Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results and Discussion
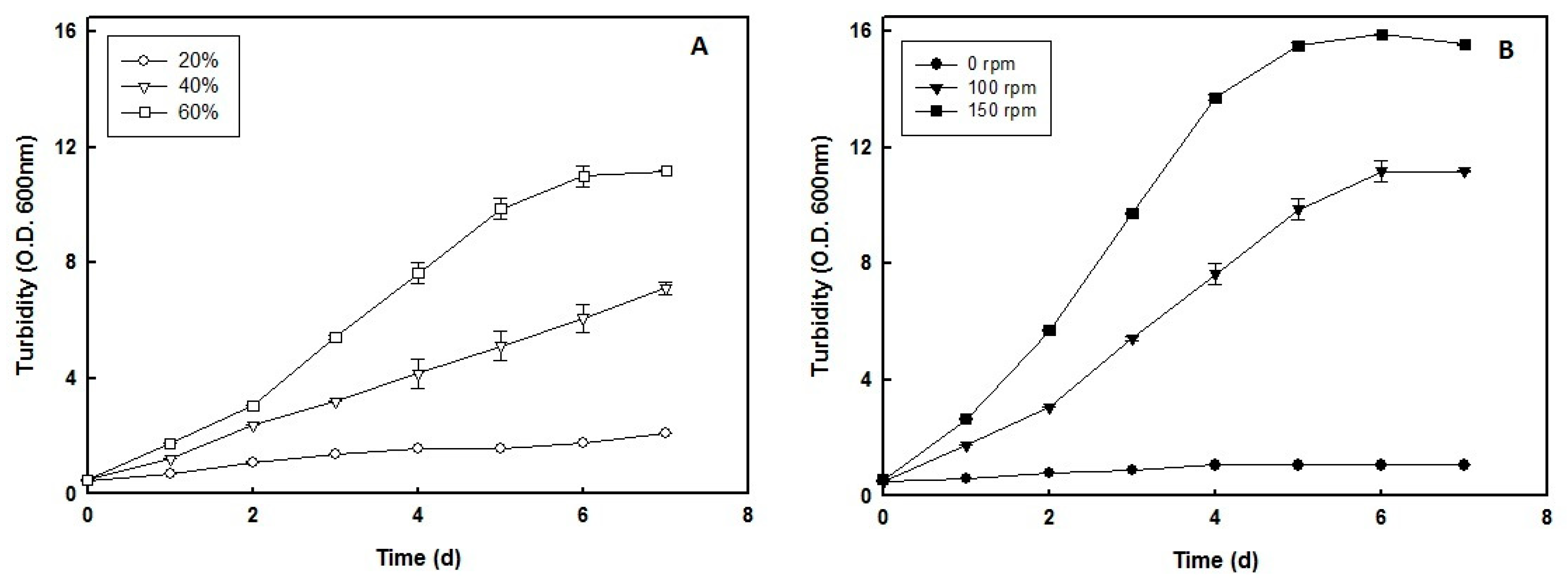
2.1. Effect of Air Volume inside the Culture Flasks and Speed of Agitation on the Growth Rate of H. mediterranei
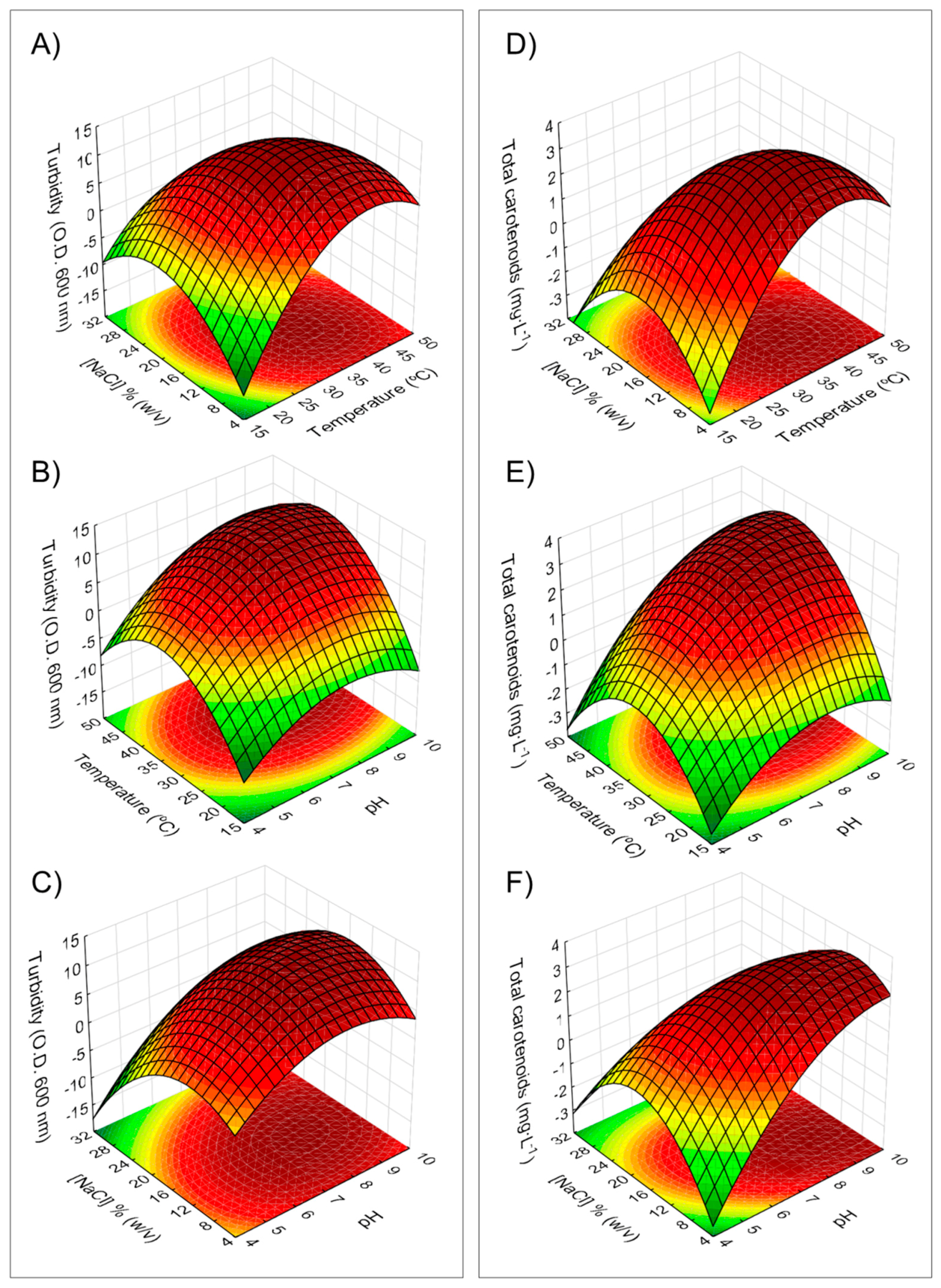
2.2. Use of RSM to Optimize Culture Conditions for Growth and Carotenoid Production by H. mediterranei
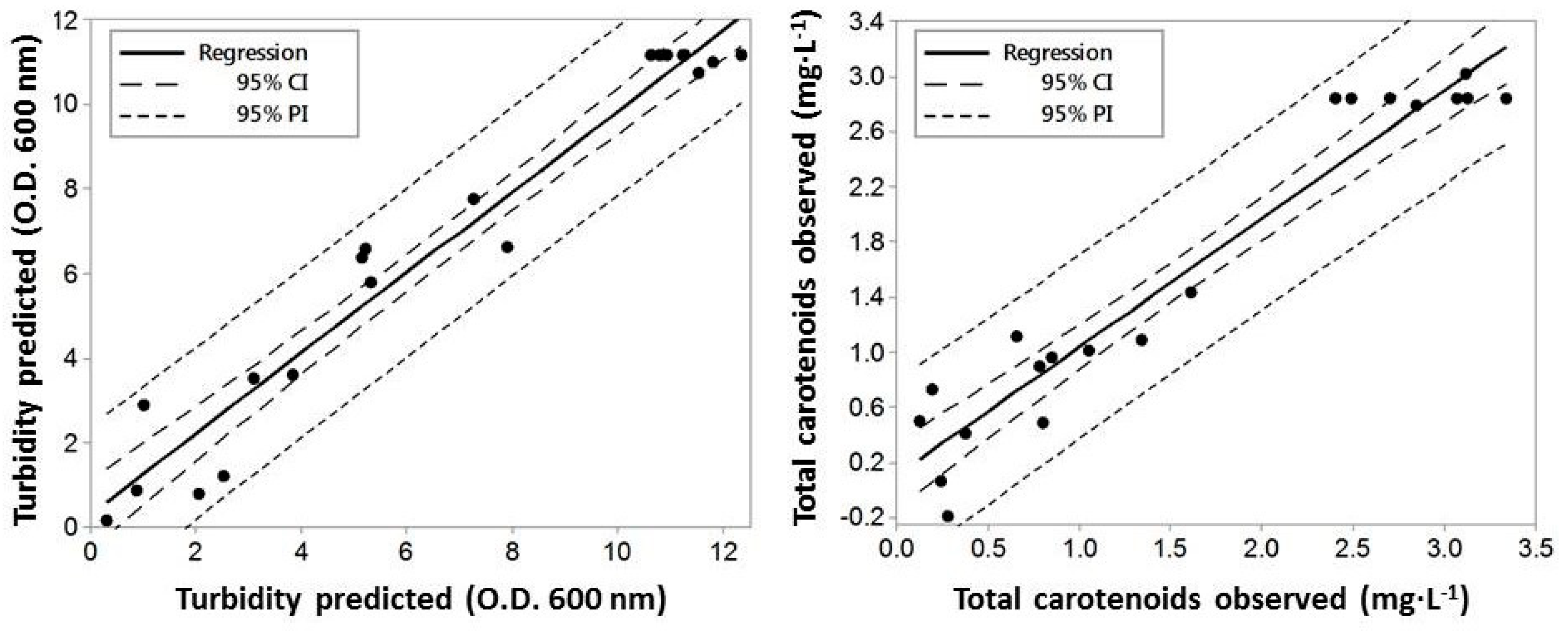
2.3. Validation of the Optimal Conditions for Growth and Total Carotenoid Production by H. mediterranei
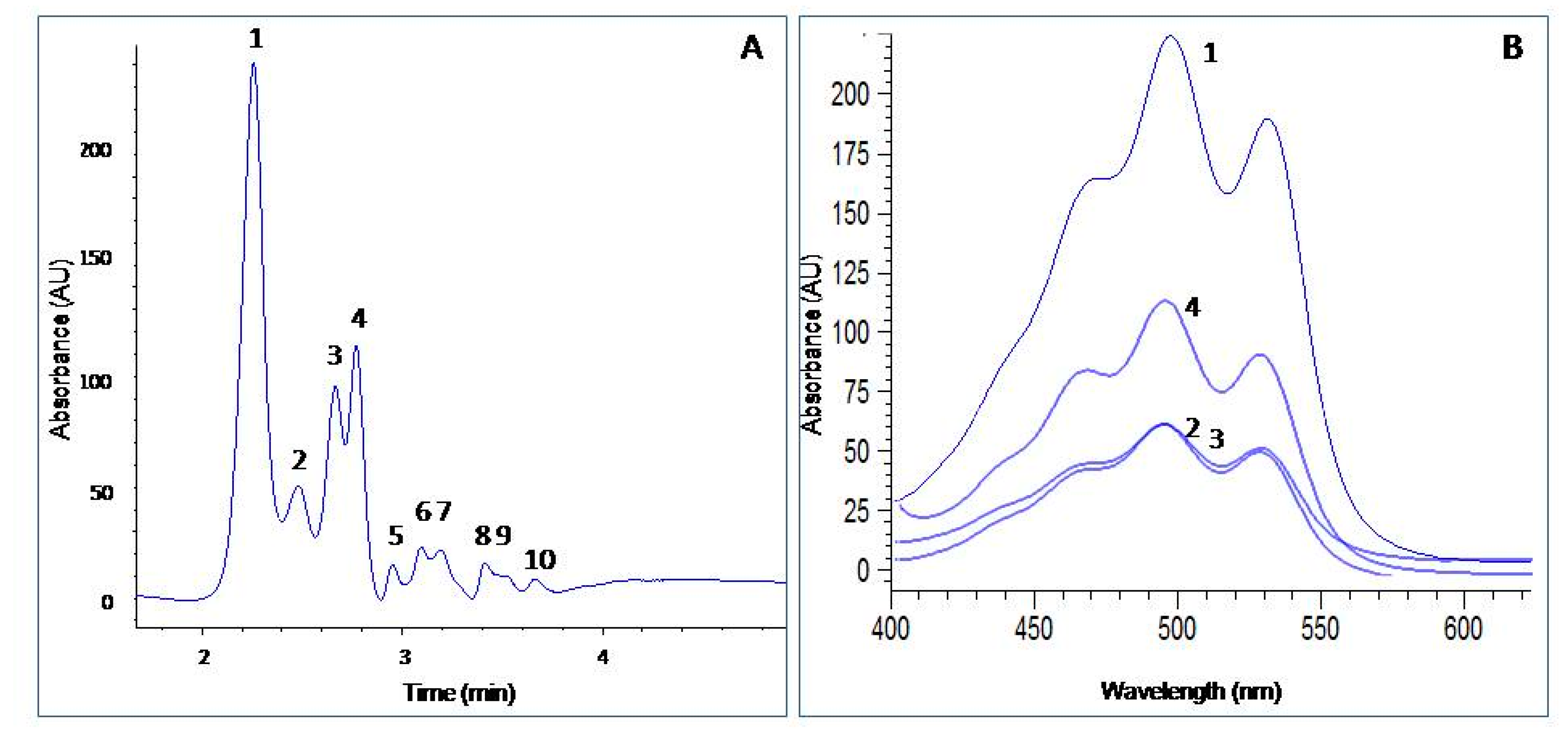
2.4. Carotenoid Profile Obtained from H. mediterranei
2.5. Bacterioruberin Production by H. mediterranei
3. Materials and Methods
3.1. Microorganism
3.2. Growth Conditions and Biomass Quantification
3.3. Extraction, Quantification, and Analysis of Pigments
3.4. Response Surface Methodology Experimental Design
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vílchez, C.; Forján, E.; Cuaresma, M.; Bédmar, F.; Garbayo, I.; Vega, J.M. Marine carotenoids: Biological functions and commercial applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Jaswir, I. Carotenoids: Sources, medicinal properties and their application in food and nutraceutical industry. J. Med. Plants Res. 2011, 5, 7119–7131. [Google Scholar] [CrossRef]
- Torregrosa-Crespo, J.; Montero, Z.; Fuentes, J.L.; García-Galbis, M.R.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Exploring the valuable carotenoids for the large-scale production by marine microorganisms. Mar. Drugs 2018, 16, 203. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Hallsworth, J.E. Microbial weeds in hypersaline habitats: The enigma of the weed-like Haloferax mediterranei. FEMS Microbiol. Lett. 2014, 359, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.J.; Martínez-Espinosa, R.M. Carotenoids from Haloarchaea and their potential in biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseguer, I.; Rodriguez-Valera, F. Effect of Halocin H4 on Cells of Halobacterium halobium. Microbiology 1986, 132, 3061–3068. [Google Scholar] [CrossRef]
- Calegari-Santos, R.; Diogo, R.A.; Fontana, J.D.; Bonfim, T.M.B. Carotenoid Production by Halophilic Archaea Under Different Culture Conditions. Curr. Microbiol. 2016, 72, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.W.; Hsu, S.; Lin, M.T.; Hsu, Y. Mass production of C50 carotenoids by Haloferax mediterranei in using extruded rice bran and starch under optimal conductivity of brined medium. Bioprocess Biosyst. Eng. 2015, 38, 2361–2367. [Google Scholar] [CrossRef] [PubMed]
- Mandelli, F.; Miranda, V.S.; Rodrigues, E.; Mercadante, A.Z. Identification of carotenoids with high antioxidant capacity produced by extremophile microorganisms. World J. Microbiol. Biotechnol. 2012, 28, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Yatsunami, R.; Ando, A.; Yang, Y.; Takaichi, S.; Kohno, M.; Matsumura, Y.; Ikeda, H.; Fukui, T.; Nakasone, K.; Fujita, N.; et al. Identification of carotenoids from the extremely halophilic archaeon Haloarcula japonica. Front. Microbiol. 2014, 5, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Rodriguez-Valera, F. The contribution of halophilic Bacteria to the red coloration of saltern crystallizer ponds. FEMS Microbiol. Ecol. 2001, 36, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Oren, A.; Hirschberg, J.; Mann, V.; Jehlička, J. Effects of nicotine on the biosynthesis of carotenoids in halophilic Archaea (class Halobacteria): An HPLC and Raman spectroscopy study. Extremophiles 2018, 22, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.J.; Ku, K.L.; Lee, M.H.; Su, N.W. Influence of nutritive factors on C50 carotenoids production by Haloferax mediterranei atcc 33500 with two-stage cultivation. Bioresour. Technol. 2010, 101, 6487–6493. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, M. Optimization of Total Carotenoid Production by Halorubrum sp. TBZ126 Using Response Surface Methodology. J. Microb. Biochem. Technol. 2014, 6. [Google Scholar] [CrossRef]
- El-Sayed, W.S.M.; Takaichi, S.; Saida, H.; Kamekura, M.; Abu-Shady, M.; Seki, H.; Kuwabara, T. Effects of light and low oxygen tension on pigment biosynthesis in Halobacterium salinarum, revealed by a novel method to quantify both retinal and carotenoids. Plant Cell Physiol. 2002, 43, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Asker, D.; Ohta, Y. Production of canthaxanthin by extremely halophilic bacteria. J. Biosci. Bioeng. 1999, 88, 617–621. [Google Scholar] [CrossRef]
- De la Vega, M.; Sayago, A.; Ariza, J.; Barneto, A.G.; León, R. Characterization of a bacterioruberin-producing Haloarchaea isolated from the marshlands of the Odiel river in the southwest of Spain. Biotechnol. Prog. 2016, 32, 592–600. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.E.; Altekar, W.; D’Souza, S.F. Adaptive response of Haloferax mediterranei to low concentrations of NaCl (<20%) in the growth medium. Arch. Microbiol. 1997, 168, 68–71. [Google Scholar]
- Schneegurt, M.A. Media and conditions for the growth of halophilic and halotolerant bacteria and archaea. In Advances in Understanding the Biology of Halophilic Microorganisms; Vreeland, R.H., Ed.; Springer: Dordrecht, The Netherlands, 2012; ISBN 9789400755390. [Google Scholar]
- Jehlička, J.; Edwards, H.G.M.; Oren, A. Bacterioruberin and salinixanthin carotenoids of extremely halophilic Archaea and Bacteria: A Raman spectroscopic study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 106, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Calo, P.; de Miguel, T.; Sieiro, C.; Velazquez, J.B.; Villa, T.G. Ketocarotenoids in halobacteria: 3-hydroxy-echinenone and trans-astaxanthin. J. Appl. Bacteriol. 1995, 79, 282–285. [Google Scholar] [CrossRef]
- Sui, L.; Liu, L.; Deng, Y. Characterization of halophilic C50 carotenoid-producing archaea isolated from solar saltworks in Bohai Bay, China. Chin. J. Oceanol. Limnol. 2014, 32, 1280–1287. [Google Scholar] [CrossRef]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological properties of carotenoids extracted from Halobacterium halobium isolated from a Tunisian solar saltern. BMC Complement. Altern. Med. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the extreme halophilic archaeon Haloterrigena turkmenica: Identification and antioxidant activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Short communication isolation of Extremely Halophilic Bacteria Able to Grow in Defined Inorganic Media with Single Carbon Sources. Microbiology 1980, 119, 535–538. [Google Scholar] [CrossRef]
- Herney-Ramirez, J.; Lampinen, M.; Vicente, M.A.; Costa, C.A.; Madeira, L.M. Experimental design to optimize the oxidation of orange II dye solution using a clay-based fenton-like catalyst. Ind. Eng. Chem. Res. 2008, 47, 284–294. [Google Scholar] [CrossRef]
- Song, L.; Qin, J.G.; Su, S.; Xu, J.; Clarke, S.; Shan, Y. Micronutrient requirements for growth and hydrocarbon production in the oil producing green Alga Botryococcus braunii (Chlorophyta). PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Raheem, A.; Wan Azlina, W.A.K.G.; Taufiq Yap, Y.H.; Danquah, M.K.; Harun, R. Optimization of the microalgae Chlorella vulgaris for syngas production using central composite design. RSC Adv. 2015. [Google Scholar] [CrossRef]
- Ferreira, S.L.C.; Bruns, R.E.; da Silva, E.G.P.; dos Santos, W.N.L.; Quintella, C.M.; David, J.M.; de Andrade, J.B.; Breitkreitz, M.C.; Jardim, I.C.S.F.; Neto, B.B. Statistical designs and response surface techniques for the optimization of chromatographic systems. J. Chromatogr. A 2007, 1158, 2–14. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Responses | |||||||
|---|---|---|---|---|---|---|---|---|
| Coded Levels | ||||||||
| Std Order | Temperature (°C) | pH | Salinity (NaCl% w/v) | Turbidity (O.D.600 nm) | Total Carotenoids (mg/L) | |||
| 1 | −1 | 23.8 | −1 | 5.5 | −1 | 9.8 | 2.53 | 0.28 |
| 2 | 1 | 23.8 | −1 | 8.5 | −1 | 9.8 | 3.09 | 1.61 |
| 3 | −1 | 41.3 | 1 | 5.5 | −1 | 9.8 | 7.88 | 0.80 |
| 4 | 1 | 41.3 | 1 | 8.5 | −1 | 9.8 | 11.79 | 3.12 |
| 5 | −1 | 23.8 | −1 | 5.5 | 1 | 23.3 | 0.88 | 1.34 |
| 6 | 1 | 23.8 | −1 | 8.5 | 1 | 23.3 | 5.31 | 1.05 |
| 7 | −1 | 41.3 | 1 | 5.5 | 1 | 23.3 | 2.04 | 0.24 |
| 8 | 1 | 41.3 | 1 | 8.5 | 1 | 23.3 | 7.25 | 0.78 |
| 9 | −1.68 | 32.5 | 0 | 4.5 | 0 | 16.5 | 1.01 | 0.19 |
| 10 | 1.68 | 32.5 | 0 | 9.5 | 0 | 16.5 | 11.51 | 2.85 |
| 11 | 0 | 17.8 | 1.68 | 7.0 | 0 | 16.5 | 0.29 | 0.13 |
| 12 | 0 | 47.2 | 1.68 | 7.0 | 0 | 16.5 | 5.14 | 0.85 |
| 13 | 0 | 32.5 | 0 | 7.0 | −1.68 | 5.1 | 5.22 | 0.66 |
| 14 | 0 | 32.5 | 0 | 7.0 | 1.68 | 27.9 | 3.82 | 0.37 |
| 15 * | 0 | 32.5 | 0 | 7.0 | 0 | 16.5 | 10.93 | 3.34 |
| 16 * | 0 | 32.5 | 0 | 7.0 | 0 | 16.5 | 11.25 | 3.13 |
| 17 * | 0 | 32.5 | 0 | 7.0 | 0 | 16.5 | 10.78 | 2.40 |
| 18 * | 0 | 32.5 | 0 | 7.0 | 0 | 16.5 | 10.62 | 2.71 |
| 19 * | 0 | 32.5 | 0 | 7.0 | 0 | 16.5 | 11.21 | 2.50 |
| 20 * | 0 | 32.5 | 0 | 7.0 | 0 | 16.5 | 12.34 | 3.07 |
| Peak | Carotenoid | Retention Time (min) | λmax (nm) | Molecular Ion (m/z) M+ | Fragments Profile |
|---|---|---|---|---|---|
| 1 | BR | 2.325 | 468, 496, 530 | 740.7 | 723.7, 705.7, 687.7, 666.7, 561.5, 515,1 |
| 2 | BR | 2.553 | 468, 494, 528 | 740.7 | 723.7, 705.7, 681,6, 666.8, 655.6, 627.6 |
| 3 | BR | 2.740 | 468, 496, 528 | 740.7 | 723.7, 705.7, 682.6, 669.6, 665.6 |
| 4 | BR | 2.816 | 464, 494, 524 | 740.7 | 723.7, 705.7, 682.6, 665.6 |
| 5 | MABR | 3.021 | 470, 500, 534 | 737.7 | 725.6, 709.6, 699.7 |
| 6 | BABR | 3.168 | 460, 488, 520 | 705.7 | 681.6, 669.7, 579.7, 522.7 |
| 7 | BABR | 3.233 | 456, 485, 526 | 705.7 | 699.7, 671.7, 668.7, 647.6, 579.6 |
| 8 | BABR | 3.508 | 472, 498, 532 | 705.7 | 699.7, 687.7, 671.7, 653.8, 607.6 |
| 9 | BABR | 3.620 | 468, 490, 520 | 705.7 | 699.7, 671.7, 653.8, 550.6 |
| Independent Variables | Symbols | Levels | ||||
|---|---|---|---|---|---|---|
| −1.68 * | −1 | 0 | 1 | 1.68 * | ||
| Temperature (°C) | X1 | 17.8 | 23.8 | 32.5 | 41.3 | 47.2 |
| pH | X2 | 4.5 | 5.5 | 7.0 | 8.5 | 9.5 |
| [NaCl]% (w/v) | X3 | 5.15 | 9.75 | 16.50 | 23.25 | 27.85 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montero-Lobato, Z.; Ramos-Merchante, A.; Fuentes, J.L.; Sayago, A.; Fernández-Recamales, Á.; Martínez-Espinosa, R.M.; Vega, J.M.; Vílchez, C.; Garbayo, I. Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology. Mar. Drugs 2018, 16, 372. https://doi.org/10.3390/md16100372
Montero-Lobato Z, Ramos-Merchante A, Fuentes JL, Sayago A, Fernández-Recamales Á, Martínez-Espinosa RM, Vega JM, Vílchez C, Garbayo I. Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology. Marine Drugs. 2018; 16(10):372. https://doi.org/10.3390/md16100372
Chicago/Turabian StyleMontero-Lobato, Zaida, Adrián Ramos-Merchante, Juan Luis Fuentes, Ana Sayago, Ángeles Fernández-Recamales, Rosa María Martínez-Espinosa, José María Vega, Carlos Vílchez, and Inés Garbayo. 2018. "Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology" Marine Drugs 16, no. 10: 372. https://doi.org/10.3390/md16100372
APA StyleMontero-Lobato, Z., Ramos-Merchante, A., Fuentes, J. L., Sayago, A., Fernández-Recamales, Á., Martínez-Espinosa, R. M., Vega, J. M., Vílchez, C., & Garbayo, I. (2018). Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology. Marine Drugs, 16(10), 372. https://doi.org/10.3390/md16100372