
Genome Sequence of Pseudomonas stutzeri 273 and Identification of the Exopolysaccharide EPS273 Biosynthesis Locus


Abstract
:1. Introduction
2. Results
2.1. Genome Properties of P. stutzeri 273
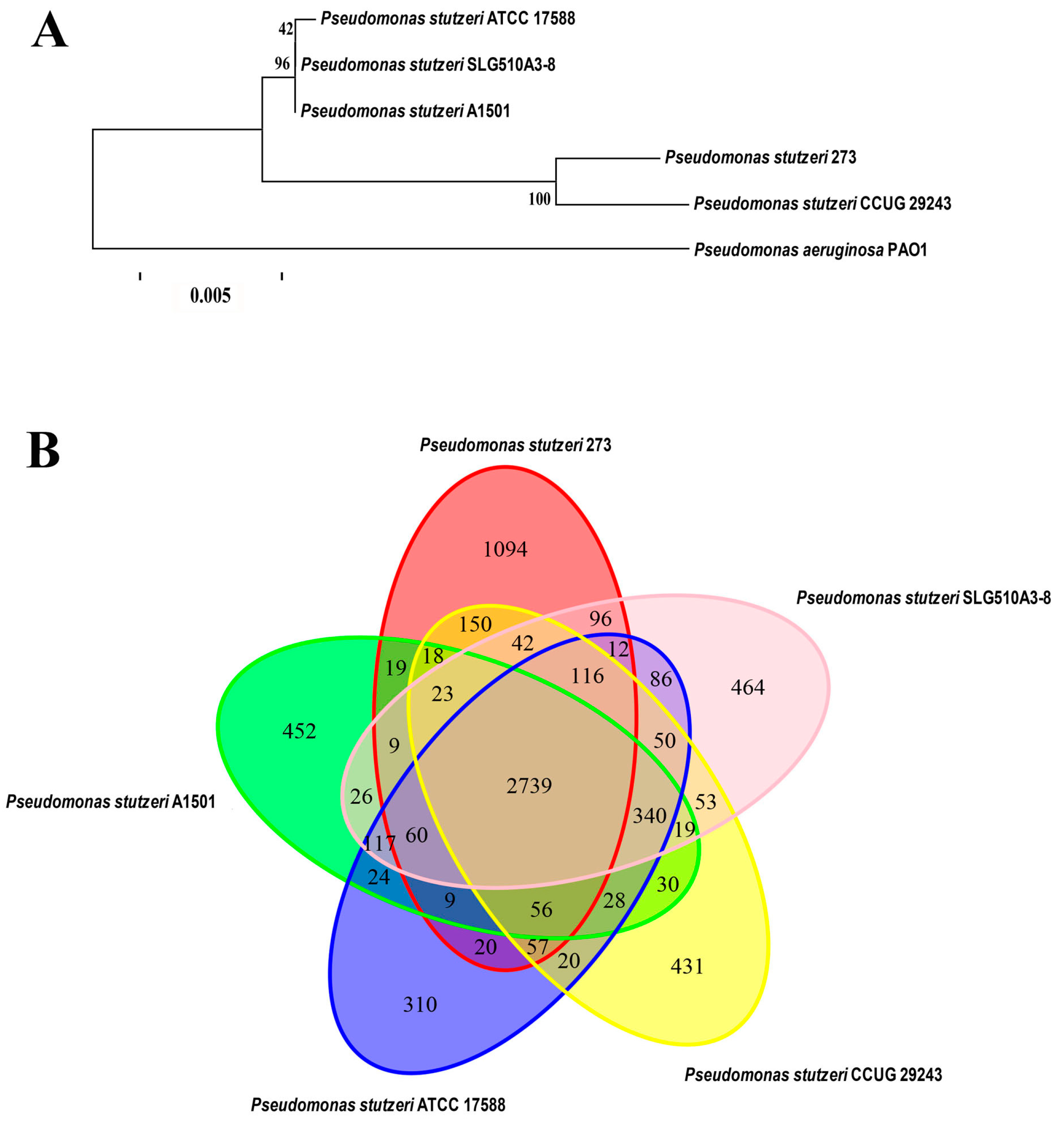
2.2. Insights from the Genome Sequence of P. stutzeri 273
2.3. Identification of EPS273 Biosynthesis Locus in the P. stutzeri 273 Genome
2.4. Insertional Inactivation of Genes Involved in EPS273 Biosynthesis
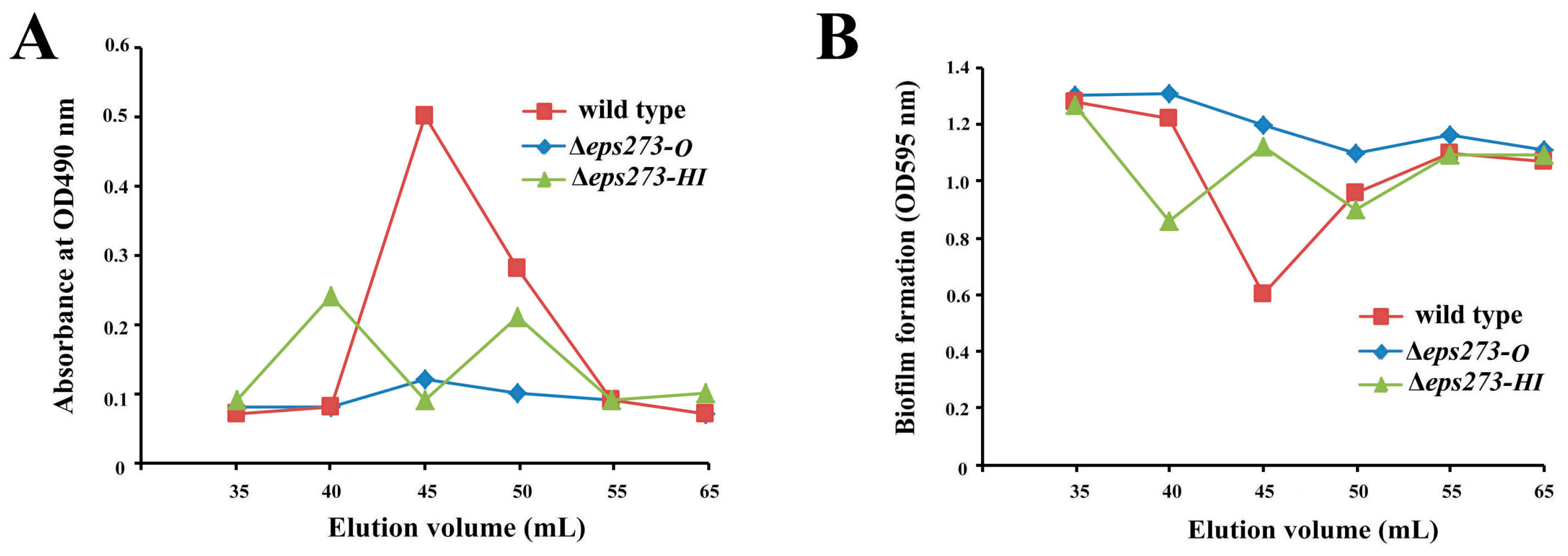
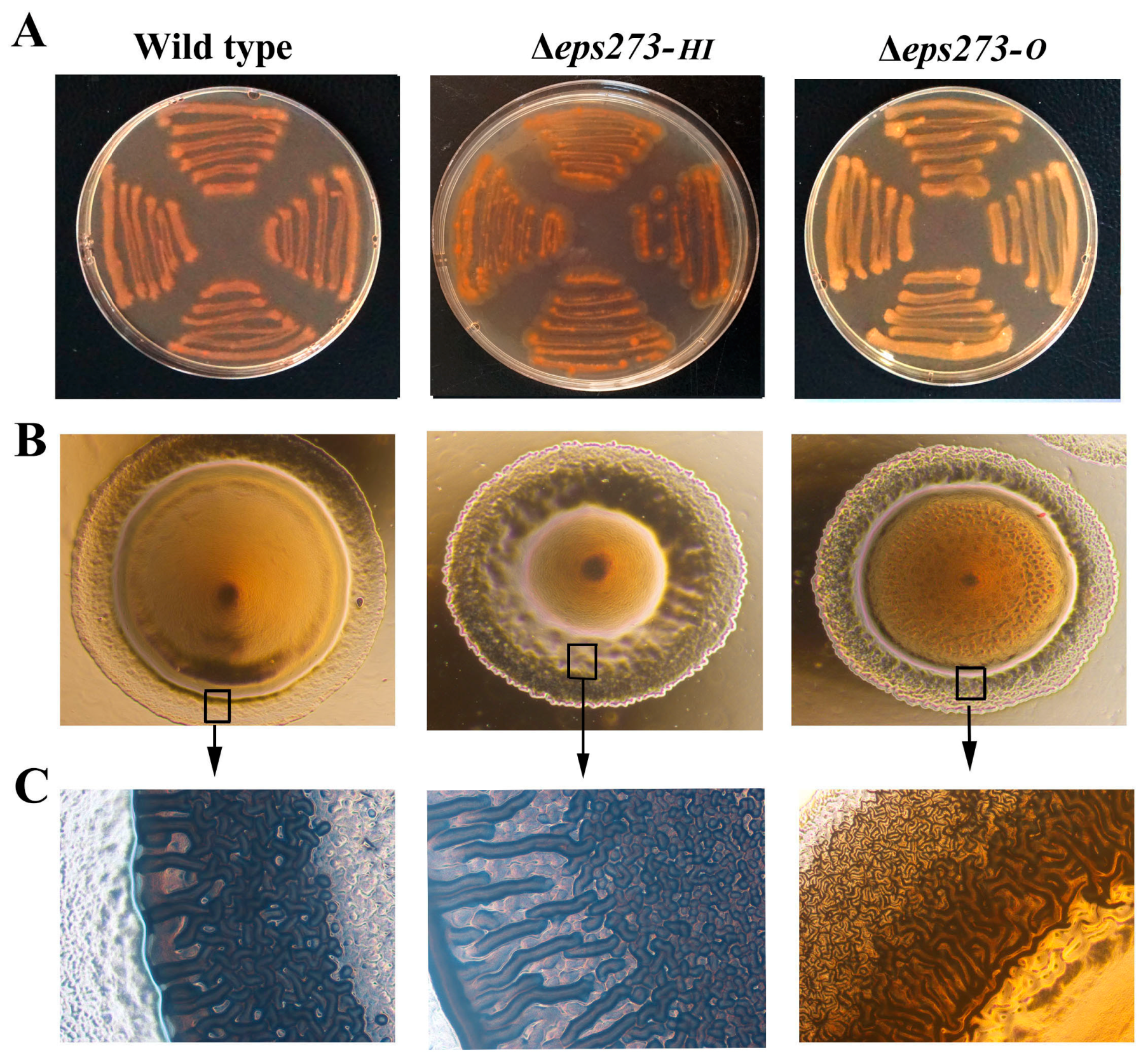
2.5. The Differences of Colony Phenotype and Biofilm Formation between P. stutzeri 273 and Its Mutants Δeps273-HI and Δeps273-O
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Media
4.2. Genomic Studies
4.3. Vector Construction and Mutagenesis
4.4. EPS Extraction, Purification, and Analysis
4.5. Anti-Biofilm Assays
4.6. Colony Morphology on Congo Red (CR) Plates
4.7. Biofilm Formation of P. stutzeri 273 and Its Derivatives Δeps273-HI or Δeps273-O
4.8. Nucleotide Sequence Accession Numbers
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sutherland, I.W. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Boels, I.C.; Ramos, A.; Kleerebezem, M.; De Vos, W.M. Functional analysis of the Lactococcus lactis galU and galE genes and their impact on sugar nucleotide and exopolysaccharide biosynthesis. Appl. Environ. Microb. 2001, 67, 3033–3040. [Google Scholar] [CrossRef] [PubMed]
- Knoshaug, E.P.; Ahlgren, J.A.; Trempy, J.E. Exopolysaccharide expression in Lactococcus lactis subsp cremoris Ropy352: Evidence for novel gene organization. Appl. Environ. Microb. 2007, 73, 897–905. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Degeest, B. Heteropolysaccharides from lactic acid bacteria. FEMS Microbiol. Rev. 1999, 23, 153–177. [Google Scholar] [CrossRef] [PubMed]
- Weigel, P.H.; DeAngelis, P.L. Hyaluronan synthases: A decade-plus of novel glycosyltransferases. J. Biol. Chem. 2007, 282, 36777–36781. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C. Biosynthesis and assembly of capsular polysaccharides in Escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.N.; Whitfield, C. Functional analysis of conserved gene products involved in assembly of Escherichia coli capsules and exopolysaccharides: Evidence for molecular recognition between Wza and Wzc for colanic acid biosynthesis. J. Bacteriol. 2005, 187, 5470–5481. [Google Scholar] [CrossRef] [PubMed]
- Elsholz, A.K.; Wacker, S.A.; Losick, R. Self-regulation of exopolysaccharide production in Bacillus subtilis by a tyrosine kinase. Genes Dev. 2014, 28, 1710–1720. [Google Scholar] [CrossRef] [PubMed]
- Grangeasse, C.; Nessler, S.; Mijakovic, I. Bacterial tyrosine kinases: Evolution, biological function and structural insights. Philos. Trans. R. Soc. B 2012, 367, 2640–2655. [Google Scholar] [CrossRef] [PubMed]
- Rossello, R.; Garciavaldes, E.; Lalucat, J.; Ursing, J. Genotypic and phenotypic diversity of Pseudomonas Stutzeri. Syst. Appl. Microbiol. 1991, 14, 150–157. [Google Scholar] [CrossRef]
- Lalucat, J.; Bennasar, A.; Bosch, R.; Garcia-Valdes, E.; Palleroni, N.J. Biology of Pseudomonas stutzeri. Microbiol. Mol. Biol. Rev. 2006, 70, 510–547. [Google Scholar] [CrossRef] [PubMed]
- Mendez, V.; Fuentes, S.; Hernandez, M.; Morgante, V.; Gonzalez, M.; Seeger, M. Isolation of hydrocarbon-degrading heavy-metal-resistant bacteria from crude oil-contaminated soil in central chile. J. Biotechnol. 2010, 150, S287. [Google Scholar] [CrossRef]
- Wu, S.M.; Liu, G.; Jin, W.H.; Xiu, P.Y.; Sun, C.M. Antibiofilm and anti-infection of a marine bacterial exopolysaccharide against Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Sieber, V.; Rehm, B. Bacterial exopolysaccharides: Biosynthesis pathways and engineering strategies. Front. Microbiol. 2015, 6, 496. [Google Scholar] [CrossRef] [PubMed]
- Brunet-Galmes, I.; Busquets, A.; Pena, A.; Gomila, M.; Nogales, B.; Garcia-Valdes, E.; Lalucat, J.; Bennasar, A.; Bosch, R. Complete genome sequence of the naphthalene-degrading bacterium Pseudomonas stutzeri AN10 (CCUG 29243). J. Bacteriol. 2012, 194, 6642–6643. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Nie, Y.; Geng, S.; Wu, X.L. Complete genome sequence of the petroleum-emulsifying bacterium Pseudomonas stutzeri SLG510A3-8. J. Biotechnol. 2015, 211, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yan, Y.L.; Zhang, W.; Lu, W.; Wang, J.; Ping, S.Z.; Lin, M. Complete genome sequence of the type strain Pseudomonas stutzeri CGMCC 1.1803. J. Bacteriol. 2011, 193, 6095. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.L.; Yang, J.; Dou, Y.T.; Chen, M.; Ping, S.Z.; Peng, J.P.; Lu, W.; Zhang, W.; Yao, Z.Y.; Li, H.Q.; et al. Nitrogen fixation island and rhizosphere competence traits in the genome of root-associated Pseudomonas stutzeri A1501. Proc. Natl. Acad. Sci. USA 2008, 105, 7564–7569. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and open software for comparing large genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.S. Improved Pairwise Alignment of Genomic DNA. Ph.D. Thesis, Pennsylvania State University, State College, PA, USA, 2007. [Google Scholar]
- Nandi, T.; Ong, C.; Singh, A.P.; Boddey, J.; Atkins, T.; Sarkar-Tyson, M.; Essex-Lopresti, A.E.; Chua, H.H.; Pearson, T.; Kreisberg, J.F.; et al. A Genomic Survey of Positive Selection in Burkholderia pseudomallei Provides Insights into the Evolution of Accidental Virulence. PLoS Pathog. 2010, 6, e1000845. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Z.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Yip, E.S.; Quirke, K.P.; Ondrey, J.M.; Visick, K.L. Roles of the structural symbiosis polysaccharide (syp) genes in host colonization, biofilm formation, and polysaccharide biosynthesis in Vibrio fischeri. J. Bacteriol. 2012, 194, 6736–6747. [Google Scholar] [CrossRef] [PubMed]
- Goudenege, D.; Boursicot, V.; Versigny, T.; Bonnetot, S.; Ratiskol, J.; Sinquin, C.; LaPointe, G.; Le Roux, F.; Delbarre-Ladrat, C. Genome sequence of Vibrio diabolicus and identification of the exopolysaccharide HE800 biosynthesis locus. Appl. Microbiol. Biotechnol. 2014, 98, 10165–10176. [Google Scholar] [CrossRef] [PubMed]
- Ryjenkov, D.A.; Tarutina, M.; Moskvin, O.V.; Gomelsky, M. Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: Insights into biochemistry of the GGDEF protein domain. J. Bacteriol. 2005, 187, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.K.; Detweiler, C.S.; Falkow, S. OmpR regulates the two-component system SsrA-SsrB in Salmonella pathogenicity island 2. J. Bacteriol. 2000, 182, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Belogurov, G.A.; Sevostyanova, A.; Svetlov, V.; Artsimovitch, I. Functional regions of the N-terminal domain of the antiterminator RfaH. Mol. Microbiol. 2010, 76, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Awram, P.; Smit, J. Identification of lipopolysaccharide O antigen synthesis genes required for attachment of the S-layer of Caulobacter crescentus. Microbiology 2001, 147, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Kido, N.; Torgov, V.I.; Sugiyama, T.; Uchiya, K.; Sugihara, H.; Komatsu, T.; Kato, N.; Jann, K. Expression of the O9 Polysaccharide of Escherichia-coli: Sequencing of the Escherichia coli O9 Rfb Gene-Cluster, Characterization of Mannosyl Transferases, and Evidence for an Atp-Binding Cassette Transport-System. J. Bacteriol. 1995, 177, 2178–2187. [Google Scholar] [CrossRef] [PubMed]
- Ruberg, S.; Puhler, A.; Becker, A. Biosynthesis of the exopolysaccharide galactoglucan in Sinorhizobium meliloti is subject to a complex control by the phasphate-dependent regulator PhoB and the proteins ExpG and MucR. Microbiology 1999, 145, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Cole, R.A.; Reeves, P.R. An O-antigen processing function for Wzx (RfbX): A promising candidate for O-unit flippase. J. Bacteriol. 1996, 178, 2102–2107. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Tao, J.; Guo, H.J.; Xu, J.G.; Li, Y.Y.; Rezwan, F.; Reeves, P.; Wang, L. Structure of the Shigella dysenteriae 7 O antigen gene cluster and identification of its antigen specific genes. Microb. Pathog. 2004, 36, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.Y.; Lee, H.J.; Yang, Y.H.; Joo, H.S.; Kim, B.G. Characterization and investigation of substrate specificity of the sugar aminotransferase WecE from E-coli K12. Chem. Biol. 2004, 11, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Stuart, R.K.; Brahamsha, B.; Busby, K.; Palenik, B. Genomic island genes in a coastal marine Synechococcus strain confer enhanced tolerance to copper and oxidative stress. ISME J. 2013, 7, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Dobrindt, U.; Hochhut, B.; Hentschel, U.; Hacker, J. Genomic islands in pathogenic and environmental microorganisms. Nat. Rev. Microbiol. 2004, 2, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Gerwig, J.; Kiley, T.B.; Gunka, K.; Stanley-Wall, N.; Stulkel, J. The protein tyrosine kinases EpsB and PtkA differentially affect biofilm formation in Bacillus subtilis. Microbiology 2014, 160, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Carneiro, M.O.; Schatz, M.C. The advantages of SMRT sequencing. Genome Biol. 2013, 14, 405. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.; Rodland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Magrane, M.; Consortium, U. UniProt Knowledgebase: A hub of integrated protein data. Database (Oxf.) 2011, 2011, bar009. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.; Lalansingh, C.M.; Babaran, H.E.; Wang, Z.Y.; Prokopec, S.D.; Fox, N.S.; Boutros, P.C. VennDiagramWeb: A web application for the generation of highly customizable Venn and Euler diagrams. BMC Bioinform. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Ramulu, H.G.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.L.; Wang, Y.S.; Qian, D.Y.; Fan, J.Q.; Hu, B.S.; Liu, F.Q. Selection of available suicide vectors for gene mutagenesis using chiA (a chitinase encoding gene) as a new reporter and primary functional analysis of chiA in Lysobacter enzymogenes strain OH11. World J. Microb. Biotechnol. 2012, 28, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Li, J.; Han, F.; Duan, G.; Lu, X.; Gu, Y.; Yu, W. Antibiofilm activity of an exopolysaccharide from marine bacterium Vibrio sp. QY101. PLoS ONE 2011, 6, e18514. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.M.; Liu, G.; Zhang, D.C.; Li, C.X.; Sun, C.M. Purification and biochemical characterization of an alkaline protease from marine bacteria Pseudoalteromonas sp 129-1. J. Basic Microb. 2015, 55, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.; Li, X.H.; Halverson, L.J. Cell-cell and cell-surface interactions mediated by cellulose and a novel exopolysaccharide contribute to Pseudomonas putida biofilm formation and fitness under water-limiting conditions. Environ. Microbiol. 2011, 13, 1342–1356. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Protein Designation | Size (Amino Acids) | Putative Function |
|---|---|---|---|
| WP_064482084.1 | EPS273-A | 534 | Diguanylate cyclase |
| WP_045428334.1 | EPS273-B | 492 | Sensor kinase |
| WP_064482085.1 | EPS273-C | 136 | Response regulator |
| WP_064482094.1 | EPS273-M | 175 | Transcription activator |
| WP_064482090.1 | EPS273-H | 405 | Glycosyltransferase family 1 |
| WP_064482091.1 | EPS273-I | 372 | Glycosyltransferase family 1 |
| WP_064482093.1 | EPS273-L | 419 | Glycosyltransferase family 1 |
| WP_064482087.1 | EPS273-E | 332 | Glycosyltransferase family 2 |
| WP_045428298.1 | EPS273-P | 309 | Glycosyltransferase family 2 |
| WP_064482086.1 | EPS273-D | 356 | Acetyltransferase |
| WP_064482092.1 | EPS273-J | 445 | Glycosyl hydrolase |
| WP_064482803.1 | EPS273-K | 417 | O-antigen ligase |
| WP_064482095.1 | EPS273-N | 247 | O-antigen production |
| WP_064482096.1 | EPS273-O | 736 | Tyrosine protein kinase |
| WP_064482098.1 | EPS273-R | 393 | Aminotransferase DegT |
| WP_064482088.1 | EPS273-F | 74 | Hypothetical protein |
| WP_064482089.1 | EPS273-G | 108 | Hypothetical protein |
| WP_064482097.1 | EPS273-Q | 508 | O-antigen unit translocase or flippase (RfbX/Wzx) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Zheng, R.; Sha, Z.; Sun, C. Genome Sequence of Pseudomonas stutzeri 273 and Identification of the Exopolysaccharide EPS273 Biosynthesis Locus. Mar. Drugs 2017, 15, 218. https://doi.org/10.3390/md15070218
Wu S, Zheng R, Sha Z, Sun C. Genome Sequence of Pseudomonas stutzeri 273 and Identification of the Exopolysaccharide EPS273 Biosynthesis Locus. Marine Drugs. 2017; 15(7):218. https://doi.org/10.3390/md15070218
Chicago/Turabian StyleWu, Shimei, Rikuan Zheng, Zhenxia Sha, and Chaomin Sun. 2017. "Genome Sequence of Pseudomonas stutzeri 273 and Identification of the Exopolysaccharide EPS273 Biosynthesis Locus" Marine Drugs 15, no. 7: 218. https://doi.org/10.3390/md15070218
APA StyleWu, S., Zheng, R., Sha, Z., & Sun, C. (2017). Genome Sequence of Pseudomonas stutzeri 273 and Identification of the Exopolysaccharide EPS273 Biosynthesis Locus. Marine Drugs, 15(7), 218. https://doi.org/10.3390/md15070218