Sterol and Sphingoid Glycoconjugates from Microalgae



Abstract
:1. Introduction
2. Sterols and Sterol Glycoconjugates. Structural Diversity and Taxonomic Distribution
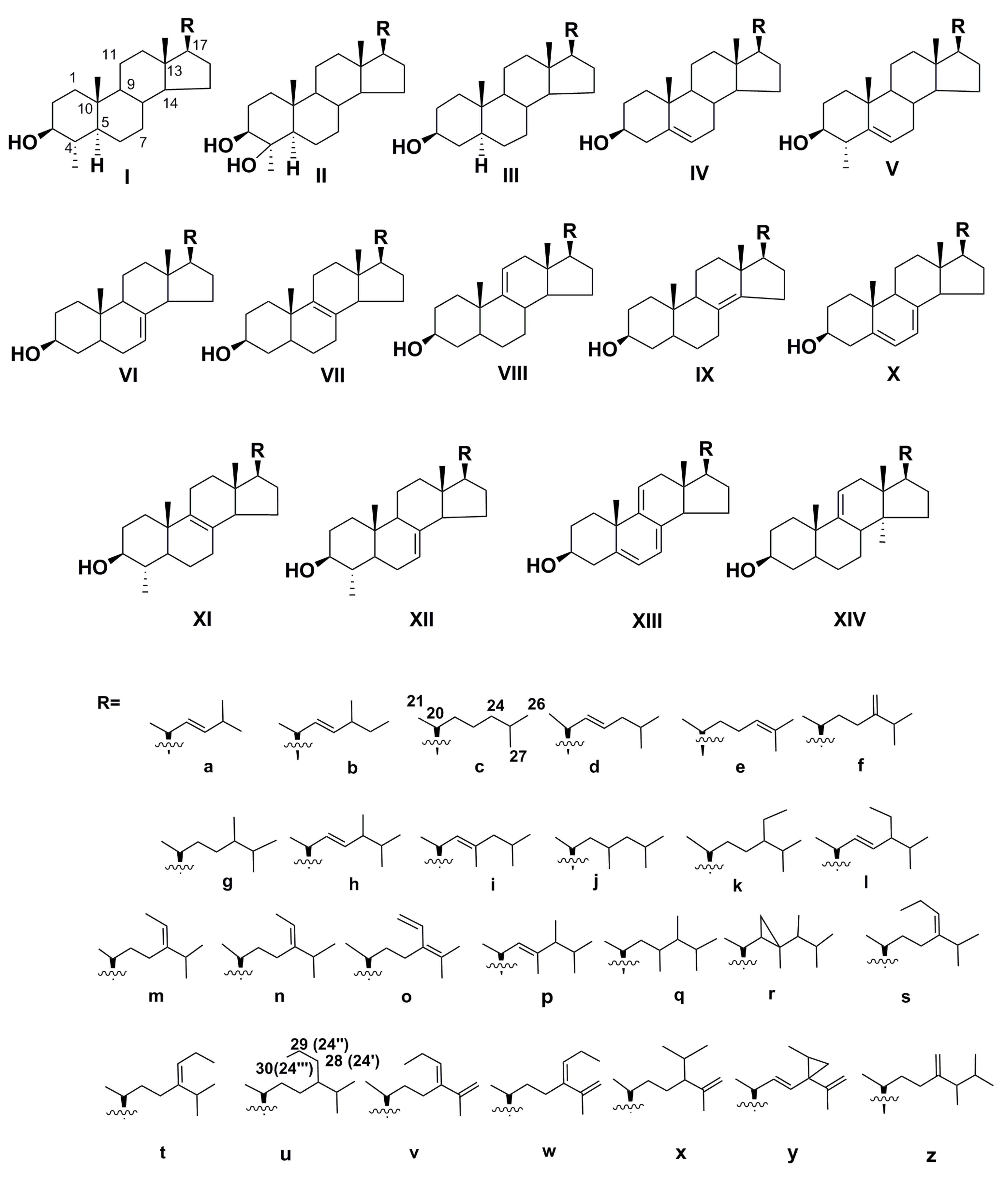
2.1. Free Sterols
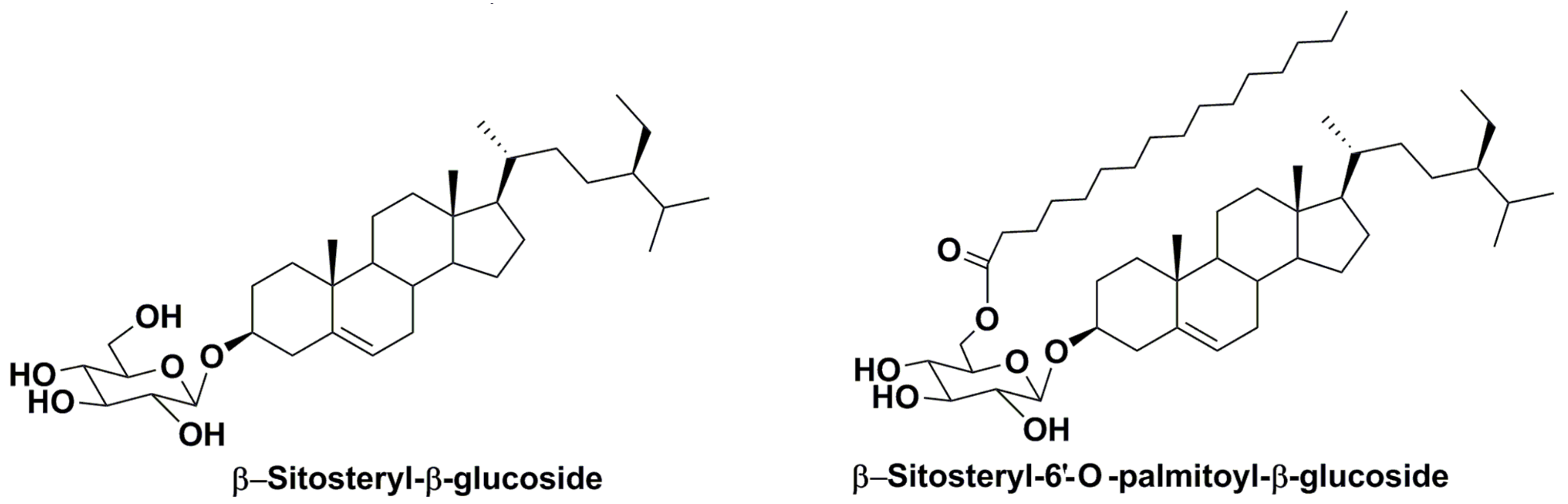
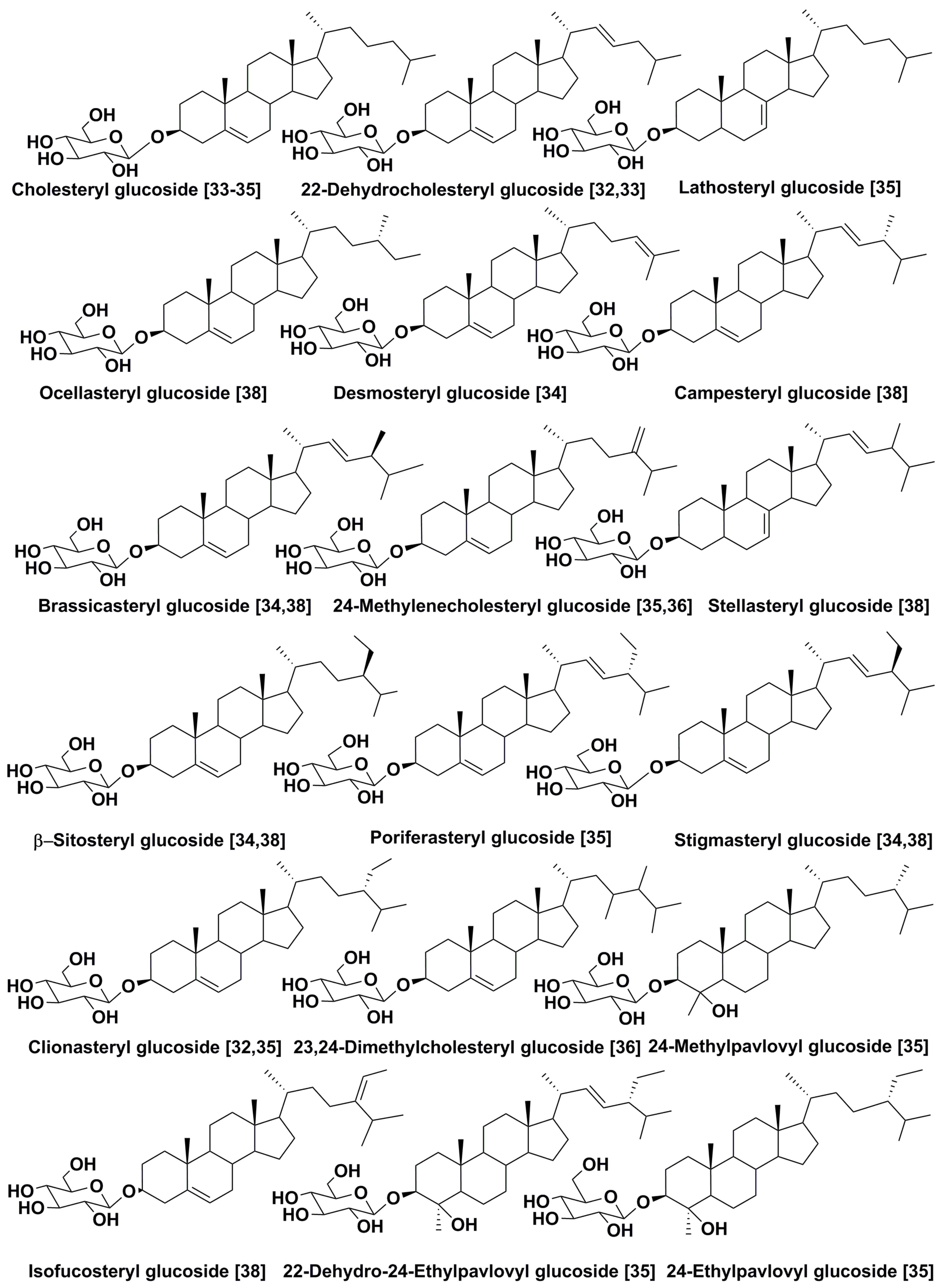
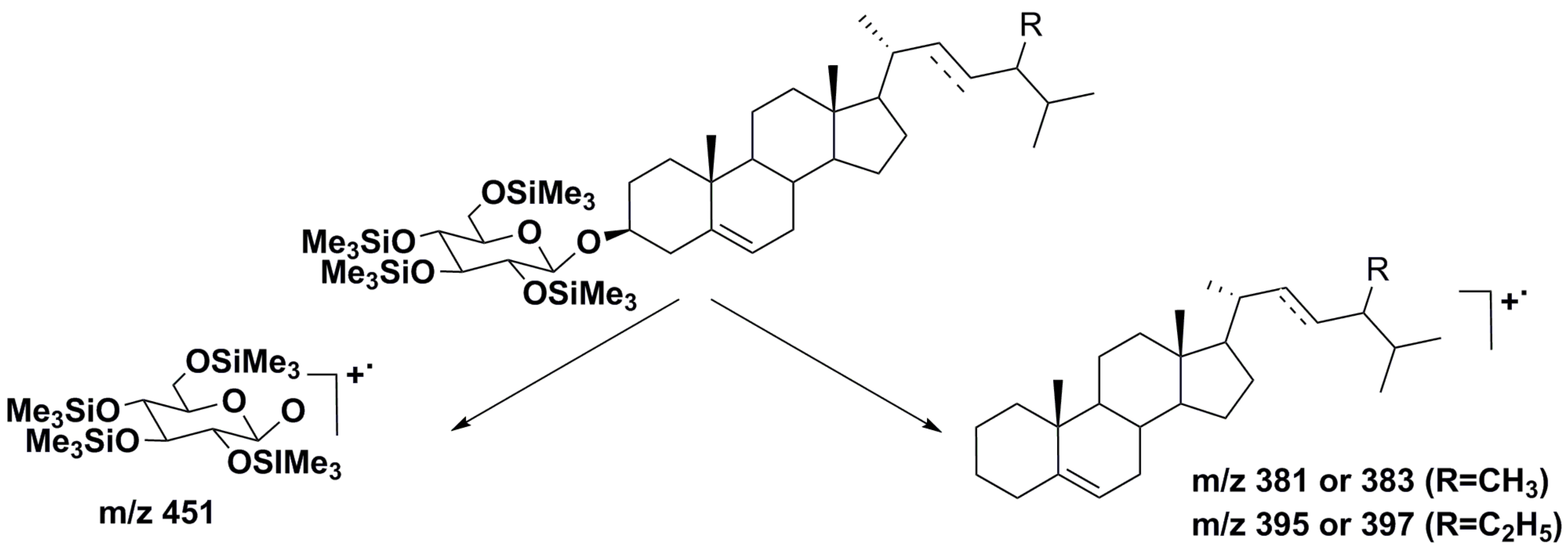
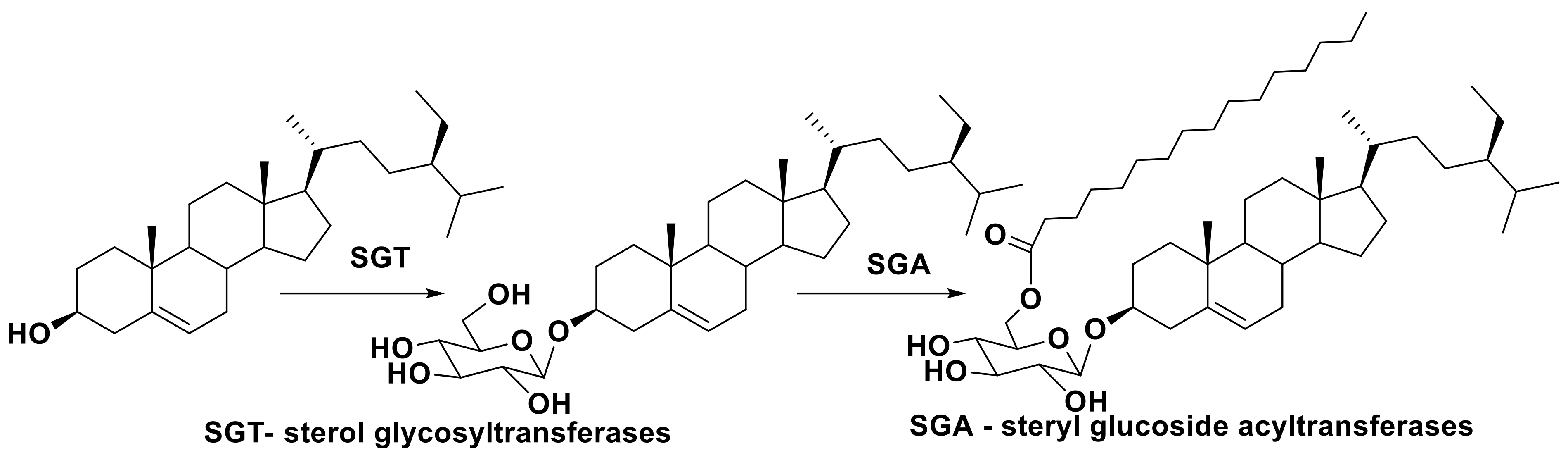
2.2. Structural Diversity and Analysis of Steryl Glycoconjugates
2.3. Biological Activities and Biological Functions
2.4. Biosynthesis of Glycosylated Sterols
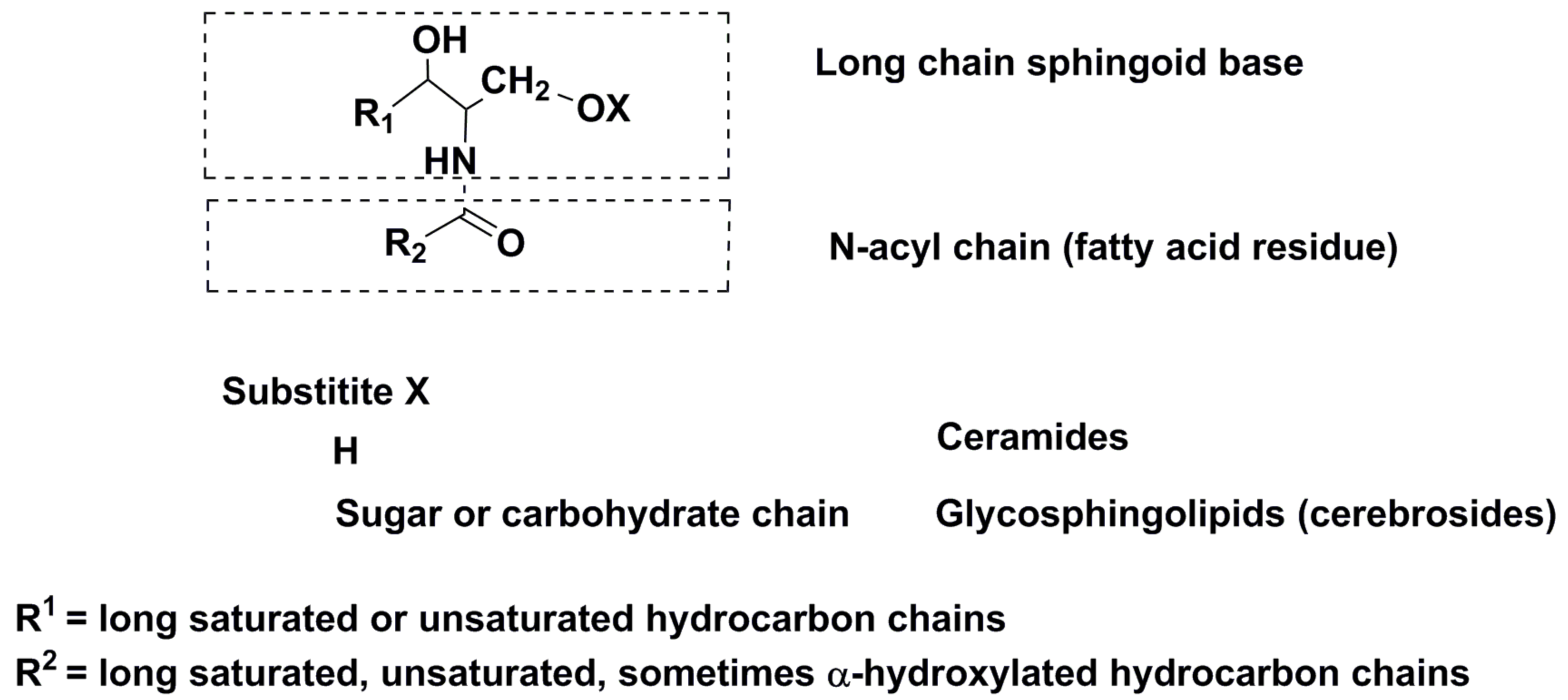
3. Sphingoid Glycoconjugates
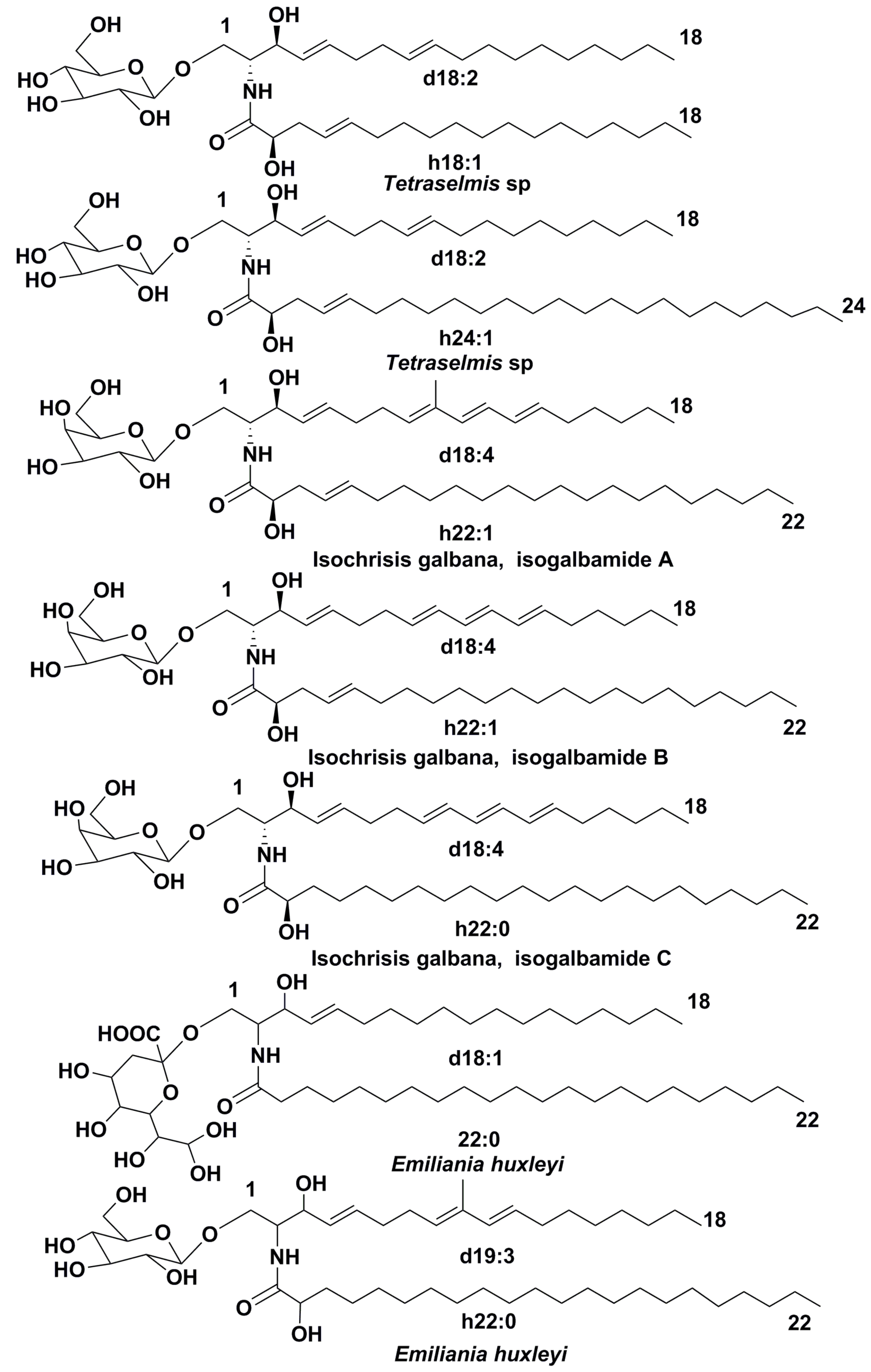
3.1. Structural Diversity
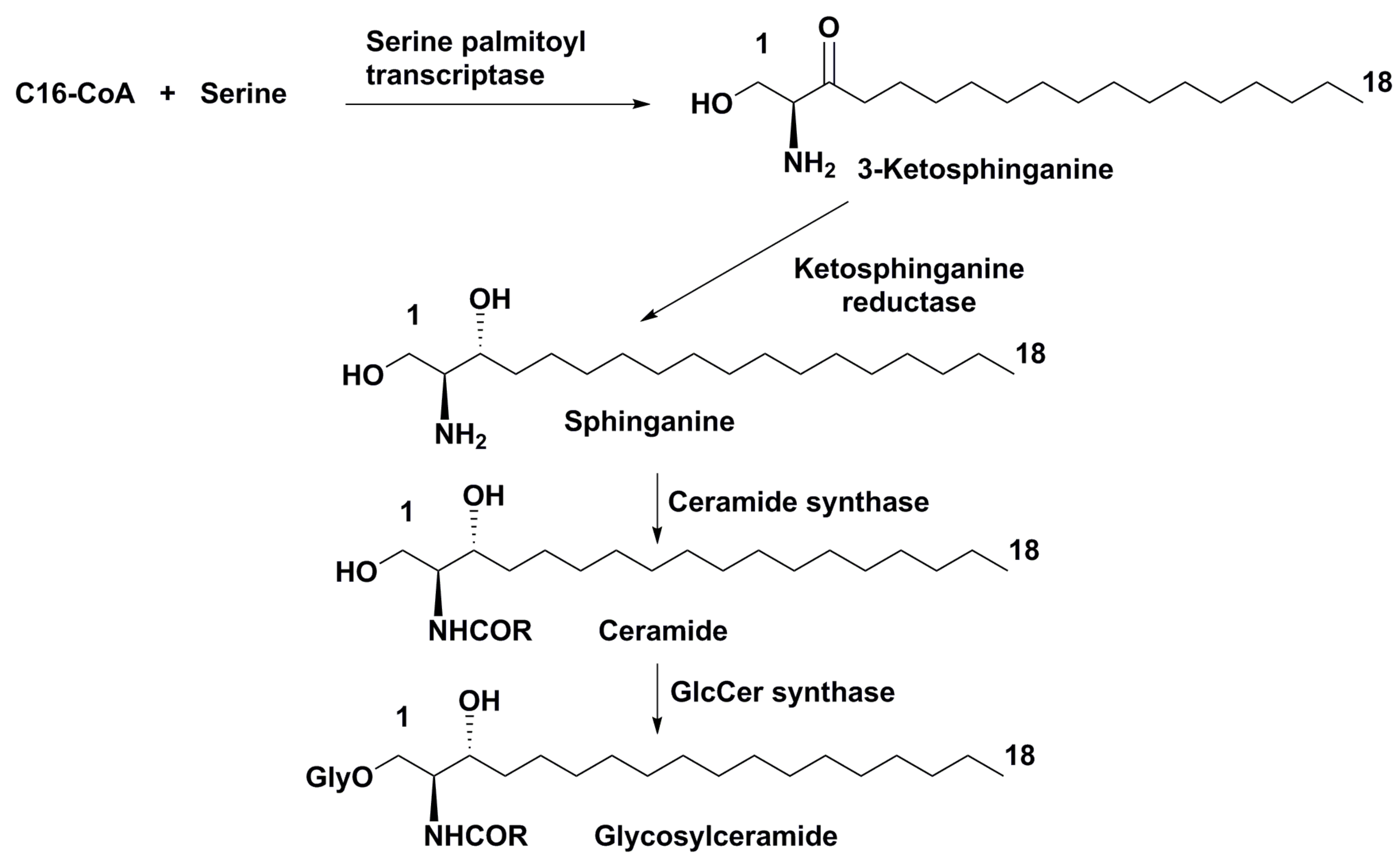
3.2. Biosynthesis, Biological Activities and Biological Roles of Microalgal Glycosphingolipids
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borowitzka, M. Systematics, taxonomy and species names: Do they matter? In The Physiology of Microalgae; Developments in Applied Phycology; Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer International Publishing: Dordrecht, The Netherlands, 2016; pp. 655–681, ISBN 978-3-319-24945-2 or 978-3-319-24943-8. [Google Scholar]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E. Phycology, 4th ed.; Cambridge University Press: New York, NY, USA, 2008; ISBN 978-0-511-38669-5. [Google Scholar]
- Stonik, V.; Stonik, I. Low-molecular weight metabolites from diatoms: Structures, biological roles and biosynthesis. Mar. Drugs 2015, 13, 3672–3709. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, J.; Škaloud, P. Chrysophyta. In Handbook of the Protists; Archibald, J., Simpson, A., Slamovits, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 331–366. ISBN 978-3-319-28147-6. [Google Scholar]
- Steindinger, K.A.; Jangen, K. Dinoflagellates. In Identifying Marine Diatoms and Dinoflagellates; Thomas, C.R., Ed.; Academic Press: San-Diego, CA, USA, 1997; pp. 387–584. ISBN 0-12-693018-X. [Google Scholar]
- Hasle, G.; Syvertsen, E. Diatoms. In Identifying Marine Diatoms and Dinoflagellates; Thomas, C.R., Ed.; Academic Press: San-Diego, CA, USA, 1997; pp. 5–385. ISBN 0-12-693018-X. [Google Scholar]
- Sirakov, I.; Velichkova, K.; Stoyanova, S.; Staykov, Y. The importance of microalgae for aquaculture industry. Review. Int. J. Fish. Aquat. Stud. 2015, 2, 81–84. [Google Scholar]
- Volkman, J.K. A review of sterol markers for marine and terrigenous organic matter. Org. Geochem. 1986, 9, 83–99. [Google Scholar] [CrossRef]
- Volkman, J.K. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Rampen, S.W.; Abbas, B.A.; Shouten, S.; Sinninghe Dumste, J.J.S. A comprehensive study of sterols in marine diatoms (Bacillariophyta): Implications for their use as tracers for diatom productivity. Limnol. Oceanogr. 2010, 55, 91–105. [Google Scholar] [CrossRef]
- Volkman, J.K. Sterols and other triterpenoids: Source specificity and evolution of biosynthetic pathways. Org. Geochem. 2005, 36, 139–159. [Google Scholar] [CrossRef]
- Volkman, J.K. Sterols in microalgae. In The Physiology of Microalgae; Developments in Applied Phycology, Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer International Publishing: Dordrecht, The Netherlands, 2016; pp. 485–505, ISBN 978-3-319-24945-2 or 978-3-319-24943-8. [Google Scholar]
- Stonik, I.V.; Kapustina, I.I.; Aizdaicher, N.A.; Svetashev, V.I. Sterols and fatty acids from Attheya planktonic diatoms. Chem. Nat. Compd. 2017, 53, 422–425. [Google Scholar] [CrossRef]
- Giner, J.L.; Wikfors, G.H. “Dinoflagellate sterols” in marine diatoms. Phytochemistry 2011, 72, 1896–1901. [Google Scholar] [CrossRef]
- Rampen, S.W.; Volkman, J.K.; Hur, S.B.; Abbas, D.A.; Shouten, S.; Jameson, I.D.; Holdsworth, D.G.; Bae, J.H.; Sinninghe Dumste, J.J.S. Occurrence of gorgosterol in diatoms of the genus Delphineis. Org. Geochem. 2009, 40, 144–147. [Google Scholar] [CrossRef]
- Giner, L.L.; Zhao, H.; Boyer, G.L.; Satchell, M.F.; Anderson, R.A. Sterol chemotaxonomy of marine Pelagophyte algae. Chem. Biodivers. 2009, 6, 1111–1130. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, G.A.; Brown, M.R.; Volkman, J.K. Cryptophyceae and Raphidophyceae; chemotaxonomy, phylogeny, and application. Phytochemistry 2005, 66, 2557–2570. [Google Scholar] [CrossRef] [PubMed]
- Leblond, J.D.; Lasiter, A.D.; Li, C.; Logares, R.; Rengefors, K.; Evens, T.J. A data mining approach to dinoflagellate clustering according to sterol composition: Correlations with evolutionary history. Int. J. Data Min. Bioinform. 2010, 4, 431–451. [Google Scholar] [CrossRef] [PubMed]
- Giner, J.L.; Ceballos, H.; Tang, Y.Z.; Gobler, C.J. Sterols and fatty acids of the harmful dinoflagellate Cochlodinium polykrikoides. Chem. Biodivers. 2016, 13, 249–252. [Google Scholar] [CrossRef]
- Volkman, J.K.; Kearney, P.; Jeffrey, S.W. A new source of 4-methyl sterols and 5a(H)-stanols in sediments: Premnesiophyte microalgae of the genus Pavlova. Org. Geochem. 1990, 15, 489–497. [Google Scholar] [CrossRef]
- Volkman, J.K.; Farmer, C.L.; Barrent, S.M.; Sikes, E.L. Unusual dihydroxysterols as chemotaxonomic markers for microalgae from the order Pavlovales (Haptophyceae). J. Phycol. 1997, 33, 1016–1023. [Google Scholar] [CrossRef]
- Leblond, J.D.; Timofte, H.I.; Roche, S.A.; Porter, N.M. Sterols of glaucocystophytes. Phycol. Res. 2011, 59, 129–134. [Google Scholar] [CrossRef]
- Leblond, J.D.; Dahmen, J.L.; Seipelt, R.L.; Elrod-Erickson, M.J.; Kincaid, R.; Howard, J.C.; Evans, T.J.; Chapman, P.J. Lipid composition of chlorarachniophytes (Chlorarachniophyceae) from genera Bigelowiella, Gymnochlora, and Lotharella. J. Phycol. 2005, 41, 311–321. [Google Scholar] [CrossRef]
- Bilbao, P.G.S.; Damani, C.; Salvador, G.A.; Leonardi, P. Haematococcus pluvalis as a source of fatty acids and phytosterols potential nutritional and biological applications. J. Appl. Phycol. 2016, 28, 3283–3294. [Google Scholar] [CrossRef]
- Martin-Creuzberg, D.; Merkel, P. Sterols in freshwater microalgae: Potential applications for zooplankton nutrition. J. Plankt. Res. 2016, 38, 865–877. [Google Scholar] [CrossRef]
- Gladu, P.K.; Paterson, G.W.; Wikfors, G.W.; Chitwood, D.J.; Lusby, W.R. The occurrence of brassicasterol and epibrassicasterol in the Chromophycota. Comp. Biochem. Physiol. 1990, 97B, 491–494. [Google Scholar] [CrossRef]
- Lewin, R.A. Biochemical taxonomy. In Algal Physiology and Biochemistry (Botanical Monographs); Stewart, W.D.P., Ed.; Blackwell Sci.: Oxford, UK, 1974; pp. 1–32. ISBN 978-0632091003. [Google Scholar]
- Wang, G.; Wang, T. Characterization of lipid components in two microalgae for biofuel application. J. Am. Oil Chem. Soc. 2011, 89, 135–143. [Google Scholar] [CrossRef]
- Yao, L.; Gerde, J.A.; Lee, S.-L.; Wang, T.; Harrata, K.A. Microalgae lipid characreization. J. Agric. Food Chem. 2015, 63, 1773–1787. [Google Scholar] [CrossRef] [PubMed]
- Duperon, R.M.; Thiersault, M.; Duperon, P. Occurrence of steryl glycosides and acylated sreyl glycosides in some marine algae. Phytochemistry 1983, 22, 535–538. [Google Scholar] [CrossRef]
- Nystrӧm, L.; Schär, A.; Lampi, A.M. Steryl glycosides and acetylated steryl glycosides in plant foods reflect unique sterol patterns. Eur. J. Lipid Sci. Technol. 2012, 114, 656–669. [Google Scholar] [CrossRef]
- Mohammady, N.G. Total, free and conjugated sterolic forms in three microalgae used in mariculture. Z. Naturforsch. C 2004, 59, 9–10. [Google Scholar] [CrossRef]
- Vernon, B.; Dauguet, J.-C.; Billard, C. Sterolic biomarkers in marine phytoplankton. I. Free and conjugated sterols of Pavlova lutheri (Haptophyta). Eur. J. Phycol. 1996, 31, 211–215. [Google Scholar] [CrossRef]
- Vernon, B.; Dauguet, J.-C.; Billard, C. Sterolic biomarkers in marine phytoplankton. I. Free and conjugated sterols of seven species used in mariculture (Haptophyta). Eur. J. Phycol. 1998, 34, 273–279. [Google Scholar] [CrossRef]
- Veron, B.; Billard, C.; Dauguet, J.C.; Hartmann, M.-A. Sterol composition of Phaeodactylum tricornutum as influenced by growth temperature and light spectral quality. Lipids 1996, 31, 989–994. [Google Scholar] [CrossRef]
- Kovganko, N.V.; Kashkan, Z.N. Sterol glycosides and acylglycosides. Chem. Nat. Compd. 1999, 35, 479–497. [Google Scholar] [CrossRef]
- Phillips, K.M.; Ruggio, D.M.; Ashraf-Khorassani, M. Analysis of steryl glucosides in foods and dietary supplement by solid-phase extraction and gas chromatography. J. Food Lipids 2005, 12, 124–140. [Google Scholar] [CrossRef]
- Pieber, B.; Schnober, S.; Goebl, C.; Mittelbach, M. Novel sensitive determination of steryl glycosides in biodiesel by gas chromatography—Mass spectrometry. J. Chromatogr. A 2010, 1217, 6555–6561. [Google Scholar] [CrossRef]
- Oppliger, S.R.; Mungen, L.H.; Nystrom, L. Rapid and highly accurate detection of steryl glycosides by ultraperformance liquid chromatography-quadropole time-of-flight mass spectrometry (UPLC-Q-TOF MS). J. Agric. Food Chem. 2014, 62, 9410–9419. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhang, Y.; Ran, Y.; Lai, W.; Ran, Z.; Xu, J.; Zhou, C.; Yan, X. Characterization of steryl glycosides in marine microalgae by gas chromatography-triple guadropole mass spectrometry (GC-QQQ-MS). J. Sci. Food Agric. 2018, 98, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Bouic, P.J. The role of phytosterols and phytosterolines in immune modulation: A review of the past 10 years. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, M. Immunological functions of steryl glycosides. Arch. Immunol. Ther. Exp. (Warsz) 2012, 60, 351–359. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.Y.; Park, J.H.; Jung, H.S.; Kim, J.S.; Kang, S.S.; Kim, Y.S.; Han, Y. Immunoregulatory activity by daucosterol, a beta-sistosterol glycoside, induces protective Th1 immune response against disseminated Candidiasis in mice. Vaccine 2007, 25, 3834–3840. [Google Scholar] [CrossRef]
- Lin, X.; Ma, L.; Racette, S.B.; Anderson Spearie, C.L.; Ostlund, R.E.J. Phytosterol glycosides reduce cholesterol absorption in humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G931–G935. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Ma, L.; Moreau, R.A.; Ostlund, R.E.J. Glycosidic bond cleavage is not required for phyrosteryl glucoside-induced reduction of cholesterol absorption in mice. Lipids 2011, 46, 701–708. [Google Scholar] [CrossRef]
- Kent, M.; Welladsen, H.M.; Mangott, A.; Li, Y. Nutritional evaluation of Australian microalgae as potential human health supplements. PLoS ONE 2015, 10, e0118985. [Google Scholar] [CrossRef]
- Bouic, P.J.; Lamprecht, J.H. Plant sterols and sterolins: A review of their immune-modulating properties. Altern. Med. Rev. 1999, 4, 170–177. [Google Scholar] [PubMed]
- Meireless, L.A.; Guedas, A.C.; Malcata, F.X. Lipid class composition of the microalga Pavlova lutheri: Eicosapentaenic and docosahexaenoic acids. J. Agric. Food Chem. 2003, 51, 2237–2241. [Google Scholar] [CrossRef] [PubMed]
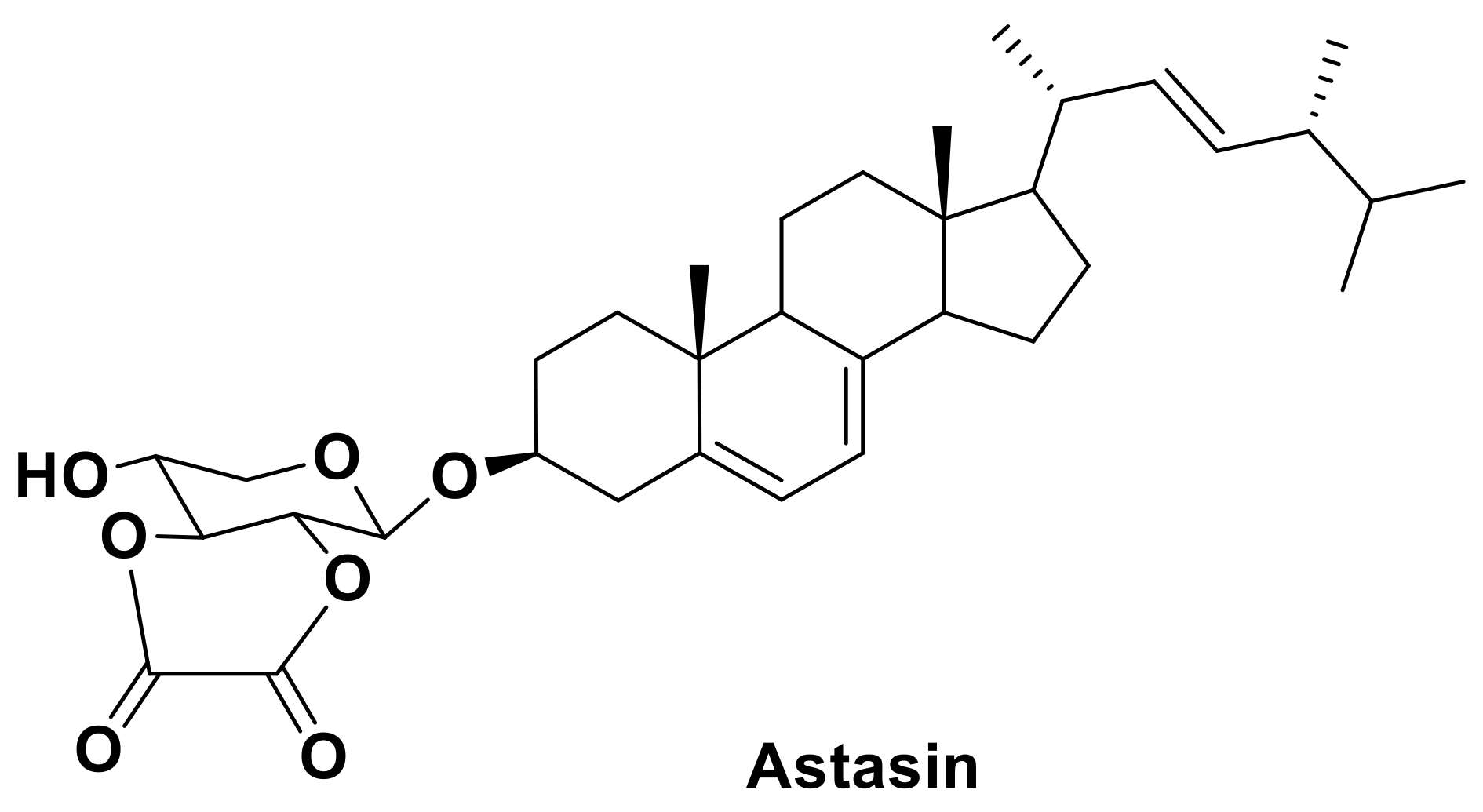
- Kaya, K.; Sano, T.; Shiraishi, F. Astasin, a novel cytotoxic carbohydrate–conjugated ergosterol from the colorless euglenoid, Astasia longa. Biochim. Biophys. Acta 1995, 1255, 201–204. [Google Scholar] [CrossRef]
- Halling, K.K.; Ramstedt, B.; Slotte, J.P. Glycosylation induces shifts in the lateral distribution of cholesterol from ordered towards less ordered domains. Biochim. Biophys. Acta 2008, 1778, 1100–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grille, S.; Zaslawski, A.; Thiele, S.; Plat, J.; Warnecke, D. The functions of steryl glycosides come to those who wait: Recent advances in plants, fungi, bacteria and animals. Prog. Lipid Res. 2010, 49, 262–288. [Google Scholar] [CrossRef]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant sterols: Diversity, biosynthesis, and physiological functions. Biochemistry (Mosc.) 2016, 81, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Kawagoe, Y.; Hogan, P.; Delmer, D. Sitosterol-beta-glucoside as primer for cellulose synthesis in plants. Science 2002, 5552, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Hopp, H.; Romero, P.A.; Daleo, G.R.; Pont Lezica, R. Steryl glycoside biosynthesis in the alga Prototeheca zopfii. Phytochemistry 1978, 17, 1049–1052. [Google Scholar] [CrossRef]
- Wojciechowski, Z.A. Biochermistry of phytosterol conjugates. In Physiology and Biochemistry of Sterols; Patterson, G.W., Ed.; AOCS Publishing: New York, NY, USA, 1992; pp. 361–395. ISBN 978-1439821831. [Google Scholar]
- Chaturvedi, P.; Misra, P.; Tuli, R. Sterol glycosyltransferases—The enzymes that modify sterols. Appl. Biochem. Biotechnol. 2011, 165, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Ullman, P.; Ury, A.; Rimmele, D.; Benveniste, P.; Bouvier-Nave, P. UDP-glucose sterol β-D- glucosyltransferase, a plasma membrane-bound enzyme of plants: Enzymatic properties and lipid dependence. Biochimie 1993, 75, 713–723. [Google Scholar] [CrossRef]
- Yoon, K.; Han, D.; Sommerfield, M.; Hu, Q. Phospholipid:diacylglycerol acyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation while synthesizing triacylglycerol in the unicellular green microalga Chlamydomonas reinhardtii. Plant Cell 2012, 24, 3708–3724. [Google Scholar] [CrossRef]
- Sonnino, S.; Prinetti, A. Membrane domains and the “lipid raft” concept. Curr. Med. Chem. 2013, 20, 4–21. [Google Scholar] [CrossRef]
- Merril, A.H.J.; Wang, M.D.; Park, M.; Sullards, M.C. (Glyco)sphingolipidology: An amazing challenge and opportunity for systems biology. Trends Biochem. Sci. 2007, 32, 457–467. [Google Scholar] [CrossRef]
- Davy, S.K.; Allemand, D.; Weis, V.M. Cell biology of cnidarian-dinoflagellate symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76, 229–261. [Google Scholar] [CrossRef]
- Arakaki, A.; Iwama, D.; Liang, D.; Murakami, N.; Ishikura, M.; Tanaka, T.; Matsunaga, T. Glycosylceramides from marine microalga Tetraselmis sp. Phytochemistry 2013, 85, 107–114. [Google Scholar] [CrossRef]
- De los Reyes, C.; Ortega, M.J.; Rodriguez-Luna, A.; Talero, E.; Motilva, V.; Zubía, E. Molecular characterization and anti-onflammatory activity of galactosylglycerides and galactosylceramides from the microalga Isochrysis galbana. J. Argric. Food Chem. 2016, 64, 8783–8794. [Google Scholar] [CrossRef]
- Fulton, J.M.; Fredericks, H.F.; Bidle, K.D.; Vardi, A.; DiTullio, R.; Van Mooy, A.D. Novel molecular determinants of viral susceptibility and resistance in the lipidome of Emiliania huxleyi. Environ. Microbiol. 2014, 16, 1137–1149. [Google Scholar] [CrossRef]
- Zhao, F.; Xu, J.; Chen, J.; Yan, X.; Zhou, C.; Li, S.; Xu, X.; Ye, F. Sructural elucidation of two types of novel glycosphingolipids in three strains of Skeletonema by liquid chromatography coupled with mass spectrometry. Rapid. Commun. Mass Spectrom. 2013, 27, 1535–1547. [Google Scholar] [CrossRef]
- Yan, X.J.; Chen, D.Y.; Xu, J.L.; Zhou, C.X. Profiles of photosynthetic glycerolipids in three strains of Skeletonema determined by UPLC-Q-TOF-MS. J. Appl. Phycol. 2010, 23, 271–282. [Google Scholar] [CrossRef]
- Li, Y.; Lou, Y.; Mu, T.; Ke, A.; Ran, Z.; Xu, J.; Chen, J.; Zhou, C.; Yan, Q.; Xu, Q.; et al. Sphingoid in marine microalgae: Development and application of a mass spectrometric method for global structural characterization of ceramides in three major phyla. Anal. Chim. Acta 2017, 986, 82–94. [Google Scholar] [CrossRef]
- Hunter, J.T.; Brandsma, J.; Dymond, M.K.; Koster, G.; Moore, C.M.; Postle, A.D.; Mills, R.A.; Attard, G.S. Lipidomics of Thalassiosira pseudonana under phosphorus stress reveal underlying phospholipid substitution dynamics and novel diglycosylceramide substitutes. Appl. Environ. Microbiol. 2018. [Google Scholar] [CrossRef]
- Sperling, P.; Henz, E. Plant sphingolipids: Structural diversity, biosynthesis, first genes and functions. Biochim. Biophys. Acta 2003, 1632, 1–15. [Google Scholar] [CrossRef]
- Lynch, D.V.; Steponkus, P.L. Plasma membrane lipid alterations associated with cold acclimation of winter rye seedlings (Secale cereale L. cv Puma). Plant Physiol. 1987, 83, 761–767. [Google Scholar] [CrossRef]
- Warntcke, D.; Heinz, E. Recently discovered functions of glycosylceramides in plants and fungi. Cell. Mol. Life Sci. 2003, 60, 919–941. [Google Scholar] [CrossRef]
- Abul-Milh, M.; Foster, D.B.; Lingwood, C.A. In vitro binding of Helicobacter pylori to monohexosylceramides. Glycoconj. J. 2001, 18, 253–260. [Google Scholar] [CrossRef]
- Wilson, W.H.; Schroeder, D.C.; Allen, M.J.; Holden, M.T.G.; Parkhill, J.; Barell, B.G.; Churcher, C.; Harnlin, N.; Mungall, K.; Norbertzak, H.; et al. Complete genome sequence and lytic phase transcription profile of a Coccolithovirus. Science 2005, 309, 1090–1092. [Google Scholar] [CrossRef]
- Vardi, A.; Van Mooy, B.A.S.; Fredericks, H.F.; Ropendorf, K.J.; Ossolinski, J.E.; Yaramaty, L.; Bidle, K.D. Viral glycosphingolipids induce lytic infection and cell death in marine phytoplankton. Science 2009, 326, 861–865. [Google Scholar] [CrossRef]
- Ziv, C.; Malitzky, S.; Othman, A.; Ben-Dor, S.; Wei, Y.; Zheng, S.; Aharoni, A.; Hornemann, T.; Vardi, A. Viral serine palmitoyltransferase induces metabolic switch in sphingolipid biosynthesis and is required for infection of a marine alga. Proc. Natl. Acad. Sci. USA 2016, 113, 1907–1916. [Google Scholar] [CrossRef]
- Vardi, A.; Haramaty, L.; Van Mooy, B.A.S.; Fredericks, H.F.; Kimmance, S.A.; Larsen, A.; Bidle, K.D. Host-virus dynamics and subcellular control of cell fate in a natural coccolithophore population. Proc. Natl. Acad. Sci. USA 2012, 109, 19327–19332. [Google Scholar] [CrossRef]
- Gallo, C.; d’Ippolito, G.; Nuzzo, G.; Sardo, A.; Fontana, A. Autoinhibitory sterol sulfates mediate programmed cell death in a bloom-forming marine diatom. Nat. Commun. 2017, 8, 1292. [Google Scholar] [CrossRef] [Green Version]
- Lynch, D.V.; Criss, A.K.; Lehoczky, J.L.; Bui, V.T. Ceramide glucosylation in bean hypocotyl microsome: Evidence that steryl glucoside serves as glucose donor. Arch. Biochem. Biophys. 1997, 340, 3111–3316. [Google Scholar] [CrossRef]
- Lynch, D.V.; Dunn, T.V. An introduction to plant sphingolipids and a review of recent advances in understanding their metabolism and function. New Phytolog. 2004, 161, 677–702. [Google Scholar] [CrossRef] [Green Version]
- Stonik, V.A.; Ponomarenko, L.P.; Makarieva, T.N.; Boguslavsky, V.M.; Dmitrenok, A.S.; Fedrov, S.N.; Strobikin, S.A. Free sterol compositions from the sea cucumbers Pseudostichopus trachus, Holothuria (Microtele) nobilis, Trochostoma orientale and Bathyplotes natans. Comp. Biochem. Physiol. 1998, 120B, 337–347. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Stonik, V.A. Steroid glycosides from marine organisms. Steroids 2011, 76, 425–454. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Sterols |
| Ochrophyta | |
| Bacillariophyceae | C28Δ5,24(28) (IVf), 24R-C28Δ5,22 (epibrassicasterol IVh) [10,11,12,13], C27Δ5 (IVc), 27-nor-C28Δ5,22 (ocellasterol IVb), C29Δ5,22 (IVl [11,13], C29Δ5 (IVk) [14], previously unknown C28Δ7,22 (VIi), C27Δ5,22 (IVd) [13], C29Δ22 (IIIp), C30Δ22 (dinosterol Ip), cyclopropane sterols (Ir, gorgosterol IVr [15,16], C28Δ7,22 (VIh), C28Δ8(9) (VIIg), C29Δ24,28 (IVo and IIIo) [13]. |
| Eustigmatophyceae | C28Δ5 (IVg), C29Δ5,24(28) (IVn), C29Δ5,24(28) (IVm), C29Δ5 (IVk), C28Δ5,24(28) (IVf), C27Δ5 (IVc) [12,13]. |
| Pelagophyceae | C30Δ5 (IVu), C30Δ5,24(28) (IVs,t), rare C30Δ5,24(28),25(26) (IVv,w), trace C30 sterols (IVx-z) [13,17] |
| Chrysophyceae | 24S- and 24R-C29Δ5,22 (IVl), 24S-C28Δ5,22 (IVh), C28Δ5,7,22 (Xh), 24R- and 24S-C29Δ5 (IVk), C29Δ5,7,22 (Xl), C29Δ5,24(28) (IVm,n), C29Δ7,24(28) (Xm), |
| Synurophyceae | C27Δ5 (IVc), C29Δ5 (IVk) [13] |
| Chrysomerophyceae | C29Δ5 (IVk), C28Δ5,24(28) (IVf), C27Δ5 (IVc), C29Δ5,22 (IVl), C29Δ5,24(28) (IVn), C28Δ5,22 (IVh) [13] |
| Xanthophyceae | C27Δ5 (IVc), C29Δ5 (IVk) [13] |
| Dictyophyceae | C28Δ5,24(28) (IVf), C28Δ5,22 (Vd) [13] |
| Rhaphidophyceae | 24S- and 24R-C29Δ5 (IVk), C27Δ5 (IVc), C27Δ8(9) (VIIc), C29Δ5,22 (IVl), 27-nor-C27Δ5,22 (IVb), C29Δ0 (IIIk) [13,18] |
| Dinophyta | |
| Dinophyceae | C30Δ22 (Ip dinosterol), C29Δ0 (Iq), C29Δ0 (1g), C29Δ24(28)(If), C28Δ22 (IIIh), C29Δ22 (IIIp), C28Δ8(14) (IXc), C28Δ8(14),24(28) (amphisterol IXf), C28Δ8(14),22 (IXh), C27Δ8(14),22 (IXb), C28Δ8(14),22 (IXi) and others [13,19,20] |
| Cryptophyta | |
| Cryptophyceae | C28Δ5,22 (IVh), C27Δ5 (IVc), C29Δ5,22 (IVl) [13,18] |
| Haptophyta | |
| Coccolithophyceae | C28Δ5,22 (epibrassicasterol IVh), C29Δ5,22 (stigmasterol IVl), C27Δ5 (IVc), C28Δ5,24(28) (IVf) [13] |
| Pavlovophyceae | C29Δ5,22 (IVl), C27Δ5 (IVc), C29Δ22 (IIIl), C30Δ22 (Il), pavlovols IIg, IIk,IId, minor Ig,h [13,21,22] |
| Euglenophyta | |
| Euglenophyceae | C28Δ5,7,22 (ergosterol Xh), C29Δ8(9) (XIg), C28Δ5,7,24(28) (Xf), C29Δ5,7 (Xk), C27Δ5 (IVc), C29Δ5,22 (IVl), C28Δ5 (IVg), C28Δ5,22 (IVh), C27Δ0 (IIIc), 23-unsaturated C29Δ5,7 (Xk) [13] |
| Glaucophyta | |
| Glaucophyceae | C28Δ5,24(28) (IVf), C29Δ5,22 (IVl), C29Δ5 (IVk) [13,23] |
| Cercozoa | |
| Chlorarachniophyceae | C28Δ5,22 (IVh), C29Δ5,22 (IVl) [24] |
| Rhodophyta | |
| Porphyridiophyceae | C28Δ5,7,22 (ergosterol Xh), C27Δ5,22 (IVd), C28Δ8,22 (XId), C29Δ8,22 (XIh), C28Δ8 (VIIg), C28Δ8 (XIc) [13] |
| Stylonematophyceae | C28Δ5,22 (IVh), C27Δ5 (IVc), C28Δ7 (XIIc), C28Δ7,22 (XIId) [13] |
| Chlorophyta | |
| Prasinophyceae | C29Δ5,24(28) (IVm), C29Δ5,24(28) (IVn) [24], C28Δ5,24(28) (IVf), C29,5,7,22 (Xl), rare C28Δ5,7,9(11),22 (XIIIh), C29Δ5,7,9(11),22 (XIIIl) [10,13] |
| Chlorophyceae | C27Δ5 (IVc), C28Δ5 (IVg), C29Δ5,22 (IVl) C29Δ7,22 (VId), C28Δ7 (VIg) [13,25,26] |
| Trebouxiophyceae | C27Δ5 (IVc), 24S-C29Δ5 (clionasterol IVk), C29Δ5,22 (poriferasterol IVl), C28Δ5 (IVg), C29Δ5,7,22 (7-dehydroporiferasterol Xl), C29Δ7,22 (chondrillasterol VIl), C28Δ8 (VIIg), C28Δ8,22 (VIIh), C28Δ5,7,22 (ergosterol Xh), unusual Δ9(11)-sterols: C28Δ9(11) (VIIIg), C29Δ9(11) (XIVg), C28Δ55,7,9(11),22 (XIIIh), C29Δ5,7,9(11),22 (XIIIl) [13] |
| Chlorodendrophyceae | C28Δ5,24(28) (IVf), C28Δ5 (IVg), C27Δ5 (IVc), C27Δ5,22 (IVd), C27Δ5,24 (IVe) [13] |
| C29Δ5,22 (stigmasterol IVl), C28Δ5 (IVg) [13] | |
| Glycosphingolipids | Taxa | ||
|---|---|---|---|
| Ochrophyta | Dinophyta | Haptophyta | |
| Ceramide moiety | d18:0/16:0; d18:1/16:0; d18:2/16:0; d18:1/22:0; d18:2/22:0; d18:3/22:0; d18:2/22:1; d18:3/23:0; d18:1/24:0; d18:2/24:0; d18:3/24:0; 18:2/24:1; d18:3/24:1; d18:2/24:2 d18:1/26:0;d18:2/26:0 [66,67]; d18:2/14:0; d18:3/14:0 [68] | d18:3/16:0; d18:4/16:0; d18:3/16:1; d18:4/16:1; d19:3/16:0; 19:4/16:1; d19:3/h18:1; 19:3/h19:1; d19:3/h24:1; d19:4/h24:1 [68] | d18:24,8/h18:14; d18:14,8/h24:14 [63]; d18:44,8,10,12/h22:14; 9-methyl-d18:44,8,10,12/h22:14; d18:44,8,10,12/h22:14; d18:14,8,10,12 /h22:0 [64]; h18:14/h22:0; C19:34,8,10/d22:0 [65]; d18:0/h22:0; d18:0/ h22:1; d18:0/h22:2, d19:2/h22:0; d19:2/h22:1; d19:2/h22:2; d18:3/h23:2 and others [68] |
| Glycosyl moiety | Monosaccharide Disaccharide Trisaccharide | Monosaccharide | Glucose [63], galactose [64], glucose and sialic acid [65] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stonik, V.A.; Stonik, I.V. Sterol and Sphingoid Glycoconjugates from Microalgae. Mar. Drugs 2018, 16, 514. https://doi.org/10.3390/md16120514
Stonik VA, Stonik IV. Sterol and Sphingoid Glycoconjugates from Microalgae. Marine Drugs. 2018; 16(12):514. https://doi.org/10.3390/md16120514
Chicago/Turabian StyleStonik, Valentin A., and Inna V. Stonik. 2018. "Sterol and Sphingoid Glycoconjugates from Microalgae" Marine Drugs 16, no. 12: 514. https://doi.org/10.3390/md16120514
APA StyleStonik, V. A., & Stonik, I. V. (2018). Sterol and Sphingoid Glycoconjugates from Microalgae. Marine Drugs, 16(12), 514. https://doi.org/10.3390/md16120514