Specific Chemical and Genetic Markers Revealed a Thousands-Year Presence of Toxic Nodularia spumigena in the Baltic Sea



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
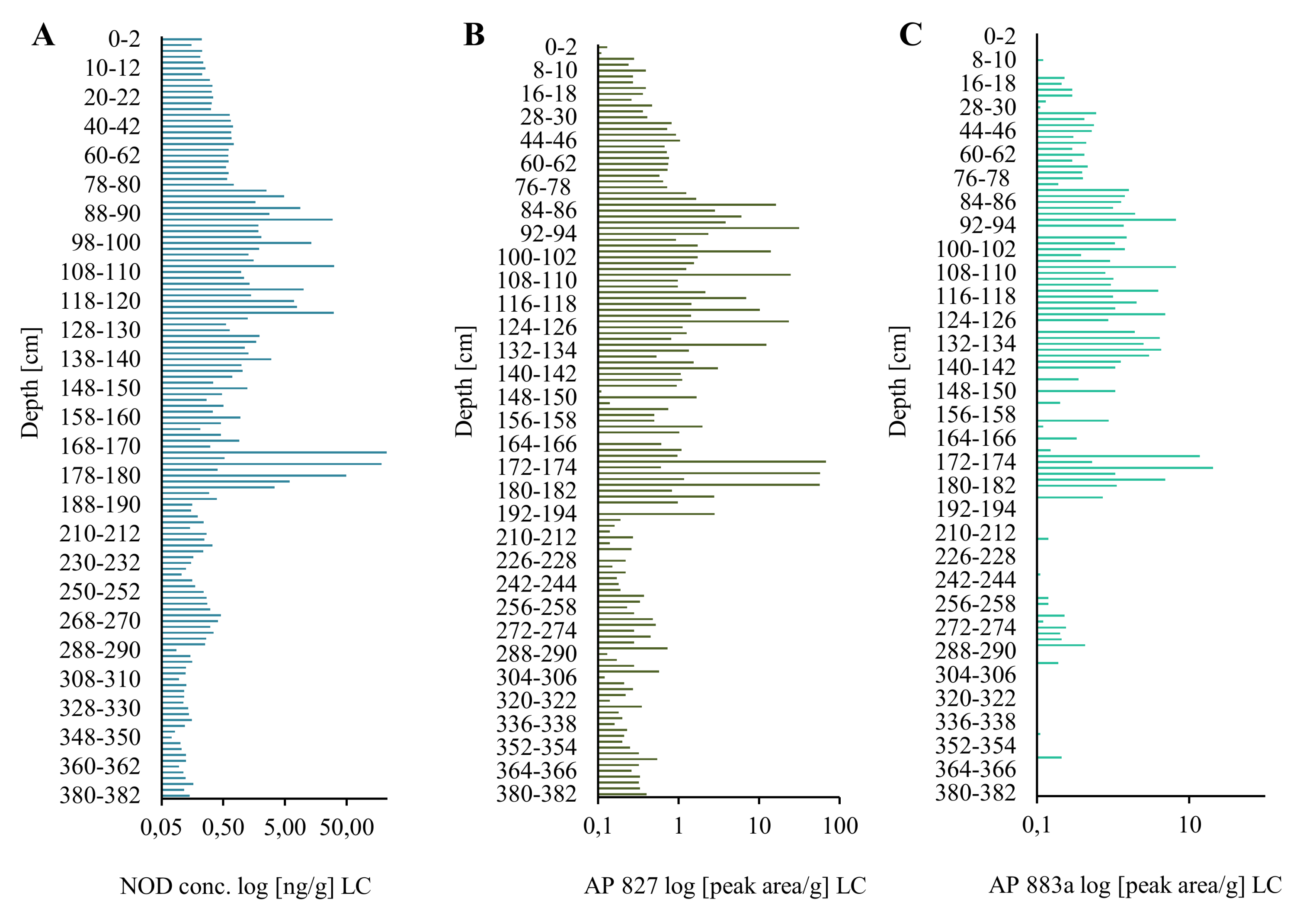
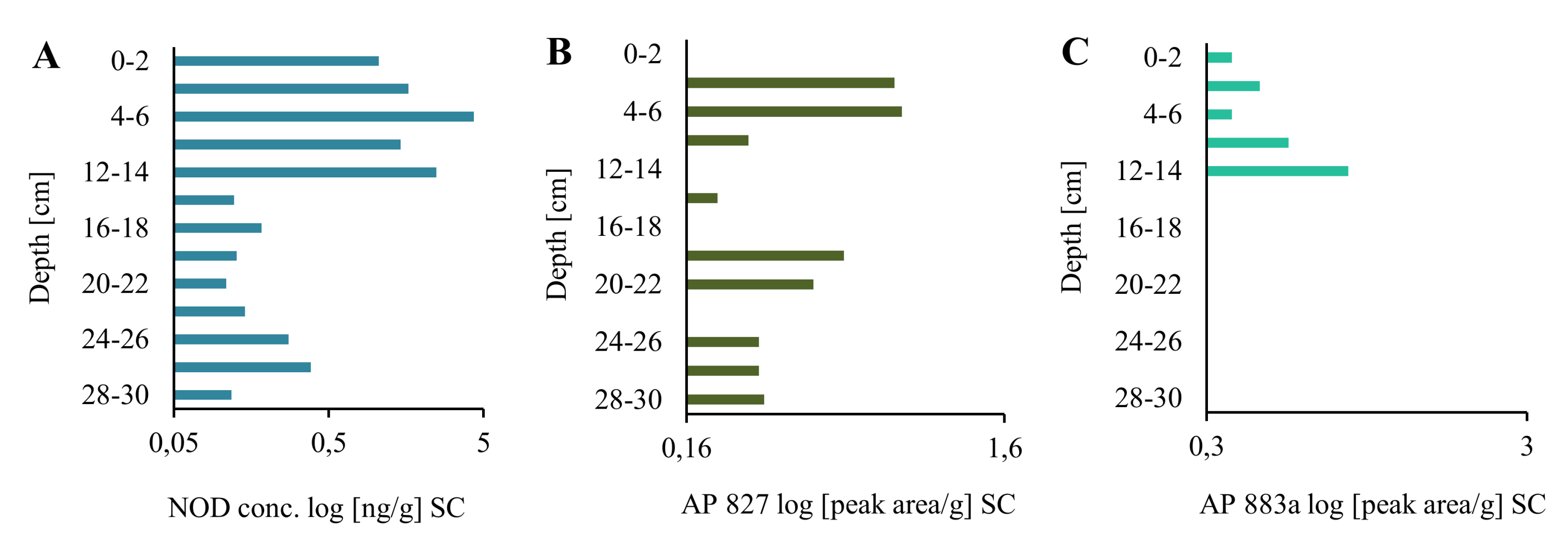
2.1. Extraction and LC-MS/MS Analysis of NOD
2.2. Genetic Analysis of Sediment Samples
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Extraction of Cyanopeptides
4.3. LC-MC/MS Analysis of Cyanopeptides
4.4. DNA Isolation
4.5. Polymerase Chain Reaction (PCR) and Sequence Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bianchi, T.S.; Westman, P.; Andrén, T.; Rolff, C.; Elmgren, R. Cyanobacterial blooms in the Baltic Sea: Natural or human-induced? Limnol. Oceanogr. 2000, 45, 716–726. [Google Scholar] [CrossRef]
- Voss, M.; Kowalewska, G.; Brenner, W. Microfossil and biogeochemical indicators of environmental changes in the Gotland Deep during the last 10.000 years. Baltica 2001, 14, 131–140. [Google Scholar]
- Funkey, C.P.; Conley, D.J.; Reuss, N.S.; Humborg, C.; Jilbert, T.; Slomp, C.P. Hypoxia sustains cyanobacteria blooms in the Baltic Sea. Environ. Sci. Technol. 2014, 48, 2598–2602. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D.E.G.; Summons, R.E. Ancient biomolecules: Their origins, fossilization, and role in revealing the history of life. Bioessa 2014, 36, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Finni, T.; Kononen, K.; Olsonen, R.; Wallström, K. The history of cyanobacterial blooms in the Baltic Sea. Ambio 2001, 30, 172–1788. [Google Scholar] [CrossRef] [PubMed]
- Hübel, H.; Hübel, M. Nitrogen-fixation in the coastal waters of the Baltic Sea. Z. Allg. Mikrobiol. 1974, 14, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Kononen, K.; Carmichael, W.W.; Dahlem, A.M.; Rinehart, K.L.; Kiviranta, J.; Niemelä, S.I. Occurrence of the hepatotoxic cyanobacterium Nodularia spumigena in the Baltic Sea and structure of the toxin. Appl. Environ. Microbiol. 1989, 55, 1990–1995. [Google Scholar] [PubMed]
- Ohta, T.; Sueoka, E.; Iida, N.; Komori, A.; Suganuma, M.; Nishiwaki, R.; Tatematsu, M.; Kim, S.J.; Carmichael, W.W.; Fujiki, H. Nodularin, a potent inhibitor of protein phosphatases 1 and 2A, is a new environmental carcinogen in male F344 rat liver. Cancer Res. 1994, 54, 6402–6406. [Google Scholar] [PubMed]
- Fujiki, H.; Suganuma, M. Tumor promoters–microcystin-LR, nodularin and TNF-α and human cancer development. Anticancer Agents Med. Chem. 2011, 11, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Kankaanpää, H.T.; Sjövall, O.; Huttunen, M.; Olin, M.; Karlsson, K.; Hyvärinen, K.; Sneitz, L.; Härkönen, J.; Sipiä, V.O.; Meriluoto, J.A. Production and sedimentation of peptide toxins nodularin-R and microcystin-LR in the northern Baltic Sea. Environ. Pollut. 2009, 157, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Jokela, J.; Paukku, E.; Österholm, J.; Wahlsten, M.; Permi, P.; Rouhiainen, L.; Gomez-Saez, G.V.; Sivonen, K. New structural variants of aeruginosin produced by the toxic bloom forming cyanobacterium Nodularia spumigena. PLoS ONE 2013, 8, e73618. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Budnjo, A.; Jokela, J.; Haug, B.E.; Fewer, D.P.; Wahlsten, M.; Rouhiainen, L.; Permi, P.; Fossen, T.; Sivonen, K. Pseudoaeruginosins, nonribosomal peptides in Nodularia spumigena. ACS Chem. Biol. 2015, 10, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Marzec, H.; Bertos-Fortis, M.; Toruńska-Sitarz, A.; Fidor, A.; Legrand, C. Chemical and Genetic Diversity of Nodularia spumigena from the Baltic Sea. Mar. Drugs 2016, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Spoof, L.; Błaszczyk, A.; Meriluoto, J.; Cegłowska, M.; Mazur-Marzec, H. Structures and activity of new anabaenopeptins produced by Baltic Sea cyanobacteria. Mar. Drugs 2016, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.T.; Fieker, C.; Neilan, B.A. Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. ISME J. 2012, 6, 1834–1847. [Google Scholar] [CrossRef] [PubMed]
- Jokela, J.; Heinilä, L.M.P.; Shishido, T.K.; Wahlsten, M.; Fewer, D.P.; Fiore, M.F.; Wang, H.; Haapaniemi, E.; Permi, P.; Sivonen, K. Production of high amounts of hepatotoxin nodularin and new protease inhibitors pseudospumigins by the Brazilian benthic Nostoc sp. CENA543. Front. Microbiol. 2017, 8, 1963. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.B.; Sendall, B.C. Iningainema pulvinus gen nov., sp nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 2017, 62, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Szymczak-Żyła, M.; Kowalewska, G. Chloropigments in sediments of the Gulf of Gdańsk deposited during the last 4000 years as indicators of eutrophication and climate change. Palaeoecology 2009, 284, 283–294. [Google Scholar] [CrossRef]
- Okano, K.; Maseda, H.; Sugita, K.; Saito, T.; Utsumi, M.; Maekawa, T.; Kobayashi, M.; Sugiura, N. Biochemical characteristics of microcystin-LR degradation by typical protease. Jpn. J. Water Treat. Biol. 2006, 42, 27–35. [Google Scholar] [CrossRef]
- Kormas, K.A.; Lymperopoulou, D.S. Cyanobacterial toxin degrading bacteria: Who are they? Biomed. Res. Int. 2013, 2013, 463894. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.I.; Tsuji, K.; Watanabe, M.F.; Kondo, F. Stability of microcystins from cyanobacteria—III. Effect of pH and temperature. Phycologia 1996, 35 (Suppl. S6), 83–88. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Microwave oven and boiling waterbath extraction of hepatotoxins from cyanobacterial cells. FEMS Microbiol. Lett. 2000, 184, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Mazur, H.; Pliński, M. Stability of cyanotoxins, microcystin-LR, microcystin-RR and nodularin in seawater and BG-11 medium of different salinity. Oceanologia 2001, 43, 329–339. [Google Scholar]
- Zhang, D.; Xie, P.; Chen, J. Effect of temperature on the stability of microcystins in muscle of fish and its consequences for food safety. Bull. Environ. Contam. Toxicol. 2010, 84, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of Microcystis strain PCC 7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Panieri, G.; Lugli, S.; Manzi, V.; Roveri, M.; Schreiber, B.C.; Palińska, K.A. Ribosomal RNA gene fragments from fossilized cyanobacteria identified in primary gypsum from the late Miocene, Italy. Geobiology 2010, 8, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Lyra, K.; Sinkko, H.; Rantanen, M.; Paulin, L.; Kotilainen, A. Sediment bacterial communities reflect the history of a Sea Basin. PLoS ONE 2013, 8, e54326. [Google Scholar] [CrossRef] [PubMed]
- Webster, G.; Newberry, C.J.; Fry, J.C.; Weightman, A.J. Assessment of bacterial community structure in the deep sub-seafloor biosphere by 16S rDNA-based techniques: A cautionary tale. J. Microbiol. Methods 2003, 55, 155–164. [Google Scholar] [CrossRef]
- Bowman, J.P.; Rea, S.M.; McCammon, S.A.; McMeekin, T.A. Diversity and community structure within anoxic sediment from marine salinity meromictic lakes and a coastal meromictic marine basin, Vestfold Hills, Eastern Antarctica. Environ. Microbiol. 2000, 2, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Dadheech, P.K.; Krienitz, L.; Kotut, K.; Ballot, A.; Casper, P. Molecular detection of uncultured cyanobacteria and aminotransferase domains for cyanotoxin production in sediments of different Kenyan lakes. FEMS Microbiol. Ecol. 2009, 68, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Saxton, M.A.; DeBruyn, J.M.; Smith, J.L.; Marvin, C.H.; Krieger, K.A.; Sayler, G.S.; Boyer, L.G.; Wilhelm, S.W. The diversity and distribution of toxigenic Microcystis spp. in present day and archived pelagic and sediment samples from Lake Erie. Harmful Algae 2009, 8, 385–394. [Google Scholar] [CrossRef]
- Savichtcheva, O.; Debroas, D.; Kurmayer, R.; Villar, C.; Jenny, J.P.; Arnaud, F.; Perga, M.E.; Domaizon, I. Quantitative PCR enumeration of total/toxic Planktothrix rubescens and total cyanobacteria in preserved DNA isolated from lake sediments. Appl. Environ. Microbiol. 2011, 77, 8744–8753. [Google Scholar] [CrossRef] [PubMed]
- Domaizon, I.; Savichtcheva, O.; Debroas, D.; Arnaud, F.; Villar, C.; Pignol, C.; Alric, B.; Perga, M.E. DNA from lake sediments reveals the long-term dynamics and diversity of Synechococcus assemblages. Biogeosciences 2013, 10, 3817–3838. [Google Scholar] [CrossRef]
- De La Escalera, M.G.; Antoniades, D.; Bonilla, S.; Piccini, C. Application of ancient DNA to the reconstruction of past microbial assemblages and for the detection of toxic cyanobacteria in subtropical freshwater ecosystems. Mol. Ecol. 2014, 23, 5791–5802. [Google Scholar] [CrossRef] [PubMed]
- Monchamp, M.E.; Walser, J.C.; Pomati, F.; Spaak, P. Sedimentary DNA reveals cyanobacterial community diversity over 200 years in two perialpine lakes. Appl. Environ. Microbiol. 2016, 82, 6472–6482. [Google Scholar] [CrossRef] [PubMed]
- Kyle, M.; Haande, S.; Sřnstebř, J.; Rohrlack, T. Amplification of DNA in sediment cores to detect historic Planktothrix occurrence in three Norwegian lakes. J. Paleolimnol. 2015, 53, 61–72. [Google Scholar] [CrossRef]
- Pal, S.; Gregory-Eaves, I.; Pick, F.R. Temporal trends in cyanobacteria revealed through DNA and pigment analyses of temperate lake sediment cores. J. Paleolimnol. 2015, 54, 87–101. [Google Scholar] [CrossRef]
- Fernandez-Carazomailto, R.; Verleyen, E.; Hodgson, D.A.; Roberts, S.J.; Waleron, K.; Vyverman, W.; Wilmotte, A. Late Holocene changes in cyanobacterial community structure in maritime Antarctic lakes. J. Paleolimnol. 2013, 50, 15–31. [Google Scholar] [CrossRef]
- Hou, W.; Dong, H.; Li, G.; Yang, J.; Coolen, M.J.; Liu, X.; Wang, S.; Jiang, H.; Wu, X.; Xiao, H.; et al. Identification of photosynthetic plankton communities using sedimentary ancient DNA and their response to late-Holocene climate change on the Tibetan Plateau. Sci. Rep. 2014, 4, 6648. [Google Scholar] [CrossRef] [PubMed]
- Coolen, M.J.L.; Talbot, H.M.; Abbas, B.A.; Ward, C.; Schouten, S.; Volkman, J.K.; Damsté, J.S.S. Sources for sedimentary bacteriohopanepolyols as revealed by 16S rDNA stratigraphy. Environ. Microbiol. 2008, 10, 1783–1803. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Dong, H.; Hou, W.; Wang, S.; Jiang, H.; Yang, J.; Wu, G. Temporal succession of ancient phytoplankton community in Qinghai Lake and implication for paleo-environmental change. Sci. Rep. 2016, 6, 19769. [Google Scholar] [CrossRef] [PubMed]
- Barker, G.L.A.; Hayes, P.K.; O’Mahony, S.L.; Vacharapiyasophon, P.; Walsby, A.E. A molecular and phenotypic analysis of Nodularia (cyanobacteria) from the Baltic Sea. J. Phycol. 1999, 35, 931–937. [Google Scholar] [CrossRef]
- Laamanen, M.J.; Gugger, M.F.; Lehtimäki, J.; Haukka, K.; Sivonen, K. Diversity of toxic and non-toxic Nodularia isolates (Cyanobacteria) and filaments from the Baltic Sea. Appl. Environ. Microbiol. 2001, 67, 4638–4647. [Google Scholar] [CrossRef] [PubMed]
- Janson, S.; Granéli, E. Phylogenetic analyses of nitrogen-fixing cyanobacteria from Baltic Sea reveal sequence anomalies in the phycocyanin operon. Int. J. Syst. Evol. Microbiol. 2002, 52, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Rounge, T.B.; Rohrlack, T.; Decenciere, B.; Edvardsen, B.; Kristensen, T.; Jakobsen, K.S. Subpopulation differentiation associated with nonribosomal peptide synthetase gene cluster dynamics in the cyanobacterium Planktothrix spp. J. Phycol. 2010, 46, 645–652. [Google Scholar] [CrossRef]
- Sønstebø, J.H.; Rohrlack, T. Possible implication of chytrid parasitism for population subdivision in freshwater cyanobacteria of the genus Planktothrix. Appl. Environ. Microbiol. 2011, 77, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Sogge, H.; Rohrlack, T.; Rounge, T.B.; Sønstebe, J.H.; Tooming-Klunderud, A.; Kristensen, T.; Jakobsen, K.S. Gene flow, recombination, and selection in cyanobacteria: Population structure of geographically related Planktothrix freshwater strains. Appl. Environ. Microbiol. 2013, 79, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Lehtimäki, J.; Moisander, P.; Sivonen, K.; Kononen, K. Growth, nitrogen fixation, and nodularin production by two Baltic Sea cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 1647–1656. [Google Scholar] [PubMed]
- Hobson, P.; Fallowfield, H. Effect of irradiance, temperature and salinity on growth and toxin production by Nodularia spumigena. Hydrobiologia 2003, 493, 7–15. [Google Scholar] [CrossRef]
- Wang, T.; Surge, D.; Mithen, S. Seasonal temperature variability of the Neoglacial (3300-2500 BP) and Roman Warm Period (2500-1600 BP) reconstructed from oxygen isotope ratios of limpet shells (Patella vulgata), Northwest Scotland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 317–318, 104–113. [Google Scholar] [CrossRef]
- Büntgen, U.; Myglan, V.S.; Ljungqvist, F.C.; McCormick, M.; Di Cosmo, N.; Sigl, M.; Jungclaus, J.; Wagner, S.; Krusic, P.J.; Esper, J.; et al. Cooling and societal change during the Late Antique Little Ice Age from 536 to around 660 AD. Nat. Geosci. 2016, 9, 231–236. [Google Scholar] [CrossRef]
- Kabel, K.; Moros, M.; Porsche, C.; Neumann, T.; Adolphi, F.; Andersen, T.J.; Siegel, H.; Gerth, M.; Leipe, T.; Jansen, A.; Damsté, J.S.S. Impact of climate change on the Baltic Sea ecosystem over the past 1000 years. Nat. Clim. Chang. 2012, 2, 871–874. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The Rise of Harmful Cyanobacteria Blooms: Potential Role of Eutrophication and Climate Change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Šulčius, S.; Pilkaityte, R.; Mazur-Marzec, H.; Kasperovičiene, J.; Ezhova, E.; Błaszczyk, A.; Paškauskas, R. Increased risk of exposure to microcystins in the scum of the filamentous cyanobacterium Aphanizomenon flos-aquae accumulated on the western shoreline of the Curonian Lagoon. Mar. Poll. Bull. 2015, 99, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Jacobs, D.; Goodman, A.E. Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl. Environ. Microbiol. 1995, 61, 3875–3883. [Google Scholar] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cegłowska, M.; Toruńska-Sitarz, A.; Kowalewska, G.; Mazur-Marzec, H. Specific Chemical and Genetic Markers Revealed a Thousands-Year Presence of Toxic Nodularia spumigena in the Baltic Sea. Mar. Drugs 2018, 16, 116. https://doi.org/10.3390/md16040116
Cegłowska M, Toruńska-Sitarz A, Kowalewska G, Mazur-Marzec H. Specific Chemical and Genetic Markers Revealed a Thousands-Year Presence of Toxic Nodularia spumigena in the Baltic Sea. Marine Drugs. 2018; 16(4):116. https://doi.org/10.3390/md16040116
Chicago/Turabian StyleCegłowska, Marta, Anna Toruńska-Sitarz, Grażyna Kowalewska, and Hanna Mazur-Marzec. 2018. "Specific Chemical and Genetic Markers Revealed a Thousands-Year Presence of Toxic Nodularia spumigena in the Baltic Sea" Marine Drugs 16, no. 4: 116. https://doi.org/10.3390/md16040116
APA StyleCegłowska, M., Toruńska-Sitarz, A., Kowalewska, G., & Mazur-Marzec, H. (2018). Specific Chemical and Genetic Markers Revealed a Thousands-Year Presence of Toxic Nodularia spumigena in the Baltic Sea. Marine Drugs, 16(4), 116. https://doi.org/10.3390/md16040116