A Novel Cold-Adapted Leucine Dehydrogenase from Antarctic Sea-Ice Bacterium Pseudoalteromonas sp. ANT178


Abstract
:1. Introduction
2. Results and Discussion
2.1. Gene Cloning and Sequence Analysis
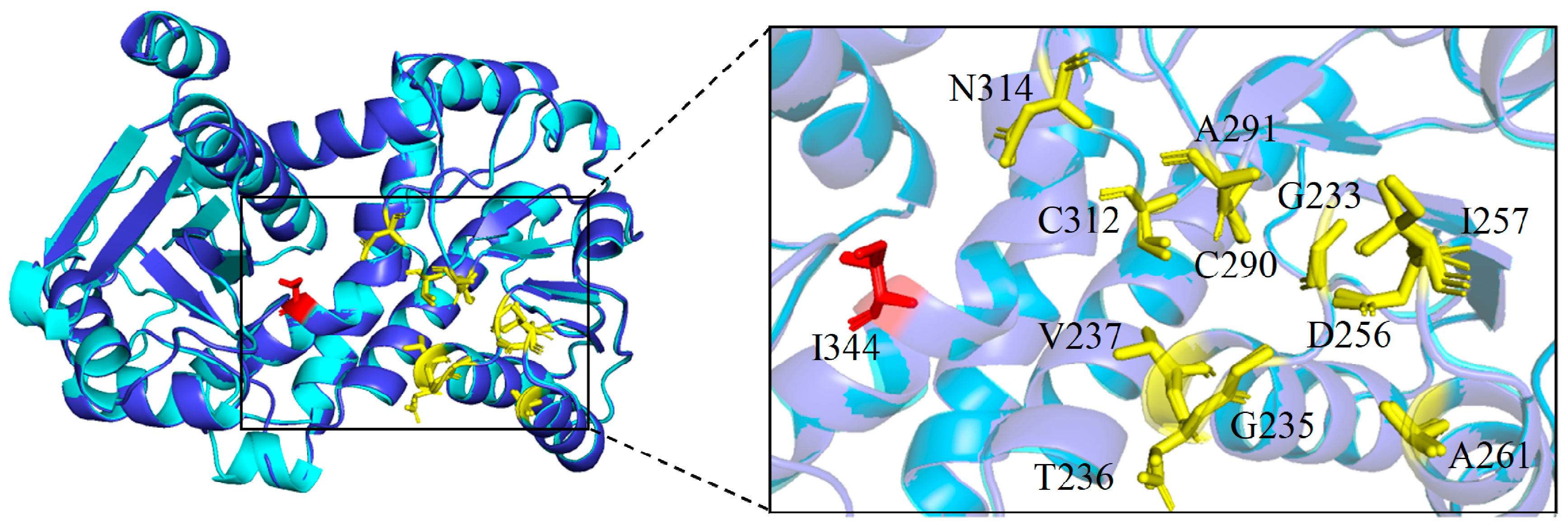
2.2. Homology Modeling and Analysis of PsLeuDH
2.3. Expression and Purification of the rPsLeuDH
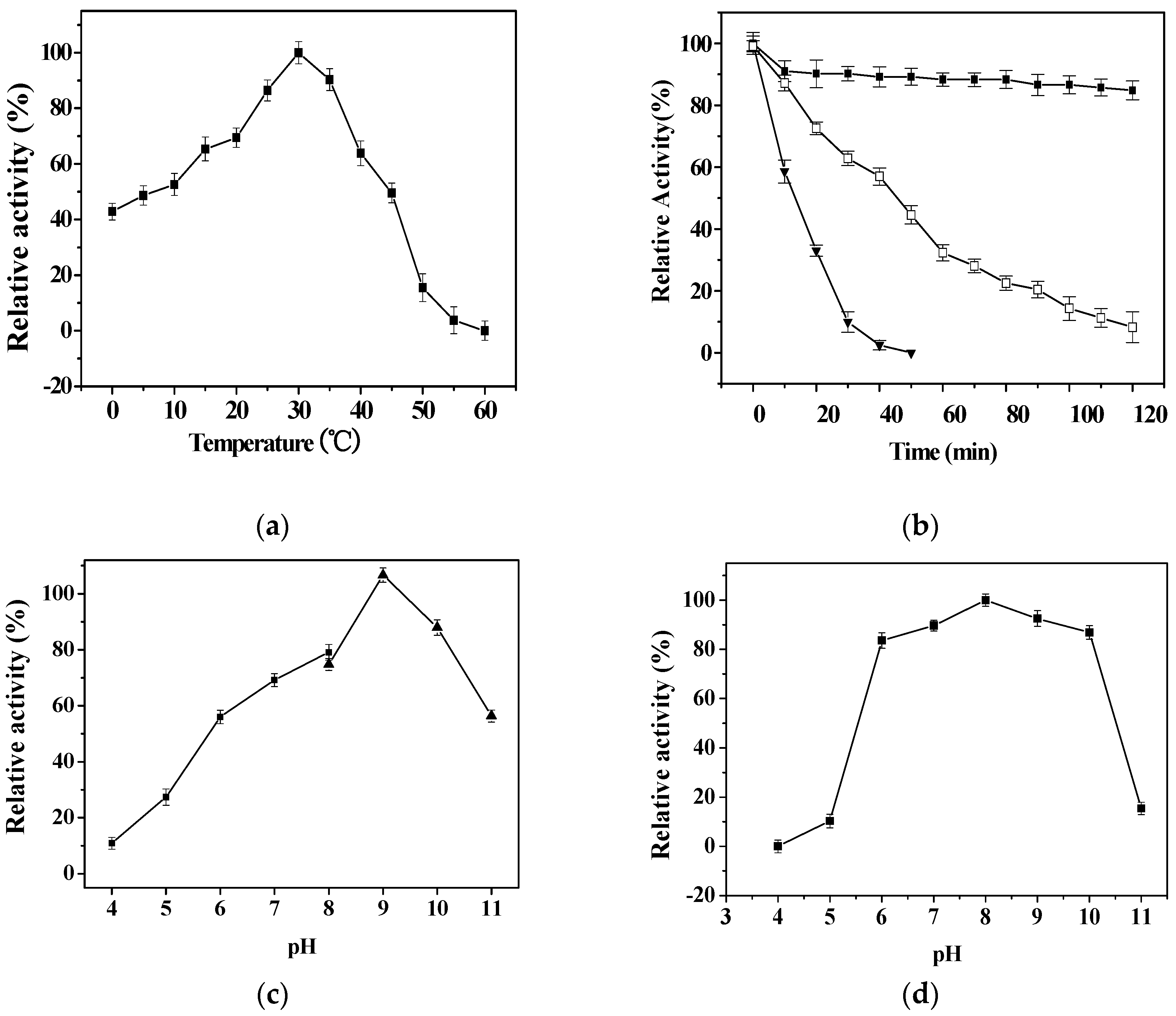
2.4. Effects of Temperature and pH on Activity and Stability of rPsLeuDH
2.5. Effects of NaCl Concentration and Different Reagents on the Activity of PsLeuDH
2.6. The Substrate Specificity Analysis and Kinetic Parameters of rPsLeuDH
2.7. The Thermodynamic Parameters of rPsLeuDH
3. Materials and Methods
3.1. Microorganisms and Growth Conditions
3.2. Sequence Analysis of LeuDH Gene
3.3. Protein Homology Modeling
3.4. Molecular Cloning, Expression and Purification of rPsLeuDH
3.5. Assay of rPsLeuDH Activity
3.6. Characterization of the Purified rPsLeuDH
3.7. Kinetic Parameter of the rPsLeuDH
3.8. Thermodynamic Parameter of the rPsLeuDH
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ohshima, T.; Wandrey, C.; Sugiura, M.; Soda, K. Screening of thermostable leucine and alanine dehydrogenases in thermophilic Bacillus strains. Biotechnol. Lett. 1985, 7, 871–876. [Google Scholar] [CrossRef]
- Sanwal, B.D.; Zink, M.W. l-leucine dehydrogenase of Bacillus cereus. Arch. Biochem. Biophys. 1961, 94, 430–435. [Google Scholar] [CrossRef]
- Nagata, S.; Bakthavatsalam, S.; Galkin, A.G.; Asada, H.; Sakai, S.; Esaki, N.; Soda, K.; Ohshima, T.; Nagasaki, S.; Misono, H. Gene cloning, purification, and characterization of thermostable and halophilic leucine dehydrogenase from a halophilic thermophile, Bacillus licheniformis TSN9. Appl. Microbiol. Biotechnol. 1995, 44, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Katoh, R.; Nagata, S.; Misono, H. Cloning and sequencing of the leucine dehydrogenase gene from Bacillus sphaericus, IFO 3525 and importance of the C-terminal region for the enzyme activity. J. Mol. Catal. B Enzym. 2003, 23, 239–247. [Google Scholar] [CrossRef]
- Mahdizadehdehosta, R.; Kianmehr, A.; Khalili, A. Isolation and characterization of leucine dehydrogenase from a thermophilic Citrobacter freundii JK-91 strain isolated from Jask Port. Iran. J. Microbiol. 2013, 5, 278–284. [Google Scholar] [PubMed]
- Zhu, W.J.; Li, Y.; Jia, H.H.; Wei, P.; Zhou, H.; Jiang, M. Expression, purification and characterization of a thermostable leucine dehydrogenase from the halophilic thermophile Laceyella sacchari. Biotechnol. Lett. 2016, 38, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wakamatsu, T.; Doi, K.; Sakuraba, H.; Ohshima, T. A psychrophilic leucine dehydrogenase from Sporosarcina psychrophila: Purification, characterization, gene sequencing and crystal structure analysis. J. Mol. Catal. B Enzym. 2012, 83, 65–72. [Google Scholar] [CrossRef]
- Turnbull, A.P.; Ashford, S.R.; Baker, P.J.; Rice, D.W.; Rodgers, F.H.; Stillman, T.J.; Hanson, R.L. Crystallization and quaternary structure analysis of the NAD(+)-dependent leucine dehydrogenase from Bacillus sphaericus. J. Mol. Biol. 1994, 236, 663–665. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wu, Z.; Jin, J.M.; Tang, S.Y. Directed evolution of leucine dehydrogenase for improved efficiency of l-tert-leucine synthesis. Appl. Microbiol. Biotechnol. 2016, 100, 5805–5813. [Google Scholar] [CrossRef] [PubMed]
- Galkin, A.; Kulakova, L.; Ashida, H.; Sawa, Y.; Esaki, N. Cold-adapted alanine dehydrogenases from two Antarctic bacterial strains: Gene cloning, protein characterization, and comparison with mesophilic and thermophilic counterparts. Appl. Environ. Microb. 1999, 65, 4014–4020. [Google Scholar]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trend Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Jiang, W.; Sun, D.F.; Lu, J.X.; Wang, Y.L.; Wang, S.Z.; Zhang, Y.H.; Fang, B.S. A cold-adapted leucine dehydrogenase from marine bacterium Alcanivorax dieselolei: Characterization and l-tert-leucine production. Eng. Life Sci. 2016, 16, 283–289. [Google Scholar] [CrossRef]
- Shi, Y.L.; Wang, Q.F.; Hou, Y.H.; Hong, Y.Y.; Han, X.; Yi, J.L.; Qu, J.J.; Lu, Y. Molecular cloning, expression and enzymatic characterization of glutathione s-transferase from Antarctic sea-ice bacteria Pseudoalteromonas sp. ANT506. Microbiol. Res. 2014, 169, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Han, H.; Cui, B.Q.; Hou, Y.H.; Wang, Y.F.; Wang, Q.F. A glutathione peroxidase from Antarctic psychrotrophic bacterium Pseudoalteromonas sp. ANT506: Cloning and heterologous expression of the gene and characterization of recombinant enzyme. Bioengineered 2017, 8, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, S.I.; Tanizawa, K.; Sakamoto, Y.; Tanaka, H.; Soda, K. Alanine dehydrogenases from two Bacillus species with distinct thermostabilities: Molecular cloning, DNA and protein sequence determination, and structural comparison with other NAD(P)(+)-dependent dehydrogenases. Biochemistry 1990, 29, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.J.; Turnbull, A.P.; Sedelnikova, S.E.; Stillman, T.J.; Rice, D.W. A role for quaternary structure in the substrate specificity of leucine dehydrogenase. Structure 1995, 3, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Paredes, D.I.; Watters, K.; Pitman, D.J.; Bystroff, C.; Dordick, J.S. Comparative void-volume analysis of psychrophilic and mesophilic enzymes: Structural bioinformatics of psychrophilic enzymes reveals sources of core flexibility. BMC Struct. Biol. 2011, 11, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Li, F.L.; Shi, Y.; Zhang, J.X.; Gao, J.; Zhang, Y.W. Cloning, expression, characterization and homology modeling of a novel water-forming NADH oxidase from Streptococcus mutans ATCC 25175. Int. J. Biol. Macromol. 2018, 113, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.; Parvizpour, S.; Razmara, J.; Abu Bakar, F.D.; Illias, R.M.; Mahadi, N.M.; Murad, A.M. Structure prediction of a novel Exo-β-1,3-Glucanase: Insights into the cold adaptation of psychrophilic yeast Glaciozyma antarctica PI12. Interdiscip. Sci. Comput. Life Sci. 2016, 10, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Herning, T.; Yutani, K.; Inaka, K.; Kuroki, R.; Matsushima, M.; Kikuchi, M. Role of proline residues in human lysozyme stability: A scanning calorimetric study combined with X-ray structure analysis of proline mutants. Biochemistry 1992, 31, 7077–7085. [Google Scholar] [CrossRef] [PubMed]
- Siglioccolo, A.; Gerace, R.; Pascarella, S. “Cold spots” in protein cold adaptation: Insights from normalized atomic displacement parameters (B′-factors). Biophys. Chem. 2010, 153, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, J.; Zhang, J.; Xu, J.H. Stereoselective synthesis of l-tert-leucine by a newly cloned leucine dehydrogenase from Exiguobacterium sibiricum. J. Mol. Catal. B Enzym. 2014, 105, 11–17. [Google Scholar] [CrossRef]
- Feller, G.; Narinx, E.; Arpigny, J.L.; Aittaleb, M.; Baise, E.; Genicot, S.; Gerday, C. Enzymes from psychrophilic organisms. FEMS Microbiol. Rev. 1996, 18, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, T.; Nishida, N.; Bakthavatsalam, S.; Kataoka, K.; Takada, H.; Yoshimura, T.; Soda, K.; Esaki, N. The purification, characterization, cloning and sequencing of the gene for a halostable and thermostable leucine dehydrogenase from Thermoactinomyces intermedius. Eur. J. Biochem. 1994, 222, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Michetti, D.; Brandsdal, B.O.; Bon, D.; Isaksen, G.V.; Tiberti, M.; Papaleo, E. A comparative study of cold- and warm-adapted endonucleases a using sequence analyses and molecular dynamics simulations. PLoS ONE 2017, 12, e0169586. [Google Scholar] [CrossRef] [PubMed]
- Pawlak-Szukalska, A.; Wanarska, M.; Popinigis, A.T.; Kur, J. A novel cold-active β-d-galactosidase with transglycosylation activity from the Antarctic Arthrobacter sp. 32cB-Gene cloning, purification and characterization. Process Biochem. 2014, 49, 2122–2133. [Google Scholar] [CrossRef]
- Lonhienne, T.; Gerday, C.; Feller, G. Psychrophilic enzymes: Revisiting the thermodynamic parameters of activation may explain local flexibility. Biochim. Biophys. Acta 2000, 1543, 1–10. [Google Scholar] [CrossRef]
- Khrapunov, S.; Chang, E.; Callender, R.H. Thermodynamic and structural adaptation differences between the mesophilic and psychrophilic lactate dehydrogenases. Biochemistry 2017, 56, 3587–3595. [Google Scholar] [CrossRef] [PubMed]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshima, T.; Nagata, S.; Soda, K. Purification and characterization of thermostable leucine dehydrogenase from Bacillus stearothermophilus. Arch. Microbiol. 1985, 141, 407–411. [Google Scholar] [CrossRef]
- Shang, Z.C.; Zhang, L.L.; Wu, Z.J.; Gong, P.; Li, D.P.; Zhu, P.; Gao, H.J. The activity and kinetic parameters of oxidoreductases in phaeozem in response to long-term fertiliser management. J. Soil Sci. Plant Nutr. 2012, 12, 597–607. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | PsLeuDH | Bs-LeuDH | Expected Effect on PsLeuDH |
|---|---|---|---|
| Electrostatic interactions | Protein stability | ||
| Salt Bridge (2.5 to 4.0) | 17 | 22 | |
| Hydrogen Bonds (≤3.3 Å) | 368 | 403 | |
| Cation-pi interactions | 3 | 11 | |
| Aromatic interactions | 6 | 8 | |
| Hydrophobic interactions | 227 | 318 | Thermolability |
| Glycine residues | 42 | 36 | Flexibility |
| Proline residues | 9 | 11 | |
| Arginine residues | 10 | 17 | |
| Glycine substitution (PsLeuDH → BsLeuDH) | G163 → N107, G177 → D121, G238 → A185, G240 → A187, G275 → A222, G401 →V346 | ||
| Proline substitution (PsLeuDH → BsLeuDH) | A94 → P38, A143 →P87, S320 → P267, S385 → P330 | ||
| Proline substitution (PsLeuDH → BsLeuDH) | P131 → N75, P63 → M7 | Stability | |
| Arginine substitution (PsLeuDH → BsLeuDH) | R219 → F166, R264 → A211, R327 → H274, R378 → I323 | ||
| Reagent | Concentration | Relative Activity (%) | Reagent | Concentration | Relative Activity (%) |
|---|---|---|---|---|---|
| None | 100 ± 0.0 | ||||
| KCl | 1 mM | 99.7 ± 1.6 | KCl | 5 mM | 40.0 ± 1.9 |
| CoCl2 | 1 mM | 90.1 ± 1.7 | CoCl2 | 5 mM | 70.0 ± 2.0 |
| MgCl2 | 1 mM | 87.9 ± 0.8 | MgCl2 | 5 mM | 65.8 ± 1.2 |
| CaCl2 | 1 mM | 87.9 ± 0.4 | CaCl2 | 5 mM | 68.1 ± 0.9 |
| ZnCl2 | 1 mM | 80.0 ± 2.5 | ZnCl2 | 5 mM | 72.2 ± 2.0 |
| FeCl2 | 1 mM | 75.1 ± 2.2 | FeCl2 | 5 mM | 62.4 ± 1.7 |
| CuCl2 | 1 mM | 61.0 ± 2.2 | CuCl2 | 5 mM | 41.0 ± 1.5 |
| HgCl2 | 1 mM | 29.2 ± 0.3 | HgCl2 | 5 mM | 12.3 ± 1.9 |
| CrCl2 | 1 mM | 13.3 ± 0.3 | CrCl2 | 5 mM | 5.8 ± 2.9 |
| CdCl2 | 1 mM | 7.6 ± 0.5 | CdCl2 | 5 mM | 0.0 ± 0.0 |
| Pb(NO3)2 | 1 mM | 0.0 ± 0.0 | Pb(NO3)2 | 5 mM | 0.0 ± 0.0 |
| BaCl2 | 1 mM | 0.0 ± 0.0 | BaCl2 | 5 mM | 0.0 ± 0.0 |
| EDTA | 1 mM | 91.8 ± 2.7 | EDTA | 5 mM | 84.2 ± 2.1 |
| Thiourea | 1 mM | 51.5 ± 4.0 | Thiourea | 5 mM | 34.3 ± 2.6 |
| Triton X-100 | 0.2% | 102.7 ± 1.4 | Ethanol | 25% | 67.5 ± 1.4 |
| Substrate | Vm (μmol/min·mg) | Km (mM) | kcat (1/s) | kcat/Km (mM−1 s−1) |
|---|---|---|---|---|
| l-lecine | 15.24 | 0.33 | 30.13 | 91.30 |
| l-tyrosine | 13.35 | 0.48 | 26.39 | 54.98 |
| l-proline | 10.52 | 0.64 | 20.80 | 32.50 |
| dl-methionine | 8.38 | 0.75 | 16.57 | 22.09 |
| l-arginine | 7.13 | 0.84 | 14.09 | 16.77 |
| Temperature (°C) | ΔH (KJ/mol) | ΔS (J/mol K) | ΔG (KJ/mol) | kcat (1/s) |
|---|---|---|---|---|
| 0 | 18.27 | −156.45 | 61.01 | 12.25 |
| 10 | 18.19 | −157.75 | 62.90 | 14.96 |
| 20 | 18.11 | −158.02 | 64.43 | 20.20 |
| 30 | 18.02 | −157.28 | 65.70 | 30.13 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Hou, Y.; Wang, Y.; Zheng, L.; Xu, X.; Pan, K.; Li, R.; Wang, Q. A Novel Cold-Adapted Leucine Dehydrogenase from Antarctic Sea-Ice Bacterium Pseudoalteromonas sp. ANT178. Mar. Drugs 2018, 16, 359. https://doi.org/10.3390/md16100359
Wang Y, Hou Y, Wang Y, Zheng L, Xu X, Pan K, Li R, Wang Q. A Novel Cold-Adapted Leucine Dehydrogenase from Antarctic Sea-Ice Bacterium Pseudoalteromonas sp. ANT178. Marine Drugs. 2018; 16(10):359. https://doi.org/10.3390/md16100359
Chicago/Turabian StyleWang, Yatong, Yanhua Hou, Yifan Wang, Lu Zheng, Xianlei Xu, Kang Pan, Rongqi Li, and Quanfu Wang. 2018. "A Novel Cold-Adapted Leucine Dehydrogenase from Antarctic Sea-Ice Bacterium Pseudoalteromonas sp. ANT178" Marine Drugs 16, no. 10: 359. https://doi.org/10.3390/md16100359
APA StyleWang, Y., Hou, Y., Wang, Y., Zheng, L., Xu, X., Pan, K., Li, R., & Wang, Q. (2018). A Novel Cold-Adapted Leucine Dehydrogenase from Antarctic Sea-Ice Bacterium Pseudoalteromonas sp. ANT178. Marine Drugs, 16(10), 359. https://doi.org/10.3390/md16100359