Structural Insights into the Cytotoxic Mechanism of Vibrio parahaemolyticus PirAvp and PirBvp Toxins



Abstract
:1. Introduction
2. The Structural Similarity between V. parahaemolyticus PirAvp/PirBvp and Bacillus thuringiensis Cry Toxins
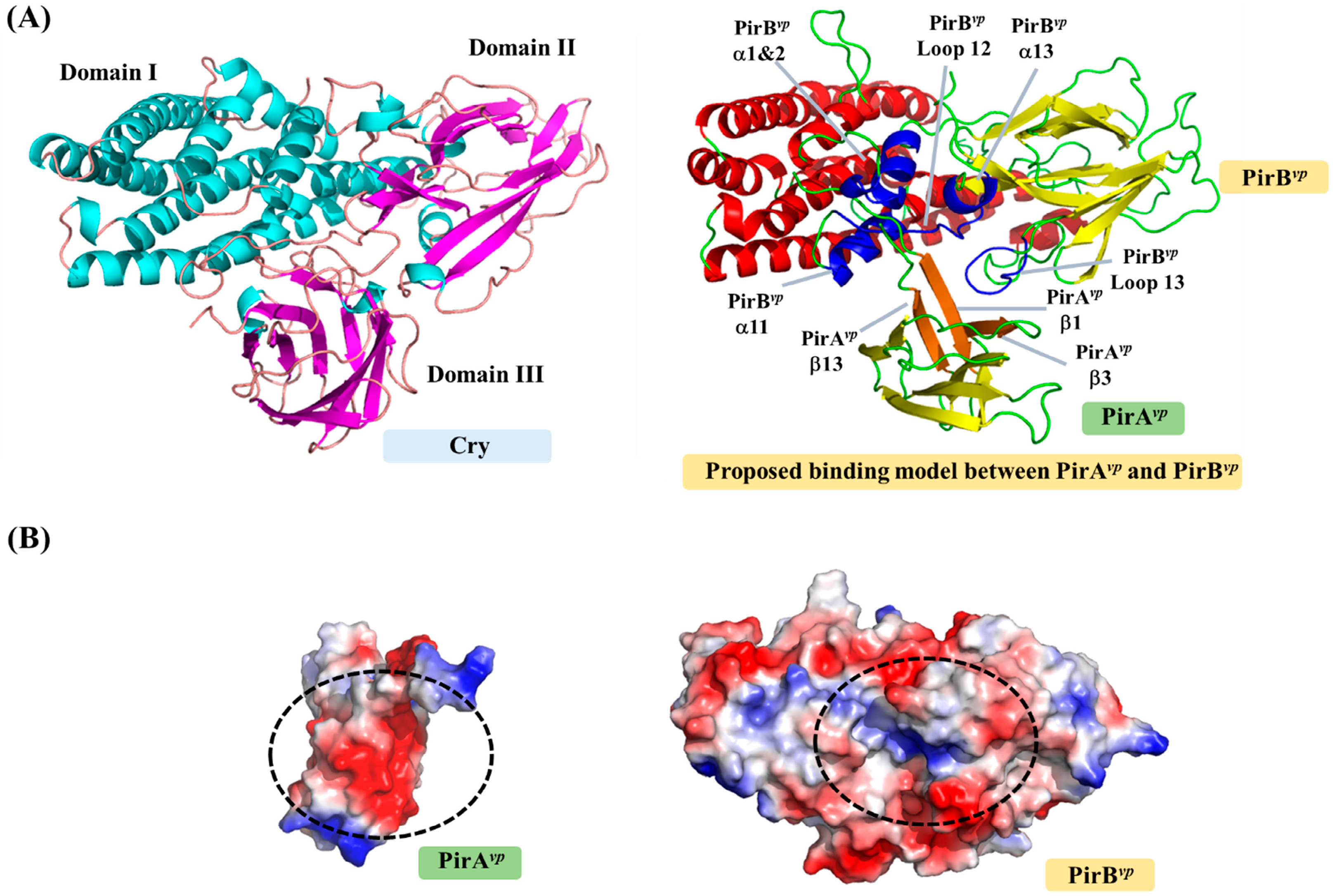
2.1. PirBvp Contains Both Cry-Like Pore-Forming and Receptor Domains
2.2. PirAvp Contains a Possible Sugar-Binding Pocket
2.3. Unanswered Questions Relating to the Cytotoxic Mechanism of PirAvp/PirBvp
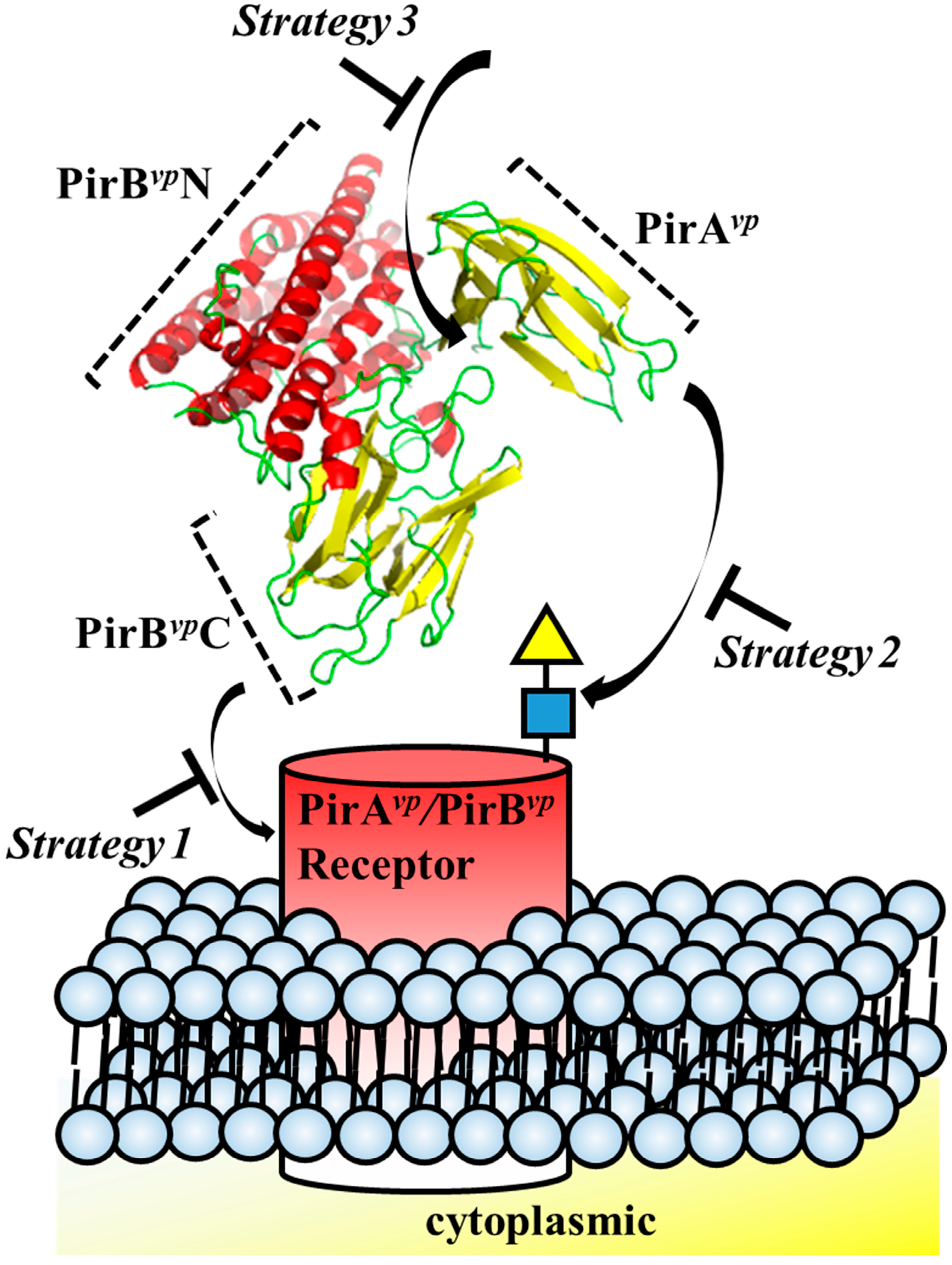
3. Strategies for Designing Drugs to Block the Cytotoxic Effects of V. parahaemolyticus PirAvp and PirBvp Toxins
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gomez-Gil, B.; Soto-Rodriguez, S.; Lozano, R.; Betancourt-Lozano, M. Draft Genome Sequence of Vibrio parahaemolyticus Strain M0605, Which Causes Severe Mortalities of Shrimps in Mexico. Genome Announc. 2014, 2, e00055-14. [Google Scholar] [CrossRef] [PubMed]
- Nunan, L.; Lightner, D.; Pantoja, C.; Gomez-Jimenez, S. Detection of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Dis. Aquat. Org. 2014, 111, 81–86. [Google Scholar] [CrossRef] [PubMed]
- FAO Fisheries and Aquaculture. Report of the FAO/MARD Technical Workshop on Early Mortality Syndrome (EMS) or Acute Hepatopancreatic Necrosis Syndrome (AHPNS) of Cultured Shrimp (under TCP/VIE/3304). Rep. No. 1053. Available online: www.fao.org/docrep/018/i3422e/i3422e.pdf (accessed on 31 October 2013).
- Tran, L.; Nunan, L.; Redman, R.; Mohney, L.; Pantoja, C.; Fitzsimmons, K.; Lightner, D. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Org. 2013, 105, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Lightner, D.V.; Redman, R.M.; Pantoja, C.R.; Noble, B.L.; Tran, L. Early mortality syndrome affects shrimp in Asia. Glob. Aquac. Advocate 2012, 15, 40. [Google Scholar]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano-Olvera, R.; Betancourt-Lozano, M.; Morales-Covarrubias, M.S. Field and Experimental Evidence of Vibrio parahaemolyticus as the Causative Agent of Acute Hepatopancreatic Necrosis Disease of Cultured Shrimp (Litopenaeus vannamei) in Northwestern Mexico. Appl. Environ. Microbiol. 2014, 81, 1689–1699. [Google Scholar] [CrossRef] [PubMed]
- Sirikharin, R.; Taengchaiyaphum, S.; Sanguanrut, P.; Chi, T.D.; Mavichak, R.; Proespraiwong, P.; Nuangsaeng, B.; Thitamadee, S.; Flegel, T.W.; Sritunyalucksana, K. Characterization and PCR Detection Of Binary, Pir-Like Toxins from Vibrio parahaemolyticus Isolates that Cause Acute Hepatopancreatic Necrosis Disease (AHPND) in Shrimp. PLoS ONE 2015, 10, e0126987. [Google Scholar] [CrossRef] [PubMed]
- Joshi, J.; Srisala, J.; Truong, V.H.; Chen, I.T.; Nuangsaeng, B.; Suthienkul, O.; Lo, C.F.; Flegel, T.W.; Sritunyalucksana, K.; Thitamadee, S. Variation in Vibrio parahaemolyticus isolates from a single Thai shrimp farm experiencing an outbreak of acute hepatopancreatic necrosis disease (AHPND). Aquaculture 2014, 428–429, 297–302. [Google Scholar] [CrossRef]
- Yang, Y.-T.; Chen, I.-T.; Lee, C.-T.; Chen, C.-Y.; Lin, S.-S.; Hor, L.-I.; Tseng, T.-C.; Huang, Y.-T.; Sritunyalucksana, K.; Thitamadee, S.; et al. Draft Genome Sequences of Four Strains of Vibrio parahaemolyticus, Three of Which Cause Early Mortality Syndrome/Acute Hepatopancreatic Necrosis Disease in Shrimp in China and Thailand. Genome Announc. 2014, 2, e00816-14. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Tinwongger, S.; Proespraiwong, P.; Mavichak, R.; Unajak, S.; Nozaki, R.; Hirono, I. Draft Genome Sequences of Six Strains of Vibrio parahaemolyticus Isolated from Early Mortality Syndrome/Acute Hepatopancreatic Necrosis Disease Shrimp in Thailand. Genome Announc. 2014, 2, e00221-14. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Jimenez, S.; Noriega-Orozco, L.; Sotelo-Mundo, R.R.; Cantu-Robles, V.A.; Cobian-Guemes, A.G.; Cota-Verdugo, R.G.; Gamez-Alejo, L.A.; Del Pozo-Yauner, L.; Guevara-Hernandez, E.; Garcia-Orozco, K.D.; et al. High-quality draft genomes of two Vibrio parahaemolyticus strains aid in understanding acute hepatopancreatic necrosis disease of cultured shrimps in Mexico. Genome Announc. 2014, 2, e00800-14. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Tang, K.; Tran, L.; Lightner, D. Photorhabdus insect-related (Pir) toxin-like genes in a plasmid of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease (AHPND) of shrimp. Dis. Aquat. Org. 2015, 113, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Chen, I.-T.; Yang, Y.-T.; Ko, T.-P.; Huang, Y.-T.; Huang, J.-Y.; Huang, M.-F.; Lin, S.-J.; Chen, C.-Y.; Lin, S.-S.; et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc. Natl. Acad. Sci. USA 2015, 112, 10798–10803. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.-C.; Ng, T.H.; Ando, M.; Lee, C.-T.; Chen, I.-T.; Chuang, J.-C.; Mavichak, R.; Chang, S.-H.; Yeh, M.-D.; Chiang, Y.-A.; et al. Pathogenesis of acute hepatopancreatic necrosis disease (AHPND) in shrimp. Fish Shellfish Immunol. 2015, 47, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Tinwongger, S.; Nochiri, Y.; Thawonsuwan, J.; Nozaki, R.; Kondo, H.; Awasthi, S.P.; Hinenoya, A.; Yamasaki, S.; Hirono, I. Virulence of acute hepatopancreatic necrosis disease PirAB-like relies on secreted proteins not on gene copy number. J. Appl. Microbiol. 2016, 121, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Theethakaew, C.; Nakamura, S.; Motooka, D.; Matsuda, S.; Kodama, T.; Chonsin, K.; Suthienkul, O.; Iida, T. Plasmid dynamics in Vibrio parahaemolyticus strains related to shrimp Acute Hepatopancreatic Necrosis Syndrome (AHPNS). Infect. Genet. Evol. 2017, 51, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.H.; Waterfield, N.; Burland, V.; Perna, N.T.; Daborn, P.J.; Bowen, D.; Blattner, F.R. A Genomic Sample Sequence of the Entomopathogenic Bacterium Photorhabdus luminescens W14: Potential Implications for Virulence. Appl. Environ. Microbiol. 2000, 66, 3310–3329. [Google Scholar] [CrossRef] [PubMed]
- Ahantarig, A.; Chantawat, N.; Waterfield, N.R.; ffrench-Constant, R.; Kittayapong, P. PirAB Toxin from Photorhabdus asymbiotica as a Larvicide against Dengue Vectors. Appl. Environ. Microbiol. 2009, 75, 4627–4629. [Google Scholar] [CrossRef] [PubMed]
- Duchaud, E.; Rusniok, C.; Frangeul, L.; Buchrieser, C.; Givaudan, A.; Taourit, S.; Bocs, S.; Boursaux-Eude, C.; Chandler, M.; Charles, J.F.; et al. The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens. Nat. Biotechnol. 2003, 21, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Waterfield, N.; Kamita, S.G.; Hammock, B.D.; ffrench-Constant, R. The Photorhabdus Pir toxins are similar to a developmentally regulated insect protein but show no juvenile hormone esterase activity. FEMS Microbiol. Lett. 2005, 245, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.H.; Dowling, A.; Waterfield, N.R. Insecticidal toxins from Photorhabdus bacteria and their potential use in agriculture. Toxicon 2007, 49, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, X.; Zhang, X.; Liu, Z.; Ding, X.; Xia, L.; Hu, S. Photorhabdus luminescens PirAB-fusion protein exhibits both cytotoxicity and insecticidal activity. FEMS Microbiol. Lett. 2014, 356, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Adang, M.; Crickmore, N.; Jurat-Fuentes, J. Diversity of Bacillus thuringiensis Crystal Toxins and Mechanism of Action. Adv. Insect Physiol. 2014, 47, 39–87. [Google Scholar] [CrossRef]
- Bravo, A.; Gómez, I.; Porta, H.; García-Gómez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberón, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2012, 6, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.; Crickmore, N. Structural classification of insecticidal proteins—Towards an in silico characterisation of novel toxins. J. Invertebr. Pathol. 2017, 142, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal δ-endotoxin from Bacillus thuringiensis at 2.5 Å resolution. Nature 1991, 353, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Grochulski, P.; Masson, L.; Borisova, S.; Pusztai-Carey, M.; Schwartz, J.L.; Brousseau, R.; Cygler, M. Bacillus thuringiensis CryIA(a) insecticidal toxin: Crystal structure and channel formation. J. Mol. Biol. 1995, 254, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Soberón, M.; Pardo, L.; Muñóz-Garay, C.; Sánchez, J.; Gómez, I.; Porta, H.; Bravo, A. Pore formation by Cry toxins. Adv. Exp. Med. Biol. 2010, 677, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef] [PubMed]
- Peraro, M.D.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2015, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.W.; Pattus, F.; Tucker, A.D.; Tsernoglou, D. Structure of the membrane-pore-forming fragment of colicin A. Nature 1989, 337, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.F.; Barclay, A.N. The immunoglobulin superfamily—Domains for cell surface recognition. Annu. Rev. Immunol. 1988, 6, 381–405. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.C.; Chen, Y.F.; Lin, S.R.; Yang, J.M. iGEMDOCK: A graphical environment of enhancing GEMDOCK using pharmacological interactions and post-screening analysis. BMC Bioinform. 2011, 12, S33. [Google Scholar] [CrossRef] [PubMed]
- Kitami, M.; Kadotani, T.; Nakanishi, K.; Atsumi, S.; Higurashi, S.; Ishizaka, T.; Watanabe, A.; Sato, R. Bacillus thuringiensis Cry Toxins Bound Specifically to Various Proteins via Domain III, Which Had a Galactose-Binding Domain-Like Fold. Biosci. Biotechnol. Biochem. 2011, 75, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Sarkar, A.; Priya, P.; Ghosh Dastidar, S.; Das, S. New Insight to Structure-Function Relationship of GalNAc Mediated Primary Interaction between Insecticidal Cry1Ac Toxin and HaALP Receptor of Helicoverpa armigera. PLoS ONE 2013, 8, e78249. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.L. Bivalent Sequential Binding Model of a Bacillus thuringiensis Toxin to Gypsy Moth Aminopeptidase N Receptor. J. Biol. Chem. 2000, 275, 14423–14431. [Google Scholar] [CrossRef] [PubMed]
- Gomez, I.; Oltean, D.I.; Gill, S.S.; Bravo, A.; Soberon, M. Mapping the epitope in cadherin-like receptors involved in Bacillus thuringiensis Cry1A toxin interaction using phage display. J. Biol. Chem. 2001, 276, 28906–28912. [Google Scholar] [CrossRef] [PubMed]
- Gomez, I.; Miranda-Rios, J.; Rudino-Pinera, E.; Oltean, D.I.; Gill, S.S.; Bravo, A.; Soberon, M. Hydropathic Complementarity Determines Interaction of Epitope (869)HITDTNNK(876) in Manduca sexta Bt-R1 Receptor with Loop 2 of Domain II of Bacillus thuringiensis Cry1A Toxins. J. Biol. Chem. 2002, 277, 30137–30143. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Dean, D.H.; Bravo, A.; Soberón, M. Molecular Basis for Bacillus thuringiensis Cry1Ab Toxin Specificity: Two Structural Determinants in the Manduca sexta Bt-R1Receptor Interact with Loops α-8 and 2 in Domain II of Cy1Ab Toxin. Biochemistry 2003, 42, 10482–10489. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, M.R.; Pyo, S.J.; Shin, I. Carbohydrate Chips for Studying High-Throughput Carbohydrate−Protein Interactions. J. Am. Chem. Soc. 2004, 126, 4812–4819. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Kim, C.S.; Hwang, B.H.; Cha, H.J. A functional carbohydrate chip platform for analysis of carbohydrate–protein interaction. Nanotechnology 2010, 21, 215101. [Google Scholar] [CrossRef] [PubMed]
- Dan, X.; Liu, W.; Ng, T. Development and applications of lectins as biological tools in biomedical research. Med. Res. Rev. 2016, 36, 221–247. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, H.E.; Crickmore, N.; van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [PubMed]
- Adalat, R.; Saleem, F.; Crickmore, N.; Naz, S.; Shakoori, A.R. In vivo crystallization of three-domain Cry toxins. Toxins 2017, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Koller, C.N.; Bauer, L.S.; Hollingworth, R.M. Characterization of the pH-mediated solubility of Bacillus thuringiensis var. san diego native δ-endotoxin crystals. Biochem. Biophys. Res. Commun. 1992, 184, 692–699. [Google Scholar] [CrossRef]
- Deist, B.; Rausch, M.; Fernandez-Luna, M.; Adang, M.; Bonning, B. Bt Toxin Modification for Enhanced Efficacy. Toxins 2014, 6, 3005–3027. [Google Scholar] [CrossRef] [PubMed]
- Flegel, T.W.; Lo, C.F. Free Release of Primers for Specific Detection of Bacterial Isolates that Cause Acute Hepatopancreatic Necrosis Disease (AHPND); Network of Aquaculture Centres in Asia and the Pacific (NACA): Bangkok, Thailand, 2013. [Google Scholar]
- Dangtip, S.; Sirikharin, R.; Sanguanrut, P.; Thitamadee, S.; Sritunyalucksana, K.; Taengchaiyaphum, S.; Mavichak, R.; Proespraiwong, P.; Flegel, T.W. AP4 method for two-tube nested PCR detection of AHPND isolates of Vibrio parahaemolyticus. Aquac. Rep. 2015, 2, 158–162. [Google Scholar] [CrossRef]
- Wu, Q.; Guo, Z. Glycosylphosphatidylinositols are potential targets for the development of novel inhibitors for aerolysin-type of pore-forming bacterial toxins. Med. Res. Rev. 2010, 30, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Foletti, D.; Strop, P.; Shaughnessy, L.; Hasa-Moreno, A.; Casas, M.G.; Russell, M.; Bee, C.; Wu, S.; Pham, A.; Zeng, Z.; et al. Mechanism of Action and In Vivo Efficacy of a Human-Derived Antibody against Staphylococcus aureus α-Hemolysin. J. Mol. Biol. 2013, 425, 1641–1654. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F., III; Kozhaya, L.; Rawlings, S.A.; Reyes-Robles, T.; DuMont, A.L.; Myszka, D.G.; Landau, N.R.; Unutmaz, D.; Torres, V.J. CCR5 is a receptor for Staphylococcus aureus leukotoxin ED. Nature 2012, 493, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Inoshima, I.; Inoshima, N.; Wilke, G.A.; Powers, M.E.; Frank, K.M.; Wang, Y.; Wardenburg, J.B. A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 2011, 17, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Sliwoski, G.; Kothiwale, S.; Meiler, J.; Lowe, E.W. Computational methods in drug discovery. Pharmacol. Rev. 2014, 66, 334–395. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.J.; Macias, A.T.; MacKerell, A.D. Computational identification of inhibitors of protein-protein interactions. Curr. Top. Med. Chem. 2007, 7, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Shoichet, B.K. ZINC—A Free Database of Commercially Available Compounds for Virtual Screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Eckert, H.; Bajorath, J. Molecular similarity analysis in virtual screening: Foundations, limitations and novel approaches. Drug Discov. Today 2007, 12, 225–233. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potential Function | Regions Involved in Possible Interactions | Amino Acid Sequences |
|---|---|---|
| Receptor binding | PirBvp Loop 12 | 274-VGFPS-278 |
| PirBvp Loop 14 | 316-SIEIHYYNRV-325 | |
| PirBvp Loop 18 | 369-GPE-371 | |
| PirBvp Loop 22 | 413-QEGSDKV-419 | |
| PirAvp/PirBvp complex formation | PirAvp β-sheet 1 | 11-YSHDWTV-17 |
| PirAvp β-sheet 3 | 26-VDSKH-30 | |
| PirAvp β-sheet 9 | 104-GFCTIYY-110 | |
| PirBvp α-helix 1 | 35-YAFKAMVSFG-43 | |
| PirBvp α-helix 2 | 45-LSN-47 | |
| PirBvp α-helix 12 | 247-MILWQKIKEL-256 | |
| PirBvp α-helix 13 | 260-DVFVHSNLISY-270 | |
| PirBvp Loop 12 | 298-PNMFGERR-305 | |
| PirBvp Loop 13 | 431-PDEF-434 | |
| Ligand binding | PirAvp β-sheet 3 | 26-VDSKH-30 * |
| PirAvp Loop 4 | 31-TPIIPEVGRS-40 * | |
| PirAvp Loop 8 | 83-QRPDNAFY-90 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-J.; Hsu, K.-C.; Wang, H.-C. Structural Insights into the Cytotoxic Mechanism of Vibrio parahaemolyticus PirAvp and PirBvp Toxins. Mar. Drugs 2017, 15, 373. https://doi.org/10.3390/md15120373
Lin S-J, Hsu K-C, Wang H-C. Structural Insights into the Cytotoxic Mechanism of Vibrio parahaemolyticus PirAvp and PirBvp Toxins. Marine Drugs. 2017; 15(12):373. https://doi.org/10.3390/md15120373
Chicago/Turabian StyleLin, Shin-Jen, Kai-Cheng Hsu, and Hao-Ching Wang. 2017. "Structural Insights into the Cytotoxic Mechanism of Vibrio parahaemolyticus PirAvp and PirBvp Toxins" Marine Drugs 15, no. 12: 373. https://doi.org/10.3390/md15120373
APA StyleLin, S. -J., Hsu, K. -C., & Wang, H. -C. (2017). Structural Insights into the Cytotoxic Mechanism of Vibrio parahaemolyticus PirAvp and PirBvp Toxins. Marine Drugs, 15(12), 373. https://doi.org/10.3390/md15120373