction in mussel spermatozoa. Dev. Growth Differ. 1986, 28, 443–448. [Google Scholar] [CrossRef]
- Amano, T.; Okita, Y.; Yasumoto, T.; Hoshi, M. Maitotoxin induces acrosome reaction and histone degradation of starfish Asterina pectinifera sperm. Zool. Sci. 1993, 10, 307–312. [Google Scholar]
- Taglialatela, M.; Amoroso, S.; Yasumoto, T.; Di Renzo, G.; Annunziato, L. Maitotoxin and Bay-K-8644: Two putative calcium channel activators with different effects on endogenous dopamine release from tuberoinfundibular neurons. Brain Res. 1986, 381, 356–358. [Google Scholar] [CrossRef]
- Ohizumi, Y.; Kajiwara, A.; Yasumoto, T. Mode of the noradrenaline release induced by maitotoxin, the principal toxin of poisonous fishes inhabiting tropical seas, from adrenergic nerves. Folia Pharmacol. Jpn. 1983, 82, 158. [Google Scholar]
- Takahashi, M.; Tatsumi, M.; Ohizumi, Y.; Yasumoto, T. Ca2+ channel activating function of maitotoxin, the most potent marine toxin known, in clonal rat pheochromocytoma cells. J. Biol. Chem. 1983, 258, 944–949. [Google Scholar]
- Pin, J.-P.; Yasumoto, T.; Bockaert, J. Maitotoxin-evoked ɣ-aminobutyric acid release is due not only to the opening of calcium channels. J. Neurochem. 1988, 50, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Leech, C.A.; Habener, J.F. Insulinotropic glucagon-like peptide-1-mediated activation of non-selective cation currents in insulinoma cells is mimicked by maitotoxin. J. Biol. Chem. 1997, 272, 17987–17993. [Google Scholar] [CrossRef] [PubMed]
- Soergel, D.G.; Gusovsky, F.; Yasumoto, T.; Daly, J.W. Stimulatory effects of maitotoxin on insulin release in insulinoma HIT cells: Role of calcium uptake and phosphoinositide breakdown. J. Pharmacol. Exp. Ther. 1990, 255, 1360–1365. [Google Scholar] [PubMed]
- Choi, O.H.; Padgett, W.L.; Nishizawa, Y.; Gusovsky, F.; Yasumoto, T.; Daly, J.W. Maitotoxin: Effects on calcium channels, phosphoinositide breakdown, and arachidonate release in pheochromocytoma PC12 cells. Mol. Pharmacol. 1990, 37, 222–230. [Google Scholar] [PubMed]
- Columbo, M.; Taglialatela, M.; Warner, J.A.; Macglashan, D.W.; Yasumoto, T.; Annunziato, L.; Marone, G. Maitotoxin, a novel activator of mediator release from human basophils, induces large increase in cytosolic calcium resulting in histamine, but not leukotriene-Crelease. J. Pharmacol. Exp. Ther. 1992, 263, 979–986. [Google Scholar] [PubMed]
- Estacion, M.; Schilling, W.P. Maitotoxin-induced membrane blebbing and cell death in aortic endothelial cells. FASEB J. 2001, 1, 2. [Google Scholar]
- Schilling, W.P.; Sinkins, W.G.; Estacion, M. Maitotoxin activates a nonselective cation channel and a P2Z/P2X(7)-like cytolytic pore in human skin fibroblasts. Am. J. Physiol. Cell Physiol. 1999, 277, C755–C765. [Google Scholar]
- Schilling, W.P.; Wasylyna, T.; Dubyak, G.R.; Humphreys, B.D.; Sinkins, W.G. Maitotoxin and P2Z/P2X(7) purinergic receptor stimulation activate a common cytolytic pore. Am. J. Physiol. Cell Physiol. 1999, 277, C766–C776. [Google Scholar]
- Sasaki, M.; Matsumori, N.; Maruyama, T.; Nonomura, T.; Murata, M.; Tachibana, K.; Yasumoto, T. The complete structure of maitotoxin, part I: Configuration of the C1-C14 side chain. Angew. Chem. Int. Ed. Engl. 1996, 35, 1672–1675. [Google Scholar] [CrossRef]
- Nonomura, T.; Sasaki, M.; Matsumori, N.; Murata, M.; Tachibana, K.; Yasumoto, T. The complete structure of maitotoxin, part II: Configuration of the C135-C142 side chain and absolute configuration of the entire molecule. Angew. Chem. Int. Ed. Engl. 1996, 35, 1675–1678. [Google Scholar] [CrossRef]
- Zheng, W.J.; DeMattei, J.A.; Wu, J.P.; Duan, J.J.W.; Cook, L.R.; Oinuma, H.; Kishi, Y. Complete relative stereochemistry of maitotoxin. J. Am. Chem. Soc. 1996, 118, 7946–7968. [Google Scholar] [CrossRef]
- Gallimore, A.R.; Spencer, J.B. Stereochemical uniformity in marine polyether ladders—Implications for the biosynthesis and structure of maitotoxin. Angew. Chem. Int. Ed. Engl. 2006, 45, 4406–4413. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Frederick, M.O. On the structure of maitotoxin. Angew. Chem. Int. Ed. 2007, 46, 5278–5282. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Cole, K.P.; Frederick, M.O.; Aversa, R.J.; Denton, R.M. Chemical synthesis of the GHIJK ring system and further experimental support for the originally assigned structure of maitotoxin. Angew. Chem. Int. Ed. 2007, 46, 8875–8879. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Ohizumi, Y.; Yasumoto, T. Maitotoxin, a Ca2+ channel activator candidate. J. Biolog. Chem. 1982, 257, 7287–7289. [Google Scholar]
- Freedman, S.B.; Miller, R.J.; Miller, D.M.; Tindall, D.R. Interactions of maitotoxin with voltage-sensitive calcium channels in cultured neuronal cells. Proc. Natl. Acad. Sci. USA 1984, 81, 4582–4585. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Secondo, A.; D’Alessio, A.; Taglialatela, M.; Hofmann, F.; Klugbauer, N.; Di Renzo, G.; Annunziato, L. Studies on maitotoxin-induced intracellular Ca2+ elevation in Chinese hamster ovary cells stably transfected with cDNAs encoding for L-type Ca2+ channel subunits. J. Pharmacol. Exp. Ther. 1999, 290, 725–730. [Google Scholar] [PubMed]
- Brereton, H.M.; Chen, J.L.; Rychkov, G.; Harland, M.L.; Barritt, G.J. Maitotoxin activates an endogenous non-selective cation channel and is an effective initiator of the activation of the heterologously expressed hTRPC-1 (transient receptor potential) non-selective cation channel in H4-IIE liver cells. Biochim. Biophys. Acta Mol. Cell Res. 2001, 1540, 107–126. [Google Scholar] [CrossRef]
- Morales-Tlalpan, V.; Vaca, L. Modulation of the maitotoxin response by intracellular and extracellular cations. Toxicon 2002, 40, 493–500. [Google Scholar] [CrossRef]
- Gutierrez, D.; Diaz de Leon, L.; Vaca, L. Characterization of the maitotoxin-induced calcium influx pathway from human skin fibroblasts. Cell Calcium 1997, 22, 31–38. [Google Scholar] [CrossRef]
- Martinez-Francois, J.R.; Morales-Tlalpan, V.; Vaca, L. Characterization of the maitotoxin-activated cationic current from human skin fibroblasts. J. Physiol. Lond. 2002, 538, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.A.; Mattei, C.; Molgo, J. Marine toxins potently affecting neurotransmitter release. Prog. Mol. Subcell. Biol. 2009, 46, 159–186. [Google Scholar] [PubMed]
- Frew, R.; Wang, Y.S.; Weiss, T.M.; Nelson, P.; Sawyer, T.W. Attenuation of maitotoxin-induced cytotoxicity in rat aortic smooth muscle cells by inhibitors of Na+/Ca2+ exchange, and calpain activation. Toxicon 2008, 51, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Weiss, M.T.; Yin, J.F.; Frew, R.; Tenn, C.; Nelson, P.P.; Vair, C.; Sawyer, T.W. Role of the sodium hydrogen exchanger in maitotoxin-induced cell death in cultured rat cortical neurons. Toxicon 2009, 54, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Vale, C.; Antelo, A.; Hirama, M.; Yamashita, S.; Vieytes, M.R.; Botana, L.M. Differential effects of ciguatoxin and maitotoxin in primary cultures of cortical neurons. Chem. Res. Toxicol. 2014, 27, 1387–1400. [Google Scholar] [CrossRef] [PubMed]
- Sinkins, W.G.; Estacion, M.; Prasad, V.; Goel, M.; Shull, G.E.; Kunze, D.L.; Schilling, W.P. Maitotoxin converts the plasmalemmal Ca2+ pump into a Ca2+-permeable nonselective cation channel. Am. J. Physiol. Cell Physiol. 2009, 297, C1533–C1543. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, L.A.; Alvarez-Parrilla, E.; Martinez-Martinez, A. Biochemistry of maitotoxin. In Phycotoxins: Chemistry and Biochemistry; Botana, L.M., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 55–74. [Google Scholar]
- Lundy, P.M.; Nelson, P.; Mi, L.; Frew, R.; Minaker, S.; Vair, C.; Sawyer, T.W. Pharmacological differentiation of the P2X(7) receptor and the maitotoxin-activated cationic channel. Eur. J. Pharmacol. 2004, 487, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Konoki, K.; Hashimoto, M.; Murata, M.; Tachibana, K. Maitotoxin-induced calcium influx in erythrocyte ghosts and rat glioma C6 cells, and blockade by gangliosides and other membrane lipids. Chem. Res. Toxicol. 1999, 12, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Gusovsky, F.; Sasaki, M.; Yokoyama, A.; Yasumoto, T.; Daly, J.W. Effect of maitotoxin analogs on calcium influx and phosphoinositide breakdown in cultured cells. Toxicon 1991, 29, 1085–1096. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Purification and characterisation of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Matsumori, N.; Konoki, K.; Oishi, T. Structural features of dinoflagellate toxins underlying biological activity as viewed by NMR. Bull. Chem. Soc. Jpn. 2008, 81, 307–319. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Gillespie, N.C. Toxicity of Australian and French Polynesian strains of Gambierdiscus toxicus (Dinophyceae) grown in culture: Characterization of a new type of maitotoxin. Toxicon 1990, 28, 1159–1172. [Google Scholar] [CrossRef]
- Lewis, R.J.; Holmes, M.J.; Alewood, P.F.; Jones, A. Ionspray mass spectrometry of ciguatoxin-1, maitotoxin-2 and -3, and related marine polyether toxins. Nat. Toxins 1994, 2, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Inserra, M.; Vetter, I.; Holland, W.C.; Hardison, D.R.; Tester, P.A.; Litaker, R.W. Rapid extraction and identification of maitotoxin and ciguatoxin-like toxins from Caribbean and Pacific Gambierdiscus using a new functional bioassay. PLoS ONE 2016, 11, e0160006. [Google Scholar] [CrossRef] [PubMed]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Chinain, M.; Holmes, M.J.; Holland, W.C.; Tester, P.A. Taxonomy of Gambierdiscus including four new species, Gambierdiscus caribaeus, Gambierdiscus carolinianus, Gambierdiscus carpenteri and Gambierdiscus ruetzleri (Gonyaulacales, Dinophyceae). Phycologia 2009, 48, 344–390. [Google Scholar] [CrossRef]
- Smith, K.F.; Rhodes, L.; Verma, A.; Curley, B.G.; Harwood, D.T.; Kohli, G.S.; Solomona, D.; Rongo, T.; Munday, R.; Murray, S.A. A new Gambierdiscus species (Dinophyceae) from Rarotonga, Cook Islands: Gambierdiscus cheloniae sp. nov. Harmful Algae 2016, 60, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodríguez, F.; Riobó, P.; Bravo, I. Gambierdiscus balechii sp. nov. (Dinophyceae), a new benthic toxic dinoflagellate from the Celebes Sea (SW Pacific Ocean). Harmful Algae 2016, 58, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef]
- Fraga, S.; Rodríguez, F. Genus Gambierdiscus in the Canary Islands (NE Atlantic Ocean) with description of Gambierdiscus silvae sp. nov., a new potentially toxic epiphytic benthic dinoflagellate. Protist 2014, 165, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Yamaguchi, H.; Adachi, M. Morphology of Gambierdiscus scabrosus sp. nov. (Gonyaulacales): A new epiphytic toxic dinoflagellate from coastal areas of Japan. J. Phycol. 2014, 50, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, A.L.; Verma, A.; Harwood, D.T.; Hoppenrath, M.; Murray, S.A. Characterisation of Gambierdiscus lapillus sp. nov. (Gonyaulacales, Dinophyceae): A new toxic dinoflagellate from the Great Barrier Reef (Australia). J. Phycol. 2017, 53, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F.; Qiu, D.J.; Lopes, R.M.; Lin, S.J. Fukuyoa paulensis gen. et sp. nov., a new genus for the globular species of the dinoflagellate Gambierdiscus (Dinophyceae). PLoS ONE 2015, 10, e0119676. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; Smith, K.F.; Verma, A.; Curley, B.G.; Harwood, D.T.; Murray, S.; Kohli, G.S.; Solomona, D.; Rongo, T.; Munday, R.; et al. A new species of Gambierdiscus (Dinophyceae) from the south-west Pacific: Gambierdiscus honu sp. nov. Harmful Algae 2017, 65, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Tunin-Ley, A.; Chomérat, N.; Bilien, G.; Turquet, J. Taxonomic characterization of Gambierdiscus species in Reunion Island, Indian Ocean. In Proceedings of the GdR Phycotox & GIS Cyano 2017, Gif-sur-Yvette, France, 14–16 March 2017. [Google Scholar]
- Holland, W.C.; Litaker, R.W.; Tomas, C.R.; Kibler, S.R.; Place, A.R.; Davenport, E.D.; Tester, P.A. Differences in the toxicity of six Gambierdiscus (Dinophyceae) species measured using an in vitro human erythrocyte lysis assay. Toxicon 2013, 65, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; Harwood, T.; Smith, K.; Argyle, P.; Munday, R. Production of ciguatoxin and maitotoxin by strains of Gambierdiscus australes, G. pacificus and G. polynesiensis (Dinophyceae) isolated from Rarotonga, Cook Islands (vol 39, pg 185, 2014). Harmful Algae 2016, 55, 295. [Google Scholar] [CrossRef] [PubMed]
- Kohli, G.S. Diversity and Genetics of Australasian Dinoflagellates, Including Gambierdiscus spp. the Causative Agent of Ciguatera Fish Poisoning; University of New South Wales: Sidney, Australia, 2013. [Google Scholar]
- Rhodes, L.; Papiol, G.G.; Smith, K.; Harwood, T. Gambierdiscus cf. yasumotoi (Dinophyceae) isolated from New Zealand’s sub-tropical northern coastal waters. N. Z. J. Mar. Freshw. Res. 2014, 48, 303–310. [Google Scholar]
- Rhodes, L.L.; Smith, K.F.; Verma, A.; Murray, S.; Harwood, D.T.; Trnski, T. The dinoflagellate genera Gambierdiscus and Ostreopsis from subtropical Raoul Island and North Meyer Island, Kermadec Islands. N. Z. J. Mar. Freshw. Res. 2017, 1–15. [Google Scholar] [CrossRef]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Tester, P. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, unpublished. [Google Scholar]
- Boada, L.D.; Zumbado, M.; Luzardo, O.P.; Almeida-González, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.; Dickey, R.W. Ciguatera fish poisoning on the West Africa Coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Arellano, J.L.; Luzardo, O.P.; Perez Brito, A.; Hernandez Cabrera, M.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, S.M.; Diniz, B.; de Alencar, A.G.; Menegueli, A.C.; Menezes, M. First record of the ciguatera causing genus Gambierdiscus in Brazil. Harmful Algal News 2012, 45, 8–9. [Google Scholar]
- Nascimento, S.M.; Melo, G.; Salgueiro, F.; Diniz, B.D.; Fraga, S. Morphology of Gambierdiscus excentricus (Dinophyceae) with emphasis on sulcal plates. Phycologia 2015, 54, 628–639. [Google Scholar] [CrossRef]
- Caillaud, A.; de la Iglesia, P.; Barber, E.; Eixarch, H.; Mohammad-Noor, N.; Yasumoto, T.; Diogène, J. Monitoring of dissolved ciguatoxin and maitotoxin using solid-phase adsorption toxin tracking devices: Application to Gambierdiscus pacificus in culture. Harmful Algae 2011, 10, 433–446. [Google Scholar] [CrossRef]
- Kibler, S.R.; Litaker, R.W.; Holland, W.C.; Vandersea, M.W.; Tester, P.A. Growth of eight Gambierdiscus (Dinophyceae) species: Effects of temperature, salinity and irradiance. Harmful Algae 2012, 19, 1–14. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Liefer, J.D.; Robertson, A.; Kulis, D.; Smith, T.B.; Parsons, M.L.; Anderson, D.M. Influence of environmental variables on Gambierdiscus spp. (Dinophyceae) growth and distribution. PLoS ONE 2016, 11, e0153197. [Google Scholar] [CrossRef] [PubMed]
- Roeder, K.; Erler, K.; Kibler, S.; Tester, P.; Van The, H.; Nguyen-Ngoc, L.; Gerdts, G.; Luckas, B. Characteristic profiles of ciguatera toxins in different strains of Gambierdiscus spp. Toxicon 2010, 56, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a ladder-shaped polyether from the dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Uchida, H.; Suzuki, T.; Matsushima, R.; Nagae, M.; Toyohara, Y.; Satake, M.; Oshima, Y.; Inoue, A.; Yasumoto, T. Gambieroxide, a novel epoxy polyether compound from the dinoflagellate Gambierdiscus toxicus GTP2 strain. Tetrahedron 2013, 69, 10299–10303. [Google Scholar] [CrossRef]
- Babinchak, J.A.; Jollow, D.J.; Voegtline, M.S.; Higerd, T.B. Toxin production by Gambierdiscus toxicus isolated from the Florida Keys. Mar. Fish. Rev. 1986, 48, 53–56. [Google Scholar]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Uehara, K.; Shah, M.M.; Suda, S.; Yasumoto, T.; Taira, Y.; Yamaguchi, H.; et al. Genetic diversity and distribution of the ciguatera-causing dinoflagellate Gambierdiscus spp. (Dinophyceae) in coastal areas of Japan. PLoS ONE 2013, 8, e60882. [Google Scholar] [CrossRef] [PubMed]
- Bravo, I.; Figueroa, R.I.; Fraga, S. Cellular and nuclear morphological variability within a single species of the toxigenic dinoflagellate genus Gambierdiscus: Relationship to life-cycle processes. Harmful Algae 2014, 40, 1–8. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef] [PubMed]
- The, H.V.; Lam, N.N.; Larsen, J. On a toxic benthic dinoflagellate Gambierdiscus toxicus isolated from Cau Island, South Central Viet Nam. In Proceedings of the 13th International Conference on Harmful Algae, Hong Kong, China, 3–7 November 2008. [Google Scholar]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Caillaud, A.; Yasumoto, T.; Diogène, J. Detection and quantification of maitotoxin-like compounds using a neuroblastoma (Neuro-2a) cell based assay. Application to the screening of maitotoxin-like compounds in Gambierdiscus spp. Toxicon 2010, 56, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.; et al. Fluorescent receptor binding assay for detecting ciguatoxins in fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- O’Brien, P.J.; Irwin, W.; Diaz, D.; Howard-Cofield, E.; Krejsa, C.M.; Slaughter, M.R.; Gao, B.; Kaludercic, N.; Angeline, A.; Bernardi, P.; et al. High concordance of drug-induced human hepatotoxicity with in vitro cytotoxicity measured in a novel cell-based model using high content screening. Arch. Toxicol. 2006, 80, 580–604. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Carpenter, A.E.; Genovesio, A. Increasing the content of High-Content Screening. J. Biomol. Screen. 2014, 19, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Sérandour, A.L.; Ledreux, A.; Morin, B.; Derick, S.; Augier, E.; Lanceleur, R.; Hamlaoui, S.; Moukha, S.; Furger, C.; Biré, R.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part I: Screening strategy and pre-validation study with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Vial, J.; Jardy, A. Experimental comparison of the different approaches to estimate LOD and LOQ of an HPLC method. Anal. Chem. 1999, 71, 2672–2677. [Google Scholar] [CrossRef]
- Sanagi, M.M.; Ling, S.L.; Nasir, Z.; Hermawan, D.; Ibrahim, W.A.; Abu Naim, A. Comparison of signal-to-noise, blank determination, and linear regression methods for the estimation of detection and quantification limits for volatile organic compounds by gas chromatography. J. AOAC Int. 2009, 92, 1833–1838. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbr. | Formula | Mass (Da) | Structural Studies | Toxicity (i.p. LD50 in Mice, µg kg−1) | Source | Reference |
|---|---|---|---|---|---|---|---|
| Maitotoxin | MTX | C164H256O68S2Na2 | 3423.5811 | IR, UV (λmax = 230 nm) LRMS/MS: FAB− NMR with complete stereochemistry | 0.050 | GII-1 | [1,31,32,33] |
| Maitotoxin-2 | MTX2 | UNKN (mono-sodiated salt of a molecule containing one sulfate ester) | 3298 | UV (λmax = 230 nm) LRMS: IS+, IS−, FAB− | 0.080 | NQ1 | [55] |
| Maitotoxin-3 | MTX3 | UNKN (di-sodiated salt of a molecule containing two sulfate esters) | 1060.5 | UV (λmax = 200, 235 nm) LRMS: IS+ | UNKN | WC1/1 | [53] |
| MTX | MTX4 | ||
|---|---|---|---|
| Elemental formula (free acid form) | C164H258O68S2 | UNKN | |
| Retention time (RT, min) | 4.09 min | 4.58 min | |
| Ion species (accurate mono-isotopic m/z) | [M − 2H]2− | 1688.8027 (Δppm: −0.8) | 1645.2357 |
| [M + Na − 3H]2− | 1699.7914 (Δppm: +0.5) | 1656.2256 | |
| [M + 2Na − 4H]2− | 1710.7814 (Δppm: +1.1) | 1667.2075 | |
| [M − 3H]3− | 1125.5334 (Δppm: −1.4) | 1096.4889 | |
| [M − 4H]4− | 843.8989 (Δppm: −2.1) | ND | |
| Strain Name | Sample Name | MTX4 (µg MTX eq) | % Recovery (Liquid-Liquid Partitioning) | % Recovery (LH-20) |
|---|---|---|---|---|
| Gambierdiscus excentricus VGO791 | Crude extract | 160.1 | 100% | |
| Liquid-liquid partitioning of crude extract | ||||
| MSF * | 137.4 | 85.8% | ||
| Size-exclusion chromatography (LH-20) of MSF sample | ||||
| Ve = 12.5–15.0 mL | 19.1 | 13.9% | ||
| Ve = 15.0–17.5 mL | 69.2 | 50.4% | ||
| Ve = 17.5–20.0 mL | 19.0 | 13.8% | ||
| Ve = 20.0–22.5 mL | 4.5 | 3.3% | ||
| Ve = 22.5–25.0 mL | 1.5 | 1.1% | ||
| Ve = 25.0–27.5 mL | 1.2 | 0.9% | ||
| Gambierdiscus excentricus VGO792 | Crude extract | 43.1 | 100% | |
| Liquid-liquid partitioning of crude extract | ||||
| MSF * | 34.2 | 79.4% | ||
| Size-exclusion chromatography (LH-20) of MSF sample | ||||
| Ve = 13.0–14.0 mL | 0.6 | 1.7% | ||
| Ve = 14.0–15.0 mL | 4.6 | 13.5% | ||
| Ve = 15.0–16.0 mL | 10.3 | 30.1% | ||
| Ve = 16.0–17.0 mL | 6.2 | 18.0% | ||
| Ve = 17.0–18.0 mL | 3.3 | 9.6% | ||
| Ve = 18.0–19.0 mL | 1.8 | 5.3% | ||
| Ve = 19.0–20.0 mL | 1.2 | 3.4% | ||
| Ve = 20.0–21.0 mL | 0.8 | 2.5% | ||
| Ve = 21.0–22.0 mL | 0.6 | 1.9% | ||
| Ve = 22.0–23.0 mL | 0.6 | 1.7% | ||
| Ve = 23.0–24.0 mL | <LOQ | |||
| Ve = 24.0–25.0 mL | <LOQ | |||
| Ve = 25.0–26.0 mL | <LOQ | |||
| Species | Strain | MTX | p-MTX2 | p-MTX3 | MTX4 | ||
|---|---|---|---|---|---|---|---|
| pg MTX cell−1 ± SD (n = 3) RT = 4.64 min | 1091.5 → 96.9 | 1103.8 → 96.9 | 1015.5 → 96.9 | 1037.6 → 96.9 | pg MTX eq cell−1 ± SD (n = 3) RT = 5.05 min | ||
| F. ruetzleri | Gam1 | ND (<2.97) | ND | ND | ND | RT = 4.89 & 5.74 | ND (<2.97) |
| WH55-Gam4 | ND (<4.97) | ND | ND | ND | RT = 4.88 | ND (<4.97) | |
| G. australes | CCMP1653 (NOAA 24) (T39) | ND (<1.54) | ND | ND | ND | RT = 4.86 | ND (<1.54) |
| S080911_1 | 22.6 ± 0.5 | ND | ND | ND | RT = 4.86 | ND (<0.80) | |
| VGO1178 | ND (<2.46) | ND | ND | ND | RT = 4.87 | ND (<2.46) | |
| VGO1181 | ND (<2.22) | ND | ND | ND | RT = 4.87 | ND (<2.22) | |
| G. balechii | VGO917 | ND (<1.66) | ND | ND | RT = 4.10 & 4.21 | RT = 4.80 | ND (<1.66) |
| VGO920 | ND (<2.02) | ND | ND | RT = 3.94 | RT = 4.86 | ND (<2.02) | |
| G. belizeanus | CCMP399 (NOAA2) (SB03) | ND (<3.51) | ND | ND | ND | RT = 4.80 | ND (<3.51) |
| Keys Gam1 | ND (<7.79) | ND | ND | ND | RT = 4.80 | ND (<7.79) | |
| ST1-F4 | ND (<6.20) | ND | ND | ND | RT = 4.83 | ND (<6.20) | |
| G. caribaeus | CCMP1733 (NOAA11) | ND (<2.27) | RT = 3.64 & 4.28 | RT = 3.64 & 4.28 | ND | RT = 4.84 | ND (<2.27) |
| Bill Hi Gam8 | ND (<5.79) | RT = 4.05 & 4.30 | RT = 4.05 & 4.30 | ND | RT = 4.83 | ND (<5.79) | |
| CCMP1651 (NOAA20) | ND (<10.27) | RT = 4.27 | RT = 4.27 | ND | RT = 4.82 | ND (<10.27) | |
| Dive 1 fa Gam1 | ND (<1.69) | RT = 4.26 | RT = 4.26 | ND | RT = 4.83 | ND (<1.69) | |
| Mexico Algae1 Gam1 | ND (<5.19) | RT = 4.27 | RT = 4.27 | ND | RT = 4.80 | ND (<5.19) | |
| G. carolinianus | ETB Exp28 Gam10 | ND (<10.49) | ND | ND | RT = 4.55, 6.11 & 6.21 | RT = 4.89, 6.11 & 6.21 | ND (<10.49) |
| Greece Gam2 | ND (<7.73) | ND | ND | RT = 4.54 & 6.21 | RT = 4.86, 6.21 & 7.17 | ND (<7.73) | |
| RROV5 | ND (<5.47) | ND | ND | RT = 5.12 & 6.25 | RT = 5.12 & 6.25 | ND (<5.47) | |
| G. carpenteri | GT4 | ND (<7.27) | ND | ND | ND | RT = 4.86 | ND (<7.27) |
| Jamaica Algae2 Gam1 | ND (<6.51) | ND | ND | ND | RT = 4.81 | ND (<6.51) | |
| Pat Hi Jar7 Gam11 | ND (<10.72) | ND | ND | ND | RT = 4.86 | ND (<10.72) | |
| WHBR21 | ND (<6.62) | ND | ND | ND | RT = 4.84 | ND (<6.62) | |
| G. excentricus | Pulley Ridge Gam 2 | ND (<0.39) | RT = 5.68 | ND | ND | RT = 4.01 | 22.9 * |
| UNR-07 | ND (<0.63) | RT = 5.64 | ND | ND | RT = 4.05 | 16.0 ± 2.3 | |
| UNR-08 | ND (<1.45) | RT = 5.68 | ND | ND | RT = 4.05 | 19.8 ± 6.4 | |
| VGO1035 | ND (<2.64) | RT = 5.68 | ND | ND | RT = 4.01 | 13.0 * | |
| VGO790 | ND (<3.73) | RT = 5.67 | ND | ND | RT = 4.01 | 23.2 * | |
| VGO791 | ND (<0.29) | RT = 5.67 | ND | ND | RT = 3.71 & 4.01 | 72.8 ± 8.5 | |
| VGO792 | ND (<0.30) | RT = 5.68 | ND | ND | RT = 3.68 & 3.99 | 20.0 ± 2.9 | |
| G. pacificus | CCMP1650 (NOAA 9) (MR1) | ND (<1.02) | RT = 5.33, 6.06, 6.24 & 6.82 | RT = 5.33, 6.06, 6.24 & 6.82 | ND | RT = 4.89 | ND (<1.02) |
| G10DC | ND (<0.66) | RT = 5.06 | ND | RT = 3.90 & 4.15 | - | ND (<0.66) | |
| Gambierdiscus sp. ribotype 2 | CCMP1655 (MQ2) | ND (<5.87) | ND | ND | ND | RT = 4.83 | ND (<5.87) |
| Mixed PR | ND (<6.49) | ND | ND | ND | RT = 4.90 | ND (<6.49) | |
| St Maartens Gam10 | ND (<5.52) | ND | ND | ND | RT = 4.90 | ND (<5.52) | |
| SW Algae Gam1 | ND (<8.38) | ND | ND | ND | RT = 4.81 | ND (<8.38) | |
| G. scabrosus | KW070922_1 | ND (<0.83) | ND | ND | ND | RT = 4.80 | ND (<0.83) |
| G. silvae | UNR-30 | ND (<1.76) | ND | ND | RT = 5.93 & 6.13 | RT = 3.22, 4.76, 5.35, 5.93, 6.13, 6.98 | ND (<1.76) |
| VGO1167 | ND (<1.27) | ND | ND | - | RT = 4.83 | ND (<1.27) | |
| VGO1180 | ND (<1.52) | ND | ND | RT = 5.92 & 6.14 | RT = 4.68, 5.92 & 6.14 | ND (<1.52) | |
| Gambierdiscus sp. | Viet Nam | ND (<0.37) | RT = 5.14 | ND | RT = 7.02 | RT = 7.64 | ND (<0.37) |
| G. toxicus | GTT-91 | ND (<5.49) | ND | ND | RT = 2.94 | RT = 4.89 | ND (<5.49) |
| HIT-0 | ND (<6.00) | ND | ND | RT = 2.92 | RT = 4.89 | ND (<6.00) | |
| HIT-25 | ND (<8.56) | ND | ND | RT = 2.90 | RT = 4.88 | ND (<8.56) | |
| Species/Phylotype | Strain | Geographical Origin | Culture Collection | Culture Conditions | Number of Cells Extracted | Reference |
|---|---|---|---|---|---|---|
| F. ruetzleri | Gam1 | Southwater Cay, Belize | CCFHR | CCFHR | 215,190 | [57,68] |
| WH55-Gam4 | Flower Garden Banks National Marine Sanctuary (West Bank), Northwestern Gulf of Mexico, United States of America | CCFHR | CCFHR | 128,800 | [75] | |
| G. australes | CCMP1653 (NOAA 24) (T39) | Tern Island, Hawaii, United States of America | NCMA | PHYC | 416,220 | [58,86] |
| S080911_1 | Kutsu, Susaki, Kochi, Japan | KU | PHYC | 798,285 | [87] | |
| VGO1178 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 260,388 | [62,69] | |
| VGO1181 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 288,608 | [69] | |
| G. balechii | VGO917 | Manado, Celebes Sea, Indonesia | CCVIEO | PHYC | 384,650 | [60,88] |
| VGO920 | Manado, Celebes Sea, Indonesia | CCVIEO | PHYC | 317,600 | [60] | |
| G. belizeanus | CCMP399 (NOAA2) (SB03) | St. Barthélemy Island, Caribbean, Territorial collectivity of Saint-Barthélemy | NCMA | PHYC | 182,495 | [57,58] |
| Keys Gam1 | Florida Keys, Florida, United States of America | CCFHR | CCFHR | 82,125 | [68] | |
| ST1-F4 | St. Thomas, US Virgin Islands, United States of America | CCFHR | CCFHR | 103,300 | [68] | |
| G. caribaeus | CCMP1733 (NOAA11) | Carrie Bow Cay, Belize, Caribbean, United States of America | NCMA | PHYC | 282,450 | [58] |
| Bill Hi Gam8 | Waikiki Beach, Honolulu, Hawaii, United States of America | CCFHR | CCFHR | 110,600 | [69] | |
| CCMP1651 (NOAA20) | Grand Cayman Island, Caribbean, Territory of United Kingdom | NCMA | CCFHR | 62,300 | [58] | |
| Dive 1 fa Gam1 | Ft. Pierce, Florida, United States of America | CCFHR | CCFHR | 378,000 | [68] | |
| Mexico Algae1 Gam1 | Cancun, Mexico | CCFHR | CCFHR | 123,395 | [68] | |
| G. carolinianus | ETB Exp28 Gam10 | Dry Tortugas, United States of America | CCFHR | CCFHR | 61,005 | [68] |
| Greece Gam2 | Crete, Greece | CCFHR | CCFHR | 82,775 | [69] | |
| RROV5 | Puerto Rico, United States of America | CCFHR | CCFHR | 116,985 | [68] | |
| G. carpenteri | GT4 | Carrie Bow Cay, Belize | CCFHR | CCFHR | 88,000 | [57,68] |
| Jamaica Algae2 Gam1 | Ocho Rios, Jamaica | CCFHR | CCFHR | 98,250 | [68] | |
| Pat Hi Jar7 Gam11 | Waikiki Beach, Honolulu, Hawaii, United States of America | CCFHR | CCFHR | 59,680 | [57,69] | |
| WHBR21 | Flower Garden Banks National Marine Sanctuary (West Bank), Northwestern Gulf of Mexico, United States of America | CCFHR | CCFHR | 96,720 | [68] | |
| G. excentricus | Pulley Ridge Gam 2 | Pulley Ridge, Florida, United States of America | CCFHR | CCFHR | 1,630,000 | [75] |
| UNR-07 | Armação dos Búzios, Rio de Janeiro, Brazil | UNIRIO | UNIRIO | 1,013,833 | [79] | |
| UNR-08 | Armação dos Búzios, Rio de Janeiro, Brazil | UNIRIO | UNIRIO | 441,490 | [79] | |
| VGO1035 | Playa Las Cabras, La Palma, Canary Islands, Spain) | CCVIEO | PHYC | 242,050 | [61] | |
| VGO790 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 171,711 | [61] | |
| VGO791 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 2,200,160 | [61] | |
| VGO792 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 2,159,997 | [61] | |
| G. pacificus | CCMP1650 (NOAA 9) (MR1) | Moorea, Society Islands, French Polynesia | NCMA | PHYC | 630,175 | [58] |
| G10DC | Malaysia | IRTA | PHYC | 965,040 | [80] | |
| Gambierdiscus sp. ribotype 2 | CCMP1655 (MQ2) | Martinique, Caribbean, insular region of France | NCMA | PHYC | 109,080 | [89] |
| Mixed PR | Puerto Rico, United States of America | CCFHR | CCFHR | 98,560 | [75] | |
| St Maartens Gam10 | St. Maarteens, Kingdom of the Netherlands | CCFHR | CCFHR | 116,000 | [75] | |
| SW Algae Gam1 | Southwater Cay, Belize | CCFHR | CCFHR | 76,410 | [75] | |
| G. scabrosus | KW070922_1 | Kashiwa-jima Island, Otsuki, Kochi, Japan | KU | PHYC | 771,711 | [63,87] |
| G. silvae | UNR-30 | Brazil | UNIRIO | UNIRIO | 362,877 | Unpublished strain |
| VGO1167 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 502,080 | [62] | |
| VGO1180 | Punta Hidalgo, Tenerife, Canary Islands, Spain | CCVIEO | PHYC | 421,430 | [62] | |
| Gambierdiscus sp. | Viet Nam | Cau Island, Binh Thuan, South China Sea, Viet Nam | VNIO | PHYC | 1,726,949 | [67,83,90] |
| G. toxicus | GTT-91 | Teahupoo, Tahiti, French Polynesia | CCFHR | CCFHR | 116,625 | [70] |
| HIT-0 | Hitiaa, Tahiti, French Polynesia | CCFHR | CCFHR | 106,650 | [70] | |
| HIT-25 | Hitiaa, Tahiti, French Polynesia | CCFHR | CCFHR | 74,800 | [70] |
| Compound | MRM Transitions (m/z) * | CE (eV) | CXP (eV) | |
|---|---|---|---|---|
| MTX | [M − 2H]2−/[M − 2H]2− | 1689.8/1689.8 | −40 | −15 |
| [M − 2H]2−/[HOSO3]− | 1689.8/96.9 | −125 | −21 | |
| [M + Na − 3H]2−/[HOSO3]− | 1700.8/96.9 | −125 | −21 | |
| [M + 2Na − 3H]2−/[HOSO3]− | 1711.8/96.9 | −125 | −21 | |
| [M − 3H]3−/[M − 3H]3− | 1126.2/1126.2 | −40 | −15 | |
| [M − 3H]3−/[HOSO3]− | 1126.2/96.9 | −125 | −21 | |
| [M − 4H]4−/[HOSO3]− | 844.4/96.9 | −125 | −21 | |
| MTX2 | [M − 2H]2−/[HOSO3]− | 1637.5/96.9 | −125 | −21 |
| [M + Na − 3H]2−/[HOSO3]− | 1648.2/96.9 | −125 | −21 | |
| [M + K − 3H]2−/[HOSO3]− | 1656.0/96.9 | −125 | −21 | |
| [M − 3H]3−/[HOSO3]− | 1091.5/96.9 | −125 | −21 | |
| [M + Na − 4H]3−/[HOSO3]− | 1098.6/96.9 | −125 | −21 | |
| [M + K − 4H]3−/[HOSO3]− | 1103.8/96.9 | −125 | −21 | |
| MTX3 | [M − H]−/[HOSO3]− | 1015.5/96.9 | −125 | −21 |
| [M + Na − 2H]−/[HOSO3]− | 1037.6/96.9 | −125 | −21 | |
| [M + 2Na − 3H]−/[HOSO3]− | 1057.5/96.9 | −125 | −21 | |
| [M − 2H]2−/[HOSO3]− | 507.3/96.9 | −125 | −21 | |
| MTX4 | [M − 2H]2−/[M − 2H]2− | 1646.2/1646.2 | −40 | −15 |
| [M − 2H]2−/[HOSO3]− | 1646.2/96.9 | −125 | −21 | |
| [M + Na − 3H]2−/[HOSO3]− | 1657.2/96.9 | −125 | −21 | |
| [M + 2Na − 3H]2−/[HOSO3]− | 1668.2/96.9 | −125 | −21 | |
| [M − 3H]3−/[M − 3H]3− | 1097.1/1097.1 | −40 | −15 | |
| [M − 3H]3−/[HOSO3]− | 1097.1/96.9 | −125 | −21 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. https://doi.org/10.3390/md15070220
Pisapia F, Sibat M, Herrenknecht C, Lhaute K, Gaiani G, Ferron P-J, Fessard V, Fraga S, Nascimento SM, Litaker RW, et al. Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus. Marine Drugs. 2017; 15(7):220. https://doi.org/10.3390/md15070220
Chicago/Turabian StylePisapia, Francesco, Manoëlla Sibat, Christine Herrenknecht, Korian Lhaute, Greta Gaiani, Pierre-Jean Ferron, Valérie Fessard, Santiago Fraga, Silvia M. Nascimento, R. Wayne Litaker, and et al. 2017. "Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus" Marine Drugs 15, no. 7: 220. https://doi.org/10.3390/md15070220
APA StylePisapia, F., Sibat, M., Herrenknecht, C., Lhaute, K., Gaiani, G., Ferron, P. -J., Fessard, V., Fraga, S., Nascimento, S. M., Litaker, R. W., Holland, W. C., Roullier, C., & Hess, P. (2017). Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus. Marine Drugs, 15(7), 220. https://doi.org/10.3390/md15070220