Difference in Uptake of Tetrodotoxin and Saxitoxins into Liver Tissue Slices among Pufferfish, Boxfish and Porcupinefish

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
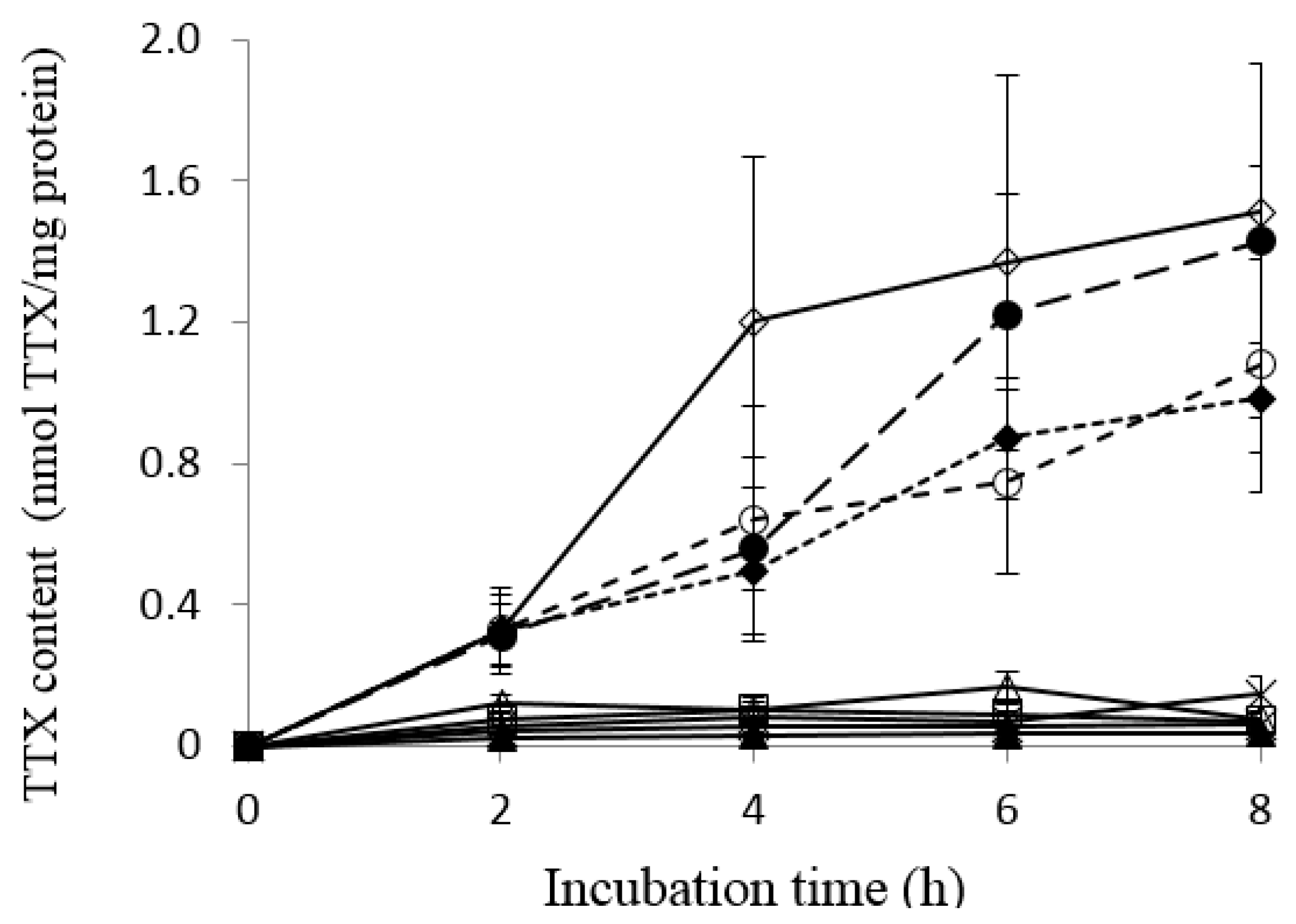
2.1. Uptake of TTX into Liver Tissue Slices of Pufferfish, Boxfish and Porcupinefish
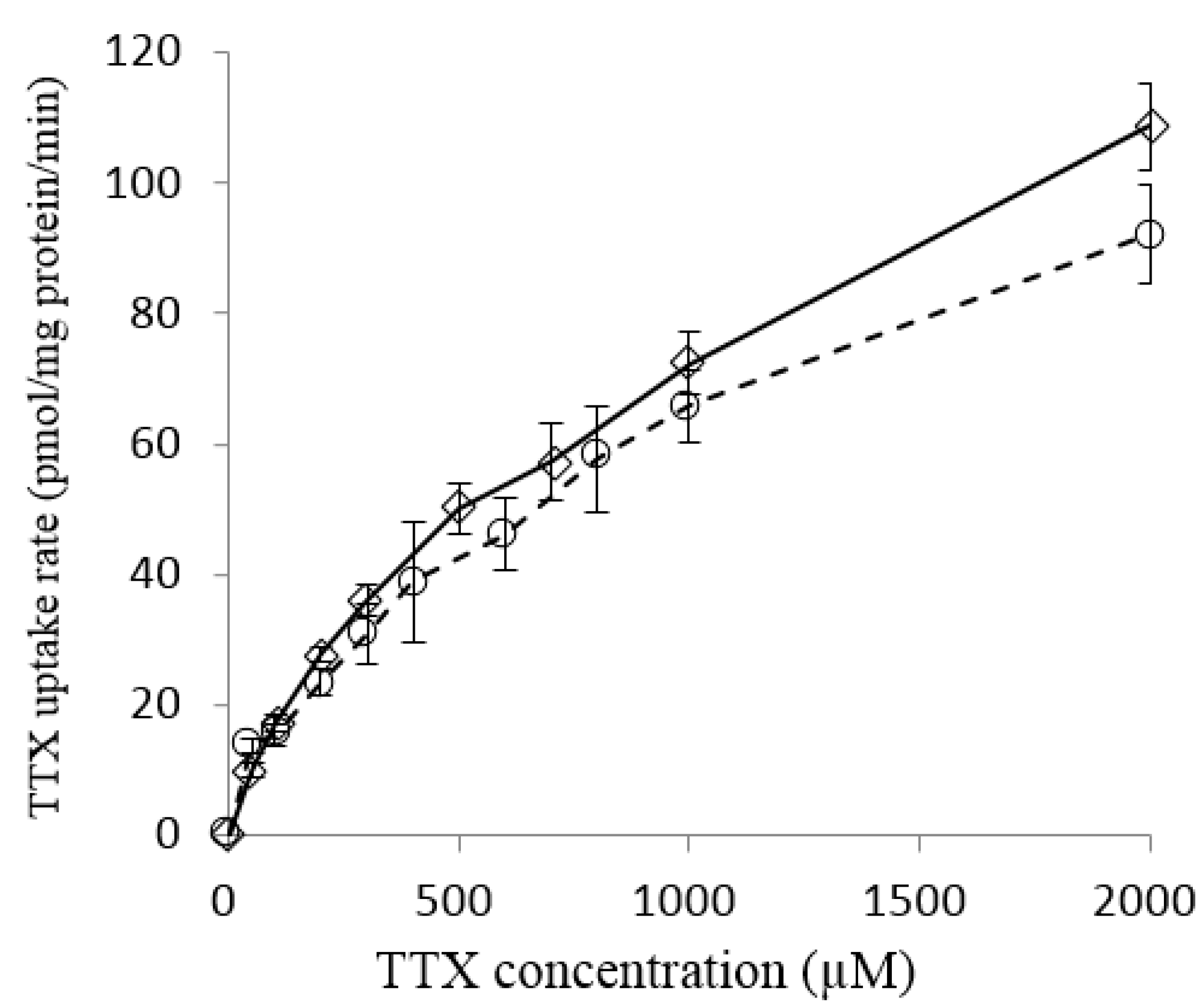
2.2. Effect of TTX Concentration on TTX Uptake into Liver Tissue Slices of L. spadiceus
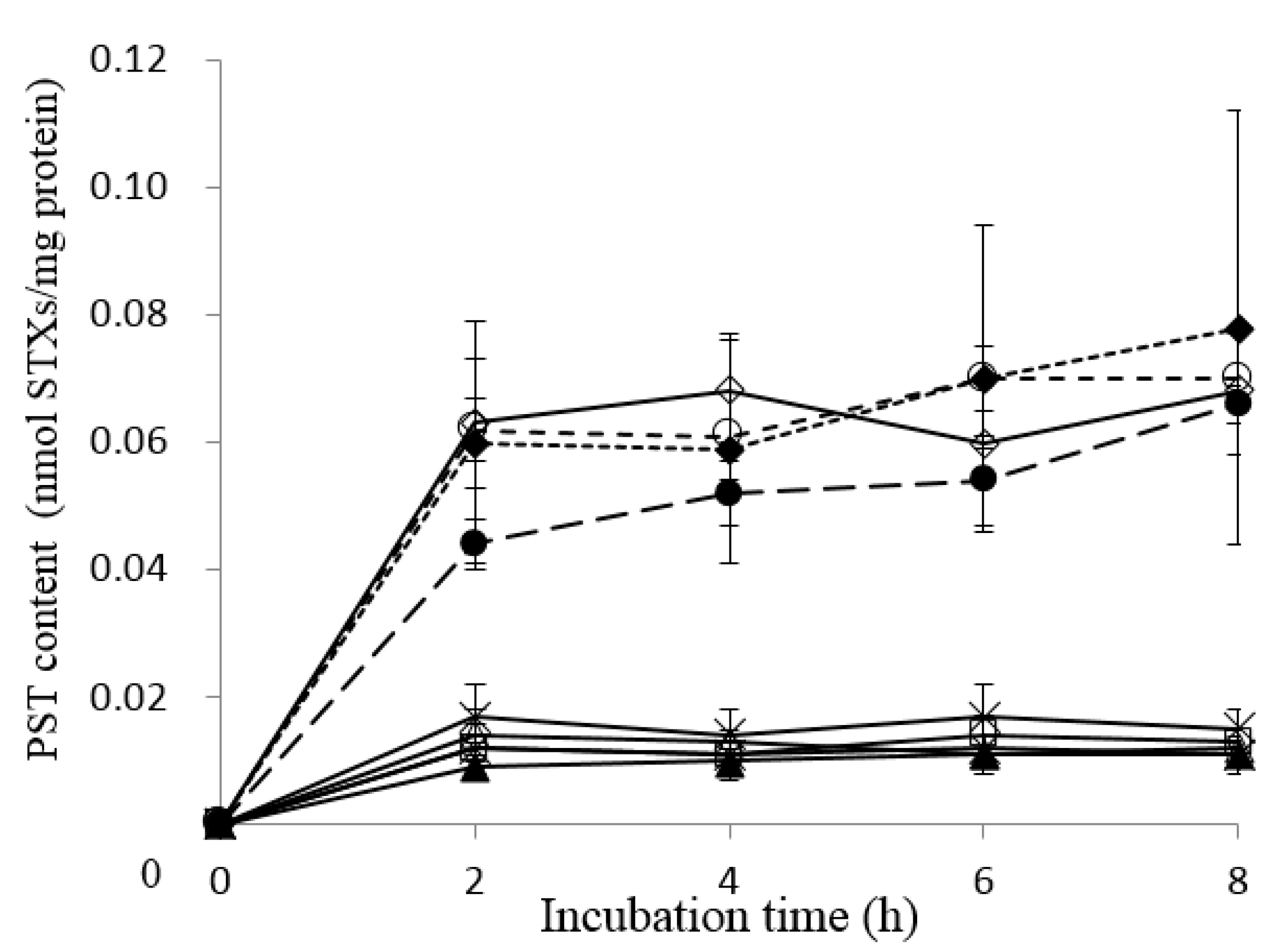
2.3. Uptake of PSTs into Liver Tissue Slices of Pufferfish, Boxfish and Porcupinefish
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. In Vitro Incubation Experiment of Liver Tissue Slices
4.3. Determination of TTX and PSTs
4.4. Determination of Protein
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Noguchi, T.; Arakawa, O. Tetrodotoxin-distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 28, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K. Taxonomic and nomenclatural comments on two puffers of the genus Takifugu with description of a new species, Takifugu flavipterus, from Japan (Actinopterygii, Tetraodontiformes, Tetraodontidae). Bull. Natl. Mus. Nat. Sci. Ser. A 2017, 43, 71–80. [Google Scholar]
- Matsui, T.; Hamada, S.; Konosu, S. Difference in accumulation of puffer fish toxin and crystalline tetrodotoxin in the puffer fish, Fugu rubripes rubripes. Nippon Suisan Gakkaishi 1981, 47, 535–537. [Google Scholar] [CrossRef]
- Matsui, T.; Sato, H.; Hamada, S.; Shimizu, C. Comparison of toxicity of the cultured and wild puffer fish Fugu niphobles. Nippon Suisan Gakkaishi 1982, 48, 253. [Google Scholar] [CrossRef]
- Yamamori, K.; Kono, M.; Furukawa, K.; Matsui, T. The toxification of juvenile cultured kusafugu Takifugu niphobles by oral administration of crystalline tetrodotoxin. J. Food Hyg. Soc. Jpn. (Shokuhin Eiseigaku Zasshi) 2004, 45, 73–75. (In Japanese) [Google Scholar] [CrossRef]
- Honda, S.; Arakawa, O.; Takatani, T.; Tachibana, K.; Yagi, M.; Tanigawa, A.; Noguchi, T. Toxification of cultured puffer fish Takifugu rubripes by feeding on tetrodotoxin-containing diet. Nippon Suisan Gakkaishi 2005, 71, 815–820. (In Japanese) [Google Scholar] [CrossRef]
- Kono, M.; Matsui, T.; Furukawa, K.; Yotsu-Yamashita, M.; Yamamori, K. Accumulation of tetrodotoxin and 4,9-anhydrotetrodotoxin in cultured juvenile kusafugu Fugu niphobles by dietary administration of natural toxic komonfugu Fugu poecilonotus liver. Toxicon 2008, 51, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. Toxicity of pufferfish Takifugu rubripes cultured in netcages at sea or aquaria on land. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Arakawa, O.; Taniyama, S.; Nonaka, M.; Takatani, T.; Yamamori, K.; Noguchi, T. Occurrence of saxitoxins as a major toxin in the ovary of a marine puffer Arothron firmamentum. Toxicon 2004, 43, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Ogata, T.; Borja, V.; Gonzales, C.; Fukuyo, Y.; Kodama, M. Frequent occurrence of paralytic shellfish poisoning toxins as dominant toxins in marine puffer from tropical water. Toxicon 2000, 38, 1101–1109. [Google Scholar] [CrossRef]
- Kungsuwan, A.; Arakawa, O.; Promdet, M.; Onoue, Y. Occurrence of paralytic shellfish poisons in Thai freshwater puffers. Toxicon 1997, 35, 1341–1346. [Google Scholar] [CrossRef]
- Zaman, L.; Arakawa, O.; Shimosu, A.; Onoue, Y. Occurrence of paralytic shellfish poison in Bangladeshi freshwater puffers. Toxicon 1997, 35, 423–431. [Google Scholar] [CrossRef]
- Sato, S.; Kodama, M.; Ogata, T.; Saitanu, K.; Fukuyo, Y.; Hirayama, K.; Kakinuma, K. Saxitoxin as a toxic principle of a freshwater puffer, Tetraodon fangi in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Update: Neurologic illness associated with eating Florida pufferfish, 2002. MMWR Morb. Mortal. Wkly. Rep. 2002, 51, 414–416. [Google Scholar]
- Kodama, M.; Ogata, T.; Noguchi, T.; Maruyama, J.; Hashimoto, K. Occurrence of saxitoxin and other toxins in the liver of the pufferfish Takifugu pardalis. Toxicon 1983, 21, 897–900. [Google Scholar] [CrossRef]
- Jang, J.; Yotsu-Yamashita, M. Distribution of tetrodotoxin, saxitoxin, and their analogs among tissues of the puffer fish Fugu pardalis. Toxicon 2006, 48, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, Y.; Toyoda, M.; Hasobe, M.; Shimakura, K.; Shiomi, K. In vitro accumulation of tetrodotoxin in pufferfish liver tissue slices. Toxicon 2003, 41, 569–574. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagashima, Y.; Takayama, K.; Shimakura, K.; Shiomi, K. Difference between tetrodotoxin and saxitoxins in accumulation in puffer fish Takifugu rubripes liver tissue slices. Fish Physiol. Biochem. 2005, 31, 95–100. [Google Scholar]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Sugiyama, Y.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Involvement of carrier-mediated transport system in uptake of tetrodotoxin into liver tissue slices of puffer fish Takifugu rubripes. Toxicon 2007, 50, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Satoh, T.P. Redescription of Lagocephalus cheesemanii (Clarke 1897), a senior synonym of Lagocephalus gloveri Abe and Tabeta 1983, based on morphological and genetic comparisons (Actinopterygii: Tetraodontiformes: Tetraodontidae). Ichtyol. Res. 2017, 64, 104–110. [Google Scholar] [CrossRef]
- Tani, I. Toxicological Study on Japanese Puffer Fishes; Teikoku Shoin Ltd.: Tokyo, Japan, 1945; pp. 1–103. (In Japanese) [Google Scholar]
- Shiroma, H.; Oshiro, N.; Yamashiro, O.; Tamaki, H. Toxicity of porcupinefish from Okinawa waters. Ann. Rep. Okinawa Prefect. Inst. Health Environ. 1995, 29, 111–117. (In Japanese) [Google Scholar]
- Taniyama, S.; Sagara, T.; Nishio, S.; Kuroki, R.; Asakawa, M.; Noguchi, T.; Yamazaki, S.; Takatani, T.; Arakawa, O. Survey of food poisoning incidents in Japan due to ingestion of marine boxfish, along with their toxicity. J. Food Hyg. Soc. Jpn. (Shokuhin Eiseigaku Zasshi) 2009, 50, 270–277. (In Japanese) [Google Scholar] [CrossRef]
- Oshima, Y. Post-column derivatization HPLC method for the analysis of PSP. J. AOAC Int. 1995, 78, 795–799. [Google Scholar]
- Kiriake, A.; Ohta, A.; Suga, E.; Matsumoto, T.; Ishizaki, S.; Nagashima, Y. Comparison of tetrodotoxin uptake and gene expression in the liver between juvenile and adult pufferfish, Takifugu rubripes. Toxicon 2016, 111, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Endo, R. Toxicological studies on puffer fishes: Comparison of toxicities on the various species. J. Toxicol. Sci. 1984, 9, 1–11. (In Japanese) [Google Scholar] [CrossRef] [PubMed]
- Taniyama, S.; Shibano, K.; Ngy, L.; Shinohara, M.; Takatani, T.; Arakawa, O. Liver toxicity of the pufferfish Lagocephalus wheeleri “shirosabafugu” collected from the coastal waters of Hagi, Yamaguchi Prefecture, Japan. Bull. Fac. Fish. Nagasaki Univ. 2010, 91, 1–3. (In Japanese) [Google Scholar]
- Matsumura, K. Tetrodotoxin as a pheromone. Nature 1995, 378, 563–564. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Yamazaki, H.; Sakiyama, K.; Yamane, H.; Niina, S.; Takatani, T.; Arakawa, O.; Sakakura, Y. Puffer smells tetrodotoxin. Ichtyol. Res. 2013, 60, 386–389. [Google Scholar] [CrossRef]
- Itoi, S.; Yoshikawa, S.; Asahina, K.; Suzuki, M.; Ishizuka, K.; Takimoto, N.; Mitsuoka, R.; Yokoyama, N.; Detake, A.; Takayanagi, C.; et al. Larval pufferfish protected by maternal tetrodotoxin. Toxicon 2014, 78, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Yamamori, K.; Nakamura, M.; Matsui, T.; Hara, T.J. Gustatory responses to tetrodotoxin and saxitoxin in fish: A possible mechanism for avoiding marine toxins. Can. J. Fish. Aquat. Sci. 1988, 45, 2182–2186. [Google Scholar] [CrossRef]
- Saito, T.; Noguchi, T.; Harada, T.; Murata, O.; Abe, T.; Hashimoto, K. Resistibility of toxic and nontoxic pufferfish against tetrodotoxin. Nippon Suisan Gakkaishi 1985, 51, 1371. [Google Scholar] [CrossRef]
- Soong, T.W.; Venkatesh, B. Adaptive evolution of tetrodotoxin resistance in animals. Trends Genet. 2006, 22, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Zakon, H.H.; Jost, M.C.; Zwickl, D.J.; Lu, Y.; Hillis, D.M. Molecular evolution of Na+ channels in teleost fishes. Integr. Zool. 2009, 4, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Kidokoro, Y.; Grinnell, A.D.; Eaton, D.C. Tetrodotoxin sensitivity of muscle action potentials in pufferfishes and related fishes. J. Comp. Physiol. 1974, 89, 59–72. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSTBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immnohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Yamaki, H.; Okoshi, N.; Araki, N. Distribution of homologous proteins to puffer fish saxitoxin and tetrodotoxin binding protein in the plasma of puffer fish and among the tissues of Fugu pardalis examined by Western blot analysis. Toxicon 2010, 55, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Lee, J.M.; Shiraishi, M.; Komatsu, S.; Miki, S.; Shimasaki, Y.; Mochioka, H.; Kusakabe, T.; Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic and nontoxic pufferfishes. J. Evol. Biol. 2015, 28, 1103–1118. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, O.; Noguchi, T.; Onoue, Y. Paralytic shellfish toxin profiles of xanthid crabs Zosimus aeneus and Atergatis floridus collected on reefs of Ishigaki Island. Fish. Sci. 1995, 61, 659–662. [Google Scholar] [CrossRef]
- Kodama, M.; Sato, S. Standard Methods of Analysis in Food Safety Regulation, Chemistry; Ministry of Health, Ed.; Japan Food Hygiene Association: Tokyo, Japan, 2015; pp. 813–820. (In Japanese) [Google Scholar]
- Horwitz, W. (Ed.) Official Methods of Analysis of AOAC International, 17th ed.; AOAC International: Gaithersburg, ML, USA, 2000; pp. 59–61. [Google Scholar]
- Watanabe, R.; Harada, T.; Matsushima, R.; Oikawa, H.; Oshima, Y.; Kaneniwa, M.; Suzuki, T. Indirect quantitation of saxitoxin by HPLC with post-column oxidation and fluorometric detection. Anal. Sci. 2014, 30, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]



 TTX;
TTX;  PSTs.
TTX; PSTs.
PSTs.
TTX; PSTs.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagashima, Y.; Ohta, A.; Yin, X.; Ishizaki, S.; Matsumoto, T.; Doi, H.; Ishibashi, T. Difference in Uptake of Tetrodotoxin and Saxitoxins into Liver Tissue Slices among Pufferfish, Boxfish and Porcupinefish. Mar. Drugs 2018, 16, 17. https://doi.org/10.3390/md16010017
Nagashima Y, Ohta A, Yin X, Ishizaki S, Matsumoto T, Doi H, Ishibashi T. Difference in Uptake of Tetrodotoxin and Saxitoxins into Liver Tissue Slices among Pufferfish, Boxfish and Porcupinefish. Marine Drugs. 2018; 16(1):17. https://doi.org/10.3390/md16010017
Chicago/Turabian StyleNagashima, Yuji, Akira Ohta, Xianzhe Yin, Shoichiro Ishizaki, Takuya Matsumoto, Hiroyuki Doi, and Toshiaki Ishibashi. 2018. "Difference in Uptake of Tetrodotoxin and Saxitoxins into Liver Tissue Slices among Pufferfish, Boxfish and Porcupinefish" Marine Drugs 16, no. 1: 17. https://doi.org/10.3390/md16010017
APA StyleNagashima, Y., Ohta, A., Yin, X., Ishizaki, S., Matsumoto, T., Doi, H., & Ishibashi, T. (2018). Difference in Uptake of Tetrodotoxin and Saxitoxins into Liver Tissue Slices among Pufferfish, Boxfish and Porcupinefish. Marine Drugs, 16(1), 17. https://doi.org/10.3390/md16010017