Attenuation of Metabolic Syndrome by EPA/DHA Ethyl Esters in Testosterone-Deficient Obese Rats


 and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of High-Carbohydrate, High-Fat Diet
2.1.1. Dietary Intakes, Body Composition, and Metabolic Parameters
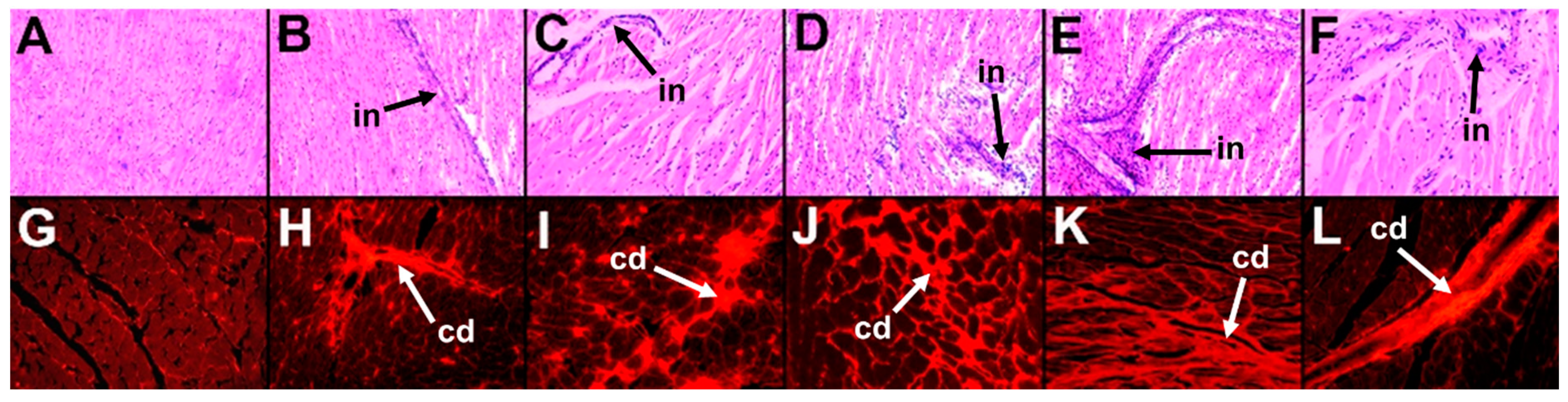
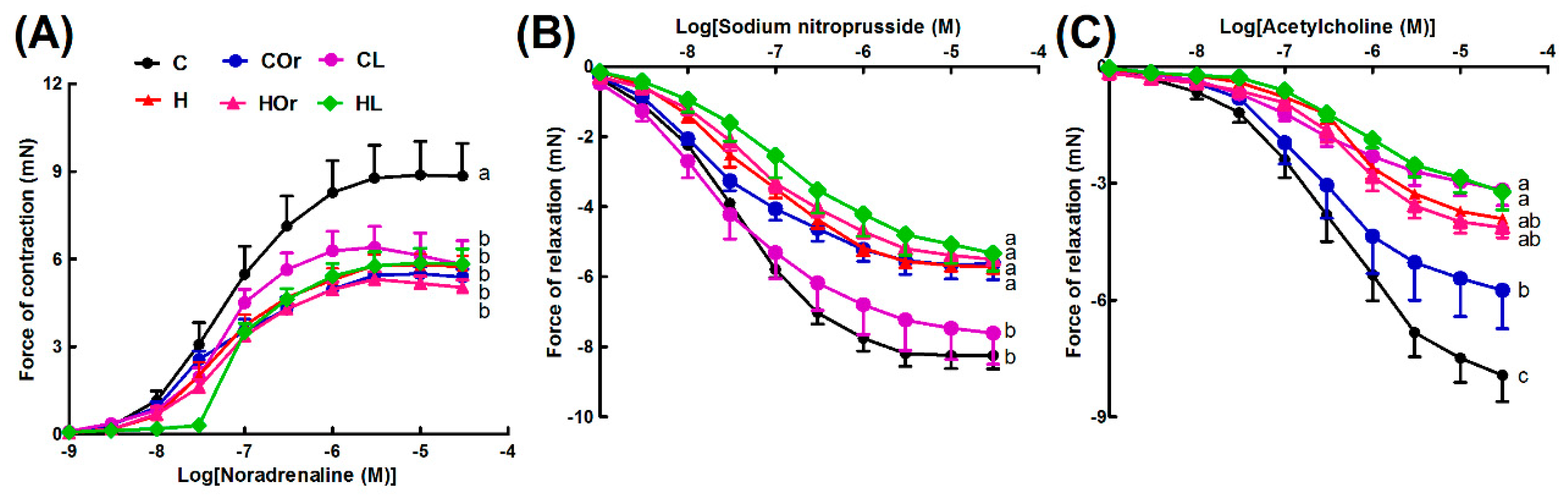
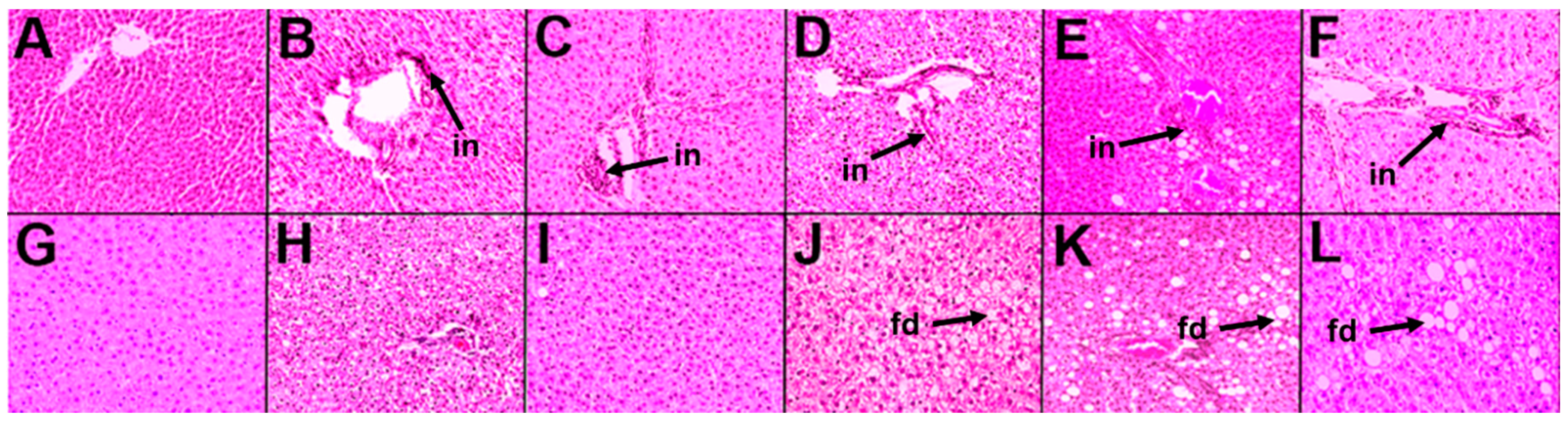
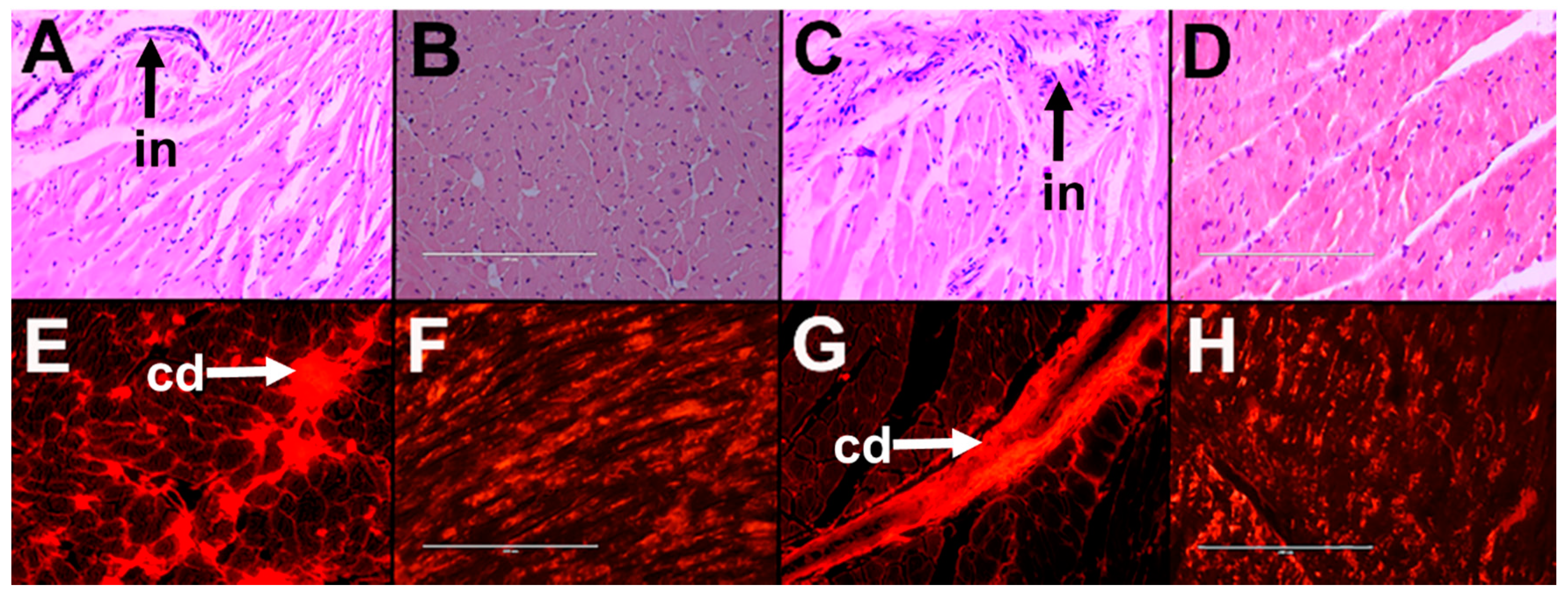
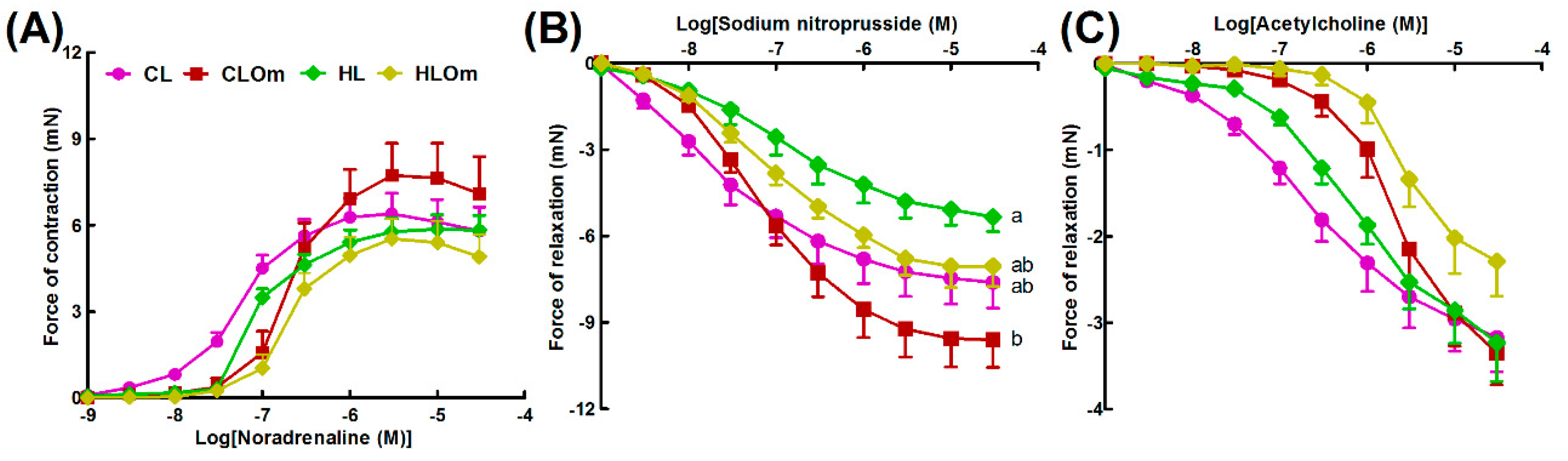
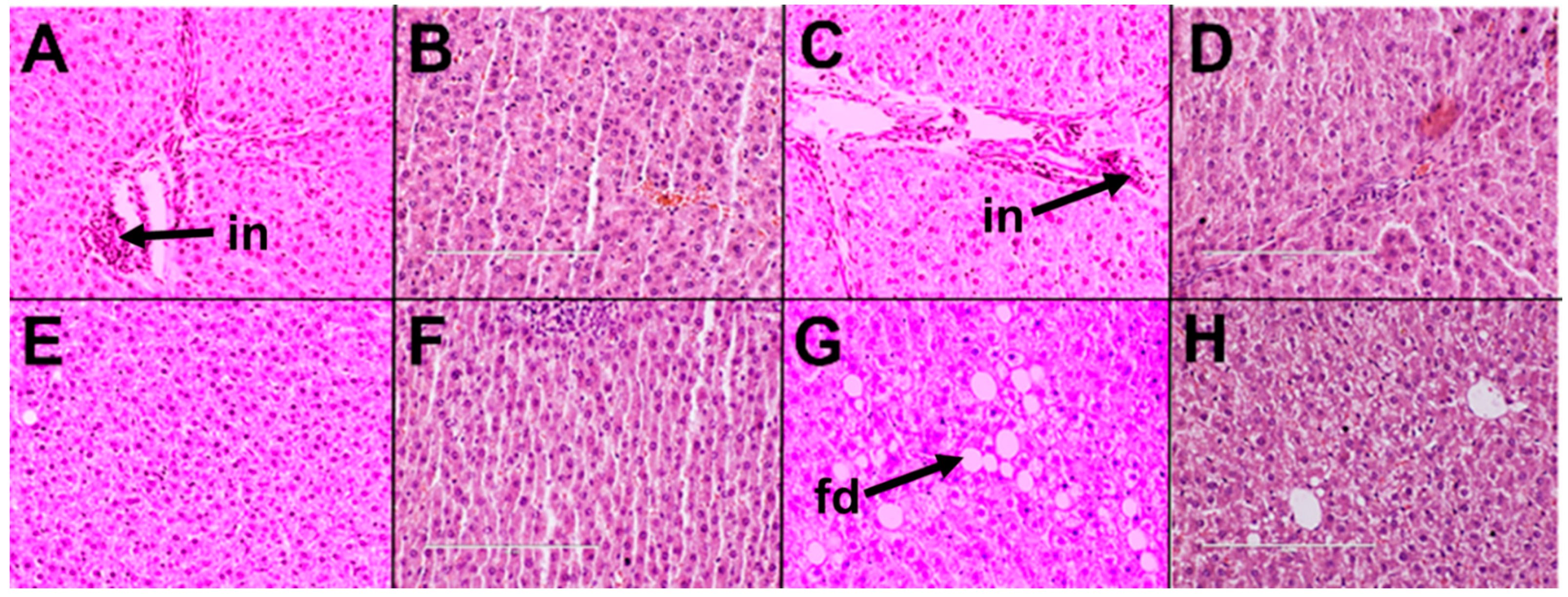
2.1.2. Cardiovascular and Liver Function
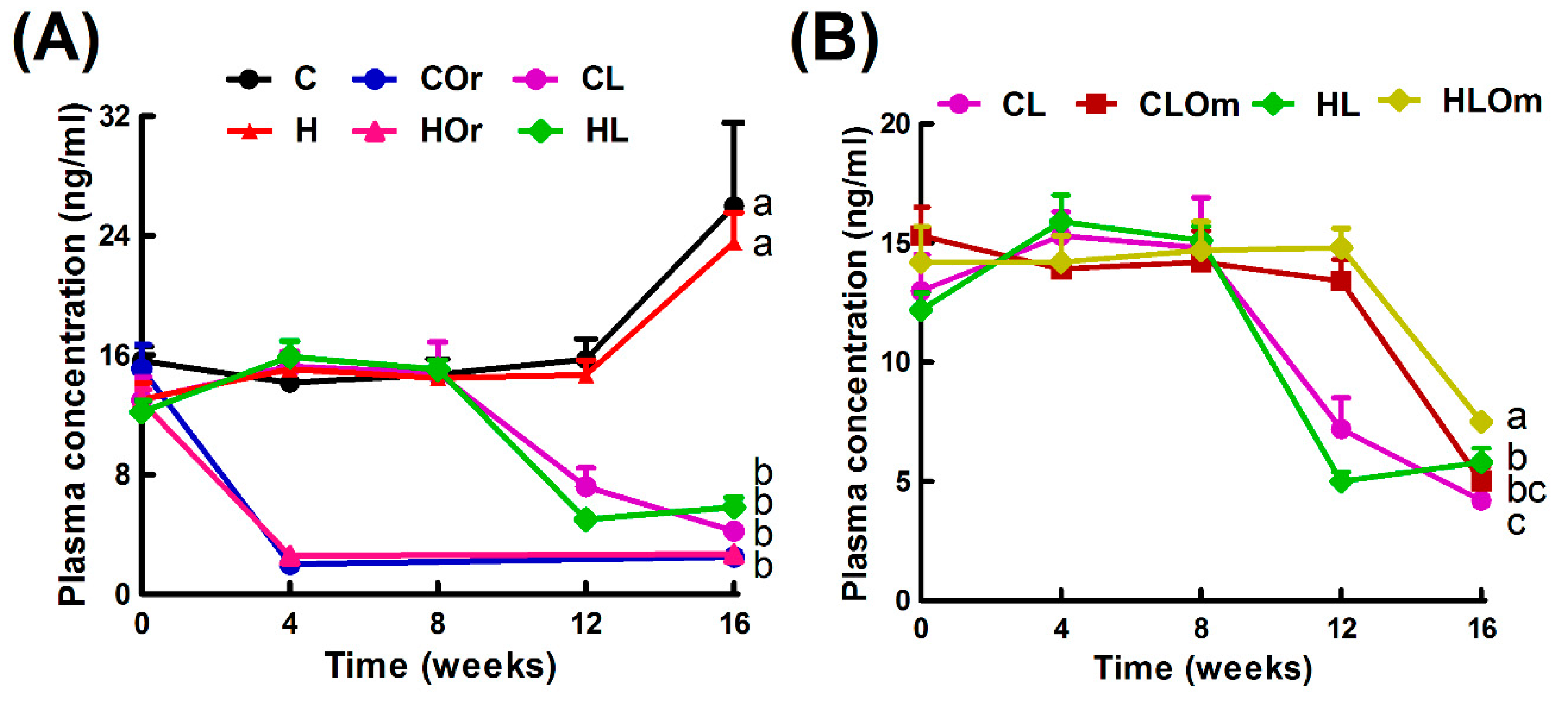
2.2. Bilateral Orchidectomy
2.2.1. Dietary Intakes, Body Composition, and Metabolic Parameters
2.2.2. Cardiovascular and Liver Function
2.3. Leuprolide Treatment
2.3.1. Dietary Intakes, Body Composition, and Metabolic Parameters
2.3.2. Cardiovascular and Liver Function
2.4. Omacor Treatment
2.4.1. Dietary Intakes, Body Composition, and Metabolic Parameters
2.4.2. Cardiovascular and Liver Function
3. Discussion
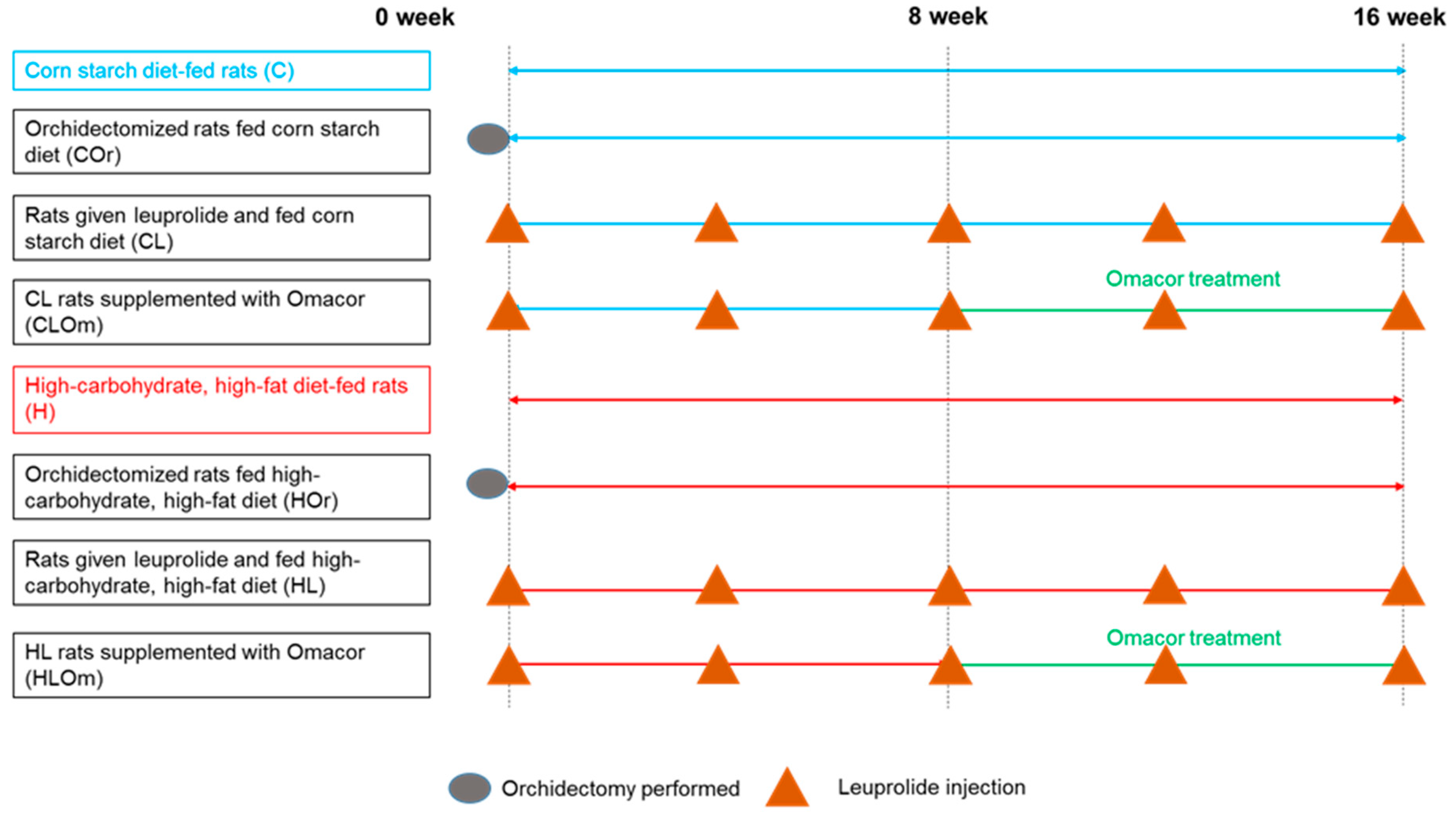
4. Materials and Methods
4.1. Rats and Diets
- C: Corn starch diet-fed rats for 16 weeks
- COr: Orchidectomized rats fed corn starch diet for 16 weeks
- CL: Rats treated with leuprolide and fed corn starch diet for 16 weeks
- CLOm: Rats treated with leuprolide for 16 weeks and fed corn starch diet for first 8 weeks followed by corn starch diet supplemented with 3% Omacor for the final 8 weeks
- H: High-carbohydrate, high-fat diet-fed rats for 16 weeks
- HOr: Orchidectomized rats fed high-carbohydrate, high-fat diet for 16 weeks
- HL: Rats treated with leuprolide and fed high-carbohydrate, high-fat diet for 16 weeks
- HLOm: Rats treated with leuprolide for 16 weeks and fed high-carbohydrate, high-fat diet for first 8 weeks followed by high-carbohydrate, high-fat diet supplemented with 3% Omacor for the final 8 weeks
4.2. Induction of Testosterone Deficiency
4.3. Rats, Diets, and Treatments
4.4. Physiological Parameters
4.5. Systolic Blood Pressure Measurements
4.6. Echocardiography
4.7. Body Composition Measurement
4.8. Oral Glucose Tolerance Test
4.9. Terminal Experiments
4.10. Left Ventricular Function
4.11. Vascular Reactivity
4.12. Organ Weights
4.13. Histology
4.14. Plasma Biochemistry
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Neill, S.; O’Driscoll, L. Metabolic syndrome: A closer look at the growing epidemic and its associated pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, H.; Li, Y.; Li, S.; Fernandez, C.; Bazzano, L.; He, J.; Xue, F.; Chen, W. Long-term impact of temporal sequence from childhood obesity to hyperinsulinemia on adult metabolic syndrome and diabetes: The Bogalusa Heart Study. Sci. Rep. 2017, 7, 43422. [Google Scholar] [CrossRef] [PubMed]
- Dockery, F.; Bulpitt, C.J.; Agarwal, S.; Donaldson, M.; Rajkumar, C. Testosterone suppression in men with prostate cancer leads to an increase in arterial stiffness and hyperinsulinaemia. Clin. Sci. 2003, 104, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Haffner, S.M.; Mykkanen, L.; Valdez, R.A.; Katz, M.S. Relationship of sex hormones to lipids and lipoproteins in nondiabetic men. J. Clin. Endocrinol. Metab. 1993, 77, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Charles, M.A.; Nahoul, K.; Orssaud, G.; Kremski, J.; Hully, V.; Joubert, E.; Papoz, L.; Eschwege, E. Association between plasma total testosterone and cardiovascular risk factors in healthy adult men: The Telecom Study. J. Clin. Endocrinol. Metab. 1997, 82, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Stellato, R.K.; Feldman, H.A.; Hamdy, O.; Horton, E.S.; McKinlay, J.B. Testosterone, sex hormone-binding globulin, and the development of type 2 diabetes in middle-aged men: Prospective results from the Massachusetts Male Aging Study. Diabetes Care 2000, 23, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, S.; Tenaglia, R.L. Metabolic syndrome and aggressive prostate cancer at initial diagnosis. Horm. Metab. Res. 2017, 49, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Gacci, M.; Russo, G.I.; De Nunzio, C.; Sebastianelli, A.; Salvi, M.; Vignozzi, L.; Tubaro, A.; Morgia, G.; Serni, S. Meta-analysis of metabolic syndrome and prostate cancer. Prostate Cancer Prostatic Dis. 2017, 20, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Lepor, H.; Shore, N.D. LHRH agonists for the treatment of prostate cancer: 2012. Rev. Urol. 2012, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, S.L.; Koupparis, A.; Robinson, M.E. Differing levels of testosterone and the prostate: A physiological interplay. Nat. Rev. Urol. 2011, 8, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; O’Malley, A.J.; Keating, N.L. Gonadotrophin-releasing hormone agonists, diabetes and cardiovascular disease in men with prostate cancer: Which metabolic syndrome? BJU Int. 2008, 101, 1335–1336. [Google Scholar] [CrossRef] [PubMed]
- van Londen, G.J.; Levy, M.E.; Perera, S.; Nelson, J.B.; Greenspan, S.L. Body composition changes during androgen deprivation therapy for prostate cancer: A 2-year prospective study. Crit. Rev. Oncol. Hematol. 2008, 68, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Bennacer, A.F.; Haffaf, E.; Kacimi, G.; Oudjit, B.; Koceir, E.A. Association of polyunsaturated/saturated fatty acids to metabolic syndrome cardiovascular risk factors and lipoprotein (a) in hypertensive type 2 diabetic patients. Ann. Biol. Clin. 2017, 75, 293–304. [Google Scholar] [CrossRef]
- Guo, X.F.; Li, X.; Shi, M.; Li, D. n-3 polyunsaturated fatty acids and metabolic syndrome risk: A meta-analysis. Nutrients 2017, 9, 703. [Google Scholar] [CrossRef] [PubMed]
- Tortosa-Caparros, E.; Navas-Carrillo, D.; Marin, F.; Orenes-Pinero, E. Anti-inflammatory effects of omega 3 and omega 6 polyunsaturated fatty acids in cardiovascular disease and metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2017, 57, 3421–3429. [Google Scholar] [CrossRef] [PubMed]
- Bouwens, M.; van de Rest, O.; Dellschaft, N.; Bromhaar, M.G.; de Groot, L.C.; Geleijnse, J.M.; Muller, M.; Afman, L.A. Fish-oil supplementation induces antiinflammatory gene expression profiles in human blood mononuclear cells. Am. J. Clin. Nutr. 2009, 90, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Beilin, L.J. Omega-3 fatty acids and inflammation. Curr. Atheroscler. Rep. 2004, 6, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Dasilva, G.; Pazos, M.; García-Egido, E.; Pérez-Jiménez, J.; Torres, J.L.; Giralt, M.; Nogués, M.-R.; Medina, I. Lipidomics to analyze the influence of diets with different EPA:DHA ratios in the progression of Metabolic Syndrome using SHROB rats as a model. Food Chem. 2016, 205, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S.; Khaza’ai, H.; Patimah, I.; Rahmat, A.; Abed, Y. Effect of long chain omega-3 polyunsaturated fatty acids on inflammation and metabolic markers in hypertensive and/or diabetic obese adults: A randomized controlled trial. Food Nutr. Res. 2016, 60, 29268. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Renehan, A.G.; Zwahlen, M.; Minder, C.; T O’Dwyer, S.; Shalet, S.M.; Egger, M. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: Systematic review and meta-regression analysis. Lancet 2004, 363, 1346–1353. [Google Scholar] [CrossRef]
- You, D.; Jeong, I.G.; Kim, S.W.; Chung, B.H.; Cho, J.S.; Lee, H.M.; Yun, S.C.; Kim, C.S. Impacts of leuprolide acetate on quality of life in patients with prostate cancer: A prospective multicenter study. Scand. J. Urol. Nephrol. 2010, 44, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, C.N.; Petrylak, D.P.; Madan, R.A.; Parker, C. Progress in the treatment of advanced prostate cancer. Am. Soc. Clin. Oncol. Educ. Book 2014, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Macleod, L.C.; Chery, L.J.; Hu, E.Y.; Zeliadt, S.B.; Holt, S.K.; Lin, D.W.; Porter, M.P.; Gore, J.L.; Wright, J.L. Metabolic syndrome, dyslipidemia and prostate cancer recurrence after primary surgery or radiation in a veterans cohort. Prostate Cancer Prostatic Dis. 2015, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Marberger, M.; Kaisary, A.V.; Shore, N.D.; Karlin, G.S.; Savulsky, C.; Mis, R.; Leuratti, C.; Germa, J.R. Effectiveness, pharmacokinetics, and safety of a new sustained-release leuprolide acetate 3.75-mg depot formulation for testosterone suppression in patients with prostate cancer: A Phase III, open-label, international multicenter study. Clin. Ther. 2010, 32, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Cakiroglu, B.; Hazar, A.I.; Eyyupoglu, S.E.; Can Balci, M.B.; Sinanoglu, O.; Tuzlali, P. The comparative evaluation of apoptosis produced by leuprolide or orchiectomy on rat prostate tissue. Arch. Ital. Urol. Androl. 2016, 87, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Gandaglia, G.; Sun, M.; Popa, I.; Schiffmann, J.; Abdollah, F.; Trinh, Q.D.; Saad, F.; Graefen, M.; Briganti, A.; Montorsi, F.; et al. The impact of androgen-deprivation therapy (ADT) on the risk of cardiovascular (CV) events in patients with non-metastatic prostate cancer: A population-based study. BJU Int. 2014, 114, E82–E89. [Google Scholar] [CrossRef] [PubMed]
- Sidaway, P. Prostate cancer: Intermediate risk: RT or ADT? Nat. Rev. Urol. 2015, 12, 300. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Finkelstein, J.S.; McGovern, F.J.; Zietman, A.L.; Fallon, M.A.; Schoenfeld, D.A.; Kantoff, P.W. Changes in body composition during androgen deprivation therapy for prostate cancer. J. Clin. Endocrinol. Metab. 2002, 87, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jackson, G.; Jones, T.H.; Matsumoto, A.M.; Nehra, A.; Perelman, M.A.; Swerdloff, R.S.; Traish, A.; Zitzmann, M.; Cunningham, G. Low testosterone associated with obesity and the metabolic syndrome contributes to sexual dysfunction and cardiovascular disease risk in men with type 2 diabetes. Diabetes Care 2011, 34, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Tunn, U.W. A 6-month depot formulation of leuprolide acetate is safe and effective in daily clinical practice: A non-interventional prospective study in 1273 patients. BMC Urol. 2011, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Iyer, A.; Brown, L. Evaluation of the chronic complications of diabetes in a high fructose diet in rats. Indian J. Biochem. Biophys. 2009, 46, 66–72. [Google Scholar] [PubMed]
- Thirunavukkarasu, V.; Anitha Nandhini, A.T.; Anuradha, C.V. Lipoic acid attenuates hypertension and improves insulin sensitivity, kallikrein activity and nitrite levels in high fructose-fed rats. J. Comp. Physiol. B 2004, 174, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Aller, E.E.; Abete, I.; Astrup, A.; Martinez, J.A.; van Baak, M.A. Starches, sugars and obesity. Nutrients 2011, 3, 341–369. [Google Scholar] [CrossRef] [PubMed]
- Damber, J.E.; Tammela, T.L.; Iversen, P.; Abrahamsson, P.A.; Boccon-Gibod, L.; Olesen, T.K.; van der Meulen, E.; Persson, B.E. The effect of baseline testosterone on the efficacy of degarelix and leuprolide: Further insights from a 12-month, comparative, phase III study in prostate cancer patients. Urology 2012, 80, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P. Abdominal obesity and cardiovascular disease: Is inflammation the missing link? Can. J. Cardiol. 2012, 28, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Cameron, I.; Alam, M.A.; Wang, J.; Brown, L. Endurance exercise in a rat model of metabolic syndrome. Can. J. Physiol. Pharmacol. 2012, 90, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional foods as potential therapeutic options for metabolic syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Panchal, S.K.; Diwan, V.; Brown, L. Omega-3 fatty acids and metabolic syndrome: Effects and emerging mechanisms of action. Prog. Lipid Res. 2011, 50, 372–387. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.D.; Howe, P.R. Anti-obesity effects of long-chain omega-3 polyunsaturated fatty acids. Obes. Rev. 2009, 10, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Panchal, S.K.; Ward, L.C.; Brown, L. Effects of ALA, EPA and DHA in high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. J. Nutr. Biochem. 2013, 24, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrian, S.; Costa, A.G.; Navas-Carretero, S.; Zabala, M.; Martinez, J.A.; Moreno-Aliaga, M.J. Role of omega-3 fatty acids in obesity, metabolic syndrome, and cardiovascular diseases: A review of the evidence. J. Physiol. Biochem. 2013, 69, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, M.; Sotoudeh, G.; Djalali, M.; Alvandi, E.; Eshraghian, M.; Sojoudi, F.; Koohdani, F. Dietary omega-3 polyunsaturated fatty acid intake modulates impact of Insertion/Deletion polymorphism of ApoB gene on obesity risk in type 2 diabetic patients. Nutrition 2016, 32, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- Skulas-Ray, A.C.; Kris-Etherton, P.M.; Harris, W.S.; Vanden Heuvel, J.P.; Wagner, P.R.; West, S.G. Dose-response effects of omega-3 fatty acids on triglycerides, inflammation, and endothelial function in healthy persons with moderate hypertriglyceridemia. Am. J. Clin. Nutr. 2011, 93, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Essential fatty acids: Biochemistry, physiology and pathology. Biotechnol. J. 2006, 1, 420–439. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.; Strauss, J.G.; Haemmerle, G.; Schoiswohl, G.; Birner-Gruenberger, R.; Riederer, M.; Lass, A.; Neuberger, G.; Eisenhaber, F.; Hermetter, A.; et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 2004, 306, 1383–1386. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Takahashi, M.; Ezaki, O. Fish oil feeding decreases mature sterol regulatory element-binding protein 1 (SREBP-1) by down-regulation of SREBP-1c mRNA in mouse liver. A possible mechanism for down-regulation of lipogenic enzyme mRNAs. J. Biol. Chem. 1999, 274, 25892–25898. [Google Scholar] [CrossRef] [PubMed]
- Okada, L.; Oliveira, C.P.; Stefano, J.T.; Nogueira, M.A.; Silva, I.; Cordeiro, F.B.; Alves, V.A.F.; Torrinhas, R.S.; Carrilho, F.J.; Puri, P.; et al. Omega-3 PUFA modulate lipogenesis, ER stress, and mitochondrial dysfunction markers in NASH—Proteomic and lipidomic insight. Clin. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids and non-alcoholic fatty liver disease: Evidence of efficacy and mechanism of action. Mol. Aspects. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids, hepatic lipid metabolism, and nonalcoholic fatty liver disease. Annu. Rev. Nutr. 2013, 33, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, Y.A.; Portois, L.; Malaisse, W.J. n−3 Fatty acids and the metabolic syndrome. Am. J. Clin. Nutr. 2006, 83, 1499S–1504S. [Google Scholar] [CrossRef] [PubMed]
- Morishita, M.; Tanaka, T.; Shida, T.; Takayama, K. Usefulness of colon targeted DHA and EPA as novel diabetes medications that promote intrinsic GLP-1 secretion. J. Control Release 2008, 132, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.M.; Melo, A.L.; Damasceno, N.R. The benefits of omega-3 supplementation depend on adiponectin basal level and adiponectin increase after the supplementation: A randomized clinical trial. Nutrition 2017, 34, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Borges, M.C.; Oliveira, I.O.; Freitas, D.F.; Horta, B.L.; Ong, K.K.; Gigante, D.P.; Barros, A.J.D. Obesity-induced hypoadiponectinaemia: The opposite influences of central and peripheral fat compartments. Int. J. Epidemiol. 2017, 46, 2044–2055. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Itoh, M.; Suganami, T.; Satoh, N.; Tanimoto-Koyama, K.; Yuan, X.; Tanaka, M.; Kawano, H.; Yano, T.; Aoe, S.; Takeya, M.; et al. Increased adiponectin secretion by highly purified eicosapentaenoic acid in rodent models of obesity and human obese subjects. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, E.; Rafraf, M.; Farzadi, L.; Asghari-Jafarabadi, M.; Sabour, S. Effects of omega-3 fatty acids supplementation on serum adiponectin levels and some metabolic risk factors in women with polycystic ovary syndrome. Asia Pac. J. Clin. Nutr. 2012, 21, 511–518. [Google Scholar] [PubMed]
- Neschen, S.; Morino, K.; Rossbacher, J.C.; Pongratz, R.L.; Cline, G.W.; Sono, S.; Gillum, M.; Shulman, G.I. Fish oil regulates adiponectin secretion by a peroxisome proliferator-activated receptor-γ-dependent mechanism in mice. Diabetes 2006, 55, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Bhaswant, M.; Poudyal, H.; Brown, L. Mechanisms of enhanced insulin secretion and sensitivity with n-3 unsaturated fatty acids. J. Nutr. Biochem. 2015, 26, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Aldiss, P.; Davies, G.; Woods, R.; Budge, H.; Sacks, H.S.; Symonds, M.E. ‘Browning’ the cardiac and peri-vascular adipose tissues to modulate cardiovascular risk. Int. J. Cardiol. 2017, 228, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Lepretti, M.; Martucciello, S.; Burgos Aceves, M.A.; Putti, R.; Lionetti, L. Omega-3 fatty acids and insulin resistance: Focus on the regulation of mitochondria and endoplasmic reticulum stress. Nutrients 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.C.; Battersby, K.J. Assessment of body composition of rats by bioimpedance spectroscopy: Validation against dual-energy X-ray absorptiometry. Scand. J. Lab. Anim. Sci. 2009, 36, 235–261. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | C | COr | CL | H | HOr | HL | p Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Diet | Testosterone Deficiency | Diet × Testosterone Deficiency | |||||||
| Initial body weight (g) | 337 ± 1 | 338 ± 1 | 337 ± 1 | 336 ± 1 | 339 ± 1 | 338 ± 1 | 0.68 | 0.14 | 0.52 |
| Final body weight (g) | 419 ± 5 b | 429 ± 5 b | 417 ± 8 b | 520 ± 6 a | 522 ± 14 a | 539 ± 18 a | <0.0001 | 0.71 | 0.37 |
| Body weight gain 9–16 weeks (%) | 5.3 ± 0.9 c | 11.0 ± 1.2 b | 11.2 ± 1.1 b | 19.3 ± 0.9 a | 17.6 ± 1.6 a | 20.4 ± 0.9 a | <0.0001 | 0.011 | 0.006 |
| Water intake (mL/day) | 31.0 ± 2.1 a | 32.0 ± 2.4 a | 26.4 ± 2.6 a,b | 19.8 ± 1.6 b,c | 19.8 ± 1.6 b,c | 17.5 ± 1.7 c | <0.0001 | 0.12 | 0.71 |
| Food intake (g/day) | 31.6 ± 2.0 a | 31.9 ± 2.3 a | 36.1 ± 2.2 a | 20.8 ± 1.3 b | 21.1 ± 1.8 b | 24.3 ± 1.9 b | <0.0001 | 0.08 | 0.96 |
| Energy intake (kJ/day) | 346 ± 3 d | 356 ± 3 d | 401 ± 6 c | 444 ± 3 b | 447 ± 5 b | 507 ± 12 a | <0.0001 | <0.0001 | 0.54 |
| Feed efficiency (g/kJ) | 0.23 ± 0.01 b | 0.25 ± 0.02 b | 0.20 ± 0.02 b | 0.41 ± 0.01 a | 0.40 ± 0.03 a | 0.40 ± 0.03 a | <0.0001 | 0.48 | 0.51 |
| Abdominal circumference (cm) | 18.4 ± 0.1 d | 20.1 ± 0.4 c | 20.1 ± 0.2 c | 22.0 ± 0.2 b | 23.2 ± 0.5 a | 23.0 ± 0.3 a | <0.0001 | <0.0001 | 0.52 |
| Basal blood glucose (mmol/L) | 4.1 ± 0.1 d | 4.7 ± 0.1 c | 4.8 ± 0.1 c | 5.1 ± 0.1 b,c | 5.6 ± 0.1 a | 5.4 ± 0.2 a,b | <0.0001 | <0.0001 | 0.24 |
| Area under the curve (mmol/L × minutes) | 632 ± 21 c | 776 ± 19 b | 781 ± 10 b | 774 ± 16 b | 854 ± 14 a | 844 ± 24 a | <0.0001 | <0.0001 | 0.07 |
| Total cholesterol (mmol/L) | 1.37 ± 0.05 b | 1.71 ± 0.15 a,b | 1.65 ± 0.04 a,b | 1.88 ± 0.06 a | 1.84 ± 0.17 a | 1.90 ± 0.10 a | 0.001 | 0.28 | 0.20 |
| Triglyceride (mmol/L) | 0.60 ± 0.03 c | 0.56 ± 0.05 c | 0.60 ± 0.06 c | 1.42 ± 0.15 b | 1.06 ± 0.22 b,c | 2.02 ± 0.32 a | <0.0001 | 0.019 | 0.032 |
| NEFA (mmol/L) | 1.74 ± 0.16 c | 2.47 ± 0.17 c | 2.38 ± 0.19 c | 3.69 ± 0.37 b | 3.41 ± 0.43 b | 4.60 ± 0.35 a | <0.0001 | 0.034 | 0.09 |
| Whole-body lean mass (g) | 312 ± 11 a | 271 ± 11 b | 275 ± 5 b | 314 ± 8 a | 229 ± 14 c | 308 ± 9 a | 0.78 | <0.0001 | 0.002 |
| Whole-body fat mass (g) | 114 ± 11 c | 121 ± 10 c | 112 ± 8 c | 191 ± 8 b | 259 ± 23 a | 161 ± 15 b,c | <0.0001 | 0.0007 | 0.006 |
| Bone mineral content (g) | 13.3 ± 0.5 b | 12.4 ± 0.4 b,c | 11.9 ± 0.2 b,c | 13.3 ± 0.4 b | 16.2 ± 0.6 a | 11.3 ± 0.5 c | 0.005 | <0.0001 | <0.0001 |
| Bone mineral density (g/cm2) | 0.183 ± 0.003 | 0.173 ± 0.004 | 0.174 ± 0.003 | 0.183 ± 0.004 | 0.173 ± 0.003 | 0.177 ± 0.003 | 0.72 | 0.12 | 0.88 |
| Retroperitoneal fat (mg/mm) | 122 ± 8 d | 231 ± 18 c | 220 ± 18 c | 366 ± 17 b | 537 ± 26 a | 479 ± 54 a | <0.0001 | <0.0001 | 0.51 |
| Epididymal fat (mg/mm) | 101 ± 4 b | - | 125 ± 11 b | 224 ± 17 a | - | 263 ± 26 a | <0.0001 | 0.07 | 0.65 |
| Omental fat (mg/mm) | 65 ± 6 b | 90 ± 6 b | 83 ± 6 b | 132 ± 9 a | 137 ± 13 a | 147 ± 13 a | <0.0001 | 0.16 | 0.52 |
| Variables | C | COr | CL | H | HOr | HL | p Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Diet | Testosterone Deficiency | Diet × Testosterone Deficiency | |||||||
| Heart rate (bpm) | 253 ± 23 b,c | 227 ± 17 c | 238 ± 11 b,c | 350 ± 10 a | 282 ± 15 b,c | 290 ± 15 b | <0.0001 | 0.01 | 0.29 |
| LVIDd (mm) | 6.64 ± 0.28 c | 6.63 ± 0.30 c | 7.70 ± 0.15 a,b | 7.09 ± 0.22 b,c | 7.23 ± 0.20 b,c | 8.24 ± 0.22 a | 0.007 | <0.0001 | 0.95 |
| LVIDs (mm) | 3.11 ± 0.19 c | 3.68 ± 0.18 b,c | 4.52 ± 0.19 a | 3.22 ± 0.23 c | 3.48 ± 0.25 b,c | 4.14 ± 0.18 a,b | 0.35 | <0.0001 | 0.49 |
| IVSd (mm) | 1.91 ± 0.12 | 1.96 ± 0.10 | 1.77 ± 0.06 | 2.05 ± 0.08 | 2.05 ± 0.17 | 1.82 ± 0.06 | 0.28 | 0.10 | 0.91 |
| IVSs (mm) | 3.09 ± 0.26 a,b | 3.13 ± 0.20 a,b | 2.80 ± 0.08 b | 3.65 ± 0.09 a | 3.43 ± 0.20 a,b | 3.12 ± 0.11 a,b | 0.006 | 0.047 | 0.70 |
| LVPWd (mm) | 1.70 ± 0.09 | 1.91 ± 0.12 | 1.79 ± 0.03 | 2.04 ± 0.06 | 2.08 ± 0.15 | 1.75 ± 0.05 | 0.05 | 0.06 | 0.14 |
| LVPWs (mm) | 2.73 ± 0.16 b | 2.65 ± 0.14 b | 2.58 ± 0.15 b | 3.27 ± 0.14 a | 3.04 ± 0.13 a,b | 2.95 ± 0.10 a,b | 0.0003 | 0.23 | 0.80 |
| Diastolic volume (µL) | 317 ± 41 c | 318 ± 38 c | 481 ± 26 a,b | 427 ± 40 a,b,c | 356 ± 30 b,c | 531 ± 47 a | 0.036 | 0.0001 | 0.59 |
| Systolic volume (µL) | 34.2 ± 6.1 c | 55.0 ± 8.2 b,c | 100.2 ± 12.8 a | 38.4 ± 7.0 c | 49.2 ± 10.5 b,c | 77.1 ± 8.6 a,b | 0.28 | <0.0001 | 0.33 |
| Stroke volume (µL) | 283 ± 38 b | 263 ± 33 b | 381 ± 22 a,b | 388 ± 35 a,b | 306 ± 24 b | 454 ± 43 a | 0.009 | 0.0007 | 0.65 |
| SBP:LVIDs | 42.4 ± 2.9 a,b | 40.9 ± 1.8 a,b,c | 32.1 ± 1.5 c | 48.3 ± 3.8 a | 48.0 ± 3.0 a | 36.9 ± 2.4 b,c | 0.009 | 0.0001 | 0.91 |
| SBP:systolic volume | 4826 ± 956 a,b | 3016 ± 418 a,b | 1599 ± 198 b | 5562 ± 1462 a | 4436 ± 840 a,b | 2291 ± 479 a,b | 0.17 | 0.001 | 0.89 |
| ESS:LVIDs | 2.40 ± 0.10 b | 2.44 ± 0.08 a,b | 2.84 ± 0.16 a | 2.30 ± 0.09 b | 2.69 ± 0.09 a,b | 2.56 ± 0.11 a,b | 0.63 | 0.007 | 0.05 |
| Cardiac output (mL/min) | 70.1 ± 9.2 b | 56.3 ± 7.9 b | 90.6 ± 7.3 b | 135.2 ± 10.9 a | 84.8 ± 5.6 b | 130.9 ± 12.9 a | <0.0001 | 0.0001 | 0.14 |
| Relative wall thickness | 0.56 ± 0.05 | 0.58 ± 0.05 | 0.47 ± 0.02 | 0.56 ± 0.02 | 0.60 ± 0.05 | 0.45 ± 0.02 | 1.00 | 0.003 | 0.87 |
| Systolic wall stress | 75.7 ± 7.0 b | 104.4 ± 8.9 a,b | 129.9 ± 12.2 a | 74.8 ± 7.1 b | 93.3 ± 6.6 b | 106.4 ± 7.3 a,b | 0.09 | <0.0001 | 0.41 |
| Estimated LV mass (g) | 0.81 ± 0.03 b | 0.89 ± 0.05 a,b | 1.01 ± 0.04 a,b | 1.16 ± 0.07 a | 1.08 ± 0.12 a,b | 1.07 ± 0.06 a,b | 0.006 | 0.65 | 0.11 |
| Fractional shortening (%) | 53.1 ± 2.3 a,b | 47.1 ± 2.1 b,c | 41.2 ± 2.1 c | 56.6 ± 2.5 a | 49.6 ± 2.7 a,b,c | 47.7 ± 2.3 b,c | 0.033 | 0.0002 | 0.68 |
| Ejection fraction (%) | 89.1 ± 1.6 a | 84.0 ± 2.0 a,b | 79.3 ± 2.0 b | 91.2 ± 1.5 a | 86.6 ± 2.1 a | 85.2 ± 1.7 a,b | 0.021 | 0.0002 | 0.53 |
| Systolic blood pressure (mmHg) | 126 ± 2 d | 146 ± 1 b,c | 144 ± 2 c | 152 ± 2 b | 161 ± 2 a | 150 ± 2 b,c | <0.0001 | <0.0001 | <0.0001 |
| Right ventricular wet weight (mg/mm) | 2.37 ± 0.19 | 3.39 ± 0.64 | 2.76 ± 0.16 | 4.21 ± 1.14 | 2.88 ± 0.12 | 3.19 ± 0.15 | 0.20 | 0.85 | 0.11 |
| Left ventricular + septum wet weight (mg/mm) | 17.6 ± 0.5 b | 18.1 ± 0.7 a,b | 18.1 ± 0.5 a,b | 19.5 ± 0.5 a,b | 19.9 ± 0.2 a | 19.1 ± 0.5 a,b | 0.003 | 0.62 | 0.62 |
| Left ventricular diastolic stiffness constant | 23.8 ± 0.7 b | 29.4 ± 0.6 a | 27.3 ± 0.5 a | 29.1 ± 0.7 a | 29.1 ± 1.1 a | 27.9 ± 0.7 a | 0.003 | 0.002 | 0.0006 |
| Liver wet weight (mg/mm) | 241 ± 7 b | 239 ± 7 b | 250 ± 15 b | 333 ± 8 a | 351 ± 11 a | 325 ± 19 a | <0.0001 | 0.76 | 0.31 |
| ALT activity (U/L) | 25.9 ± 1.6 b | 29.0 ± 2.6 b | 27.6 ± 1.9 b | 41.8 ± 2.0 a | 39.6 ± 1.6 a | 40.4 ± 5.9 a | <0.0001 | 0.99 | 0.68 |
| AST activity (U/L) | 64.3 ± 2.9 b | 78.3 ± 5.5 a,b | 64.1 ± 3.5 b | 88.3 ± 3.6 a | 95.1 ± 4.3 a | 86.7 ± 9.4 a | <0.0001 | 0.07 | 0.78 |
| Variables | CL | CLOm | HL | HLOm | p Value | ||
|---|---|---|---|---|---|---|---|
| Diet + Leuprolide | Omacor | (Diet + Leuprolide) × Omacor | |||||
| Initial body weight (g) | 337 ± 1 | 342 ± 2 | 338 ± 1 | 339 ± 3 | 0.61 | 0.13 | 0.31 |
| Final body weight (g) | 417 ± 8 b | 319 ± 9 c | 539 ± 18 a | 536 ± 19 a | <0.0001 | 0.001 | 0.002 |
| Body weight gain 9–16 weeks (%) | 11.2 ± 1.1 c | −14.0 ± 1.4 d | 20.4 ± 0.9 a | 15.3 ± 1.5 b | <0.0001 | <0.0001 | <0.0001 |
| Water intake (mL/day) | 26.4 ± 2.6 b | 33.2 ± 2.6 a | 17.5 ± 1.7 c | 24.5 ± 1.1 b | 0.0001 | 0.002 | 0.96 |
| Food intake (g/day) | 36.1 ± 2.2 a | 38.3 ± 2.8 a | 24.3 ± 1.9 b | 26.1 ± 1.1 b | <0.0001 | 0.34 | 0.93 |
| Energy intake (kJ/day) | 401 ± 6 c | 487 ± 34 b | 507 ± 12 a,b | 560 ± 21 a | 0.0001 | 0.002 | 0.44 |
| Feed efficiency (g/kJ) | 0.20 ± 0.02 b | −0.03 ± 0.01 c | 0.40 ± 0.03 a | 0.35 ± 0.03 a | <0.0001 | <0.0001 | 0.0005 |
| Abdominal circumference (cm) | 20.1 ± 0.2 b | 17.0 ± 0.2 c | 23.0 ± 0.4 a | 22.0 ± 0.4 a | <0.0001 | <0.0001 | 0.002 |
| Basal blood glucose (mmol/L) | 4.8 ± 0.1 b | 3.5 ± 0.1 c | 5.4 ± 0.2 a | 4.5 ± 0.3 b | 0.0002 | <0.0001 | 0.31 |
| Area under the curve (mmol/L × minutes) | 781 ± 10 b | 599 ± 16 c | 844 ± 24 b | 933 ± 40 a | <0.0001 | 0.07 | <0.0001 |
| Total cholesterol (mmol/L) | 1.65 ± 0.05 b | 1.04 ± 0.03 d | 1.89 ± 0.11 a | 1.41 ± 0.07 c | 0.0001 | <0.0001 | 0.37 |
| Triglyceride (mmol/L) | 0.60 ± 0.06 b | 0.27 ± 0.02 b | 1.98 ± 0.31 a | 0.33 ± 0.04 b | <0.0001 | <0.0001 | 0.0004 |
| NEFA (mmol/L) | 2.38 ± 0.19 b | 0.46 ± 0.03 c | 4.63 ± 0.43 a | 0.99 ± 0.18 c | <0.0001 | <0.0001 | 0.0016 |
| Whole-body lean mass (g) | 314 ± 8 a | 261 ± 10 b | 308 ± 9 a | 251 ± 13 b | 0.44 | <0.0001 | 0.85 |
| Whole-body fat mass (g) | 191 ± 8 b | 59 ± 8 c | 161 ± 15 b | 248 ± 27 a | <0.0001 | 0.18 | <0.0001 |
| Bone mineral content (g) | 0.173 ± 0.003 | 0.179 ± 0.003 | 0.177 ± 0.004 | 0.180 ± 0.001 | 0.40 | 0.14 | 0.62 |
| Bone mineral density (g/cm2) | 12.0 ± 0.2 b | 10.6 ± 0.5 b | 11.3 ± 0.5 b | 16.8 ± 0.9 a | <0.0001 | 0.001 | <0.0001 |
| Retroperitoneal fat (mg/mm) | 220 ± 18 b | 95 ± 14 c | 479 ± 54 a | 406 ± 50 a | <0.0001 | 0.015 | 0.50 |
| Epididymal fat (mg/mm) | 125 ± 11 b | 67 ± 7 c | 263 ± 26 a | 226 ± 26 a | <0.0001 | 0.02 | 0.59 |
| Omental fat (mg/mm) | 83 ± 6 c | 79 ± 10 c | 147 ± 13 b | 249 ± 20 a | <0.0001 | 0.0007 | 0.0003 |
| Variables | CL | CLOm | HL | HLOm | p Value | ||
|---|---|---|---|---|---|---|---|
| Diet + Leuprolide | Omacor | (Diet + Leuprolide) × Omacor | |||||
| Heart rate | 238 ± 11 | 255 ± 22 | 290 ± 15 | 283 ± 11 | 0.08 | 0.75 | 0.44 |
| LVIDd (mm) | 7.70 ± 0.15 b | 7.64 ± 0.17 b | 8.24 ± 0.22 a,b | 8.54 ± 0.16 a | 0.0002 | 0.50 | 0.32 |
| LVIDs (mm) | 4.52 ± 0.19 b | 4.62 ± 0.13 b | 4.14 ± 0.18 b | 5.33 ± 0.09 a | 0.29 | 0.0001 | 0.0009 |
| IVSd (mm) | 1.77 ± 0.06 a,b | 1.68 ± 0.05 b | 1.82 ± 0.06 a,b | 1.91 ± 0.02 a | 0.008 | 1.0000 | 0.08 |
| IVSs (mm) | 2.80 ± 0.08 b | 2.72 ± 0.07 b | 3.12 ± 0.11 a | 2.92 ± 0.06 a,b | 0.0028 | 0.10 | 0.47 |
| LVPWd (mm) | 1.79 ± 0.03 a,b | 1.67 ± 0.03 b | 1.75 ± 0.05 a,b | 1.84 ± 0.04 a | 0.10 | 0.70 | 0.009 |
| LVPWs (mm) | 2.58 ± 0.15 b | 2.39 ± 0.05 b | 2.95 ± 0.10 a | 2.65 ± 0.06 b | 0.003 | 0.017 | 0.58 |
| Diastolic volume (µL) | 481 ± 26 b | 473 ± 34 b | 531 ± 47 b | 658 ± 36 a | 0.002 | 0.11 | 0.07 |
| Systolic volume (µL) | 100.0 ± 12.8 b | 104.9 ± 8.9 b | 77.0 ± 8.6 b | 159.2 ± 7.8 a | 0.12 | <0.0001 | 0.0003 |
| Stroke volume (µL) | 381 ± 22 b | 368 ± 25 b | 454 ± 43 a,b | 499 ± 31 a | 0.002 | 0.61 | 0.36 |
| SBP:LVIDs | 32.1 ± 1.5 b | 26.5 ± 0.9 c | 36.9 ± 2.4 a | 25.9 ± 0.7 c | 0.18 | <0.0001 | 0.08 |
| SBP:systolic volume | 1599 ± 198 a,b | 1216 ± 101 b | 2291 ± 479 a | 882 ± 48 b | 0.50 | 0.002 | 0.06 |
| ESS:LVIDs | 2.84 ± 0.16 | 2.55 ± 0.07 | 2.56 ± 0.11 | 2.61 ± 0.08 | 0.33 | 0.28 | 0.13 |
| Cardiac output (mL) | 91 ± 7 b | 95 ± 11 b | 131 ± 13 a | 141 ± 10 a | 0.0001 | 0.50 | 0.76 |
| Relative wall thickness | 0.47 ± 0.02 | 0.44 ± 0.01 | 0.45 ± 0.02 | 0.44 ± 0.01 | 0.53 | 0.21 | 0.53 |
| Systolic wall stress | 129.9 ± 12.2 a,b | 117.8 ± 4.0 a,b | 106.4 ± 7.3 b | 139.0 ± 4.0 a | 0.88 | 0.19 | 0.006 |
| Estimated LV mass (g) | 1.01 ± 0.04 b | 0.92 ± 0.05 b | 1.07 ± 0.06 b | 1.29 ± 0.05 a | 0.0001 | 0.21 | 0.004 |
| Fractional shortening (%) | 41.2 ± 2.1 b | 39.6 ± 0.7 b | 47.7 ± 2.3 a | 37.6 ± 0.8 b | 0.18 | 0.0009 | 0.013 |
| Ejection fraction (%) | 79.3 ± 2.0 b | 77.9 ± 0.7 b | 85.2 ± 1.7 a | 75.6 ± 0.9 b | 0.22 | 0.0004 | 0.006 |
| Systolic blood pressure (mmHg) | 144 ± 2 b | 122 ± 1 d | 150 ± 2 a | 137 ± 2 c | <0.0001 | <0.0001 | 0.016 |
| Right ventricular wet weight (mg/mm) | 2.76 ± 0.16 c | 3.59 ± 0.29 b | 3.19 ± 0.15 b,c | 4.71 ± 0.22 a | 0.0008 | <0.0001 | 0.11 |
| Left ventricular + septum wet weight (mg/mm) | 18.1 ± 0.5 b | 17.2 ± 0.8 b | 19.1 ± 0.5 b | 22.9 ± 0.9 a | <0.0001 | 0.044 | 0.002 |
| Left ventricular diastolic stiffness constant | 27.3 ± 0.5 a | 22.8 ± 1.0 bc | 27.9 ± 0.7 a | 21.3 ± 1.0 c | 0.59 | <0.0001 | 0.21 |
| Liver wet weight (mg/mm) | 250 ± 15 b | 161 ± 7 c | 325 ± 19 a | 294 ± 14 a | <0.0001 | 0.0001 | 0.05 |
| ALT activity (U/L) | 27.6 ± 1.9 b | 41.4 ± 4.4 a | 34.9 ± 2.4 ab | 39.7 ± 2.8 a | 0.36 | 0.004 | 0.15 |
| AST activity (U/L) | 64.1 ± 3.5 b | 72.5 ± 3.1 ab | 86.7 ± 9.4 a | 70.0 ± 4.2 ab | 0.08 | 0.47 | 0.032 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhandarkar, N.S.; Kumar, S.A.; Martin, J.; Brown, L.; Panchal, S.K. Attenuation of Metabolic Syndrome by EPA/DHA Ethyl Esters in Testosterone-Deficient Obese Rats. Mar. Drugs 2018, 16, 182. https://doi.org/10.3390/md16060182
Bhandarkar NS, Kumar SA, Martin J, Brown L, Panchal SK. Attenuation of Metabolic Syndrome by EPA/DHA Ethyl Esters in Testosterone-Deficient Obese Rats. Marine Drugs. 2018; 16(6):182. https://doi.org/10.3390/md16060182
Chicago/Turabian StyleBhandarkar, Nikhil S., Senthil Arun Kumar, Jarad Martin, Lindsay Brown, and Sunil K. Panchal. 2018. "Attenuation of Metabolic Syndrome by EPA/DHA Ethyl Esters in Testosterone-Deficient Obese Rats" Marine Drugs 16, no. 6: 182. https://doi.org/10.3390/md16060182
APA StyleBhandarkar, N. S., Kumar, S. A., Martin, J., Brown, L., & Panchal, S. K. (2018). Attenuation of Metabolic Syndrome by EPA/DHA Ethyl Esters in Testosterone-Deficient Obese Rats. Marine Drugs, 16(6), 182. https://doi.org/10.3390/md16060182