Marine Myxobacteria: A Few Good Halophiles


Abstract
:1. Introduction
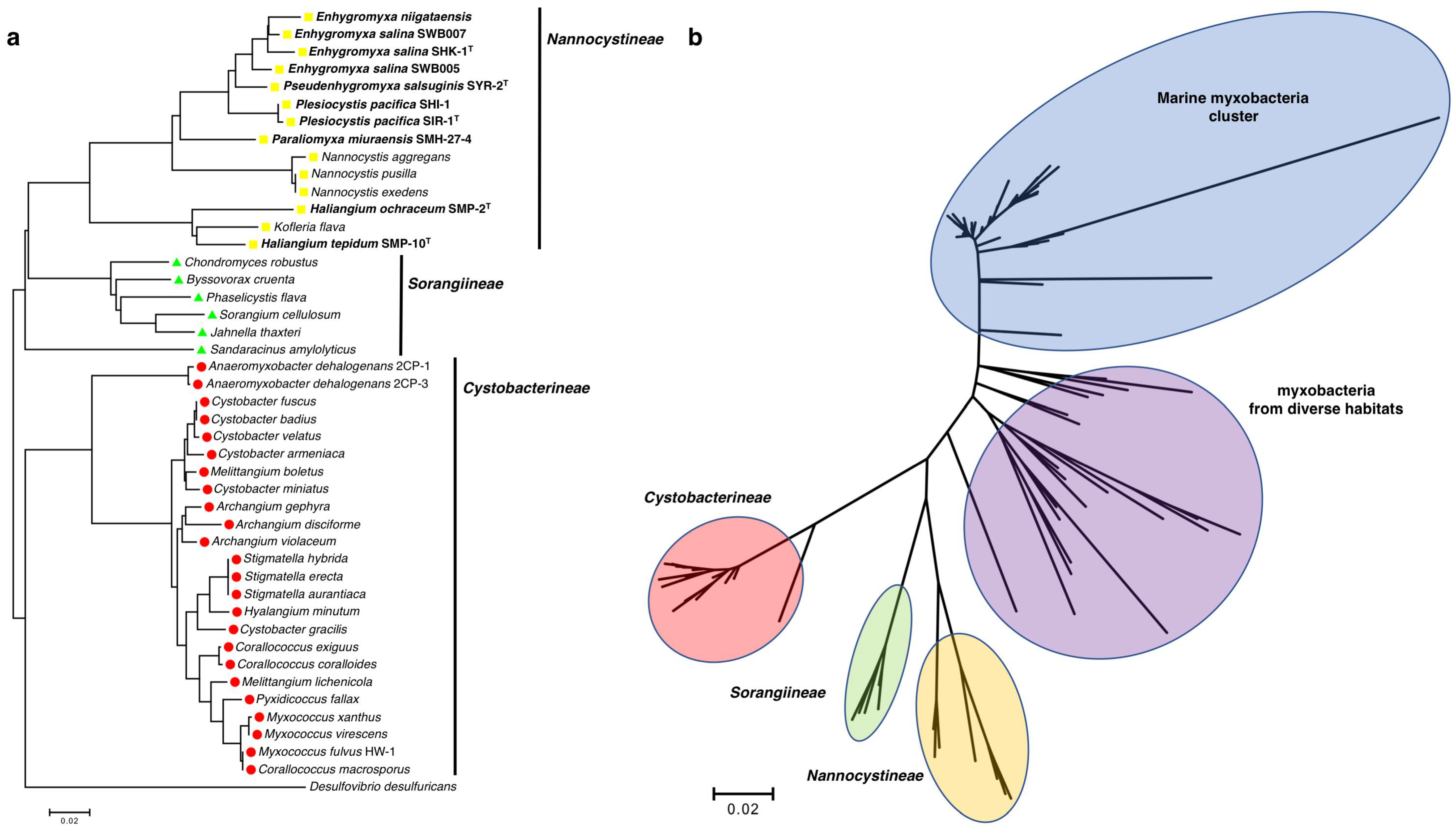
2. Characteristics Unique to Marine Myxobacteria
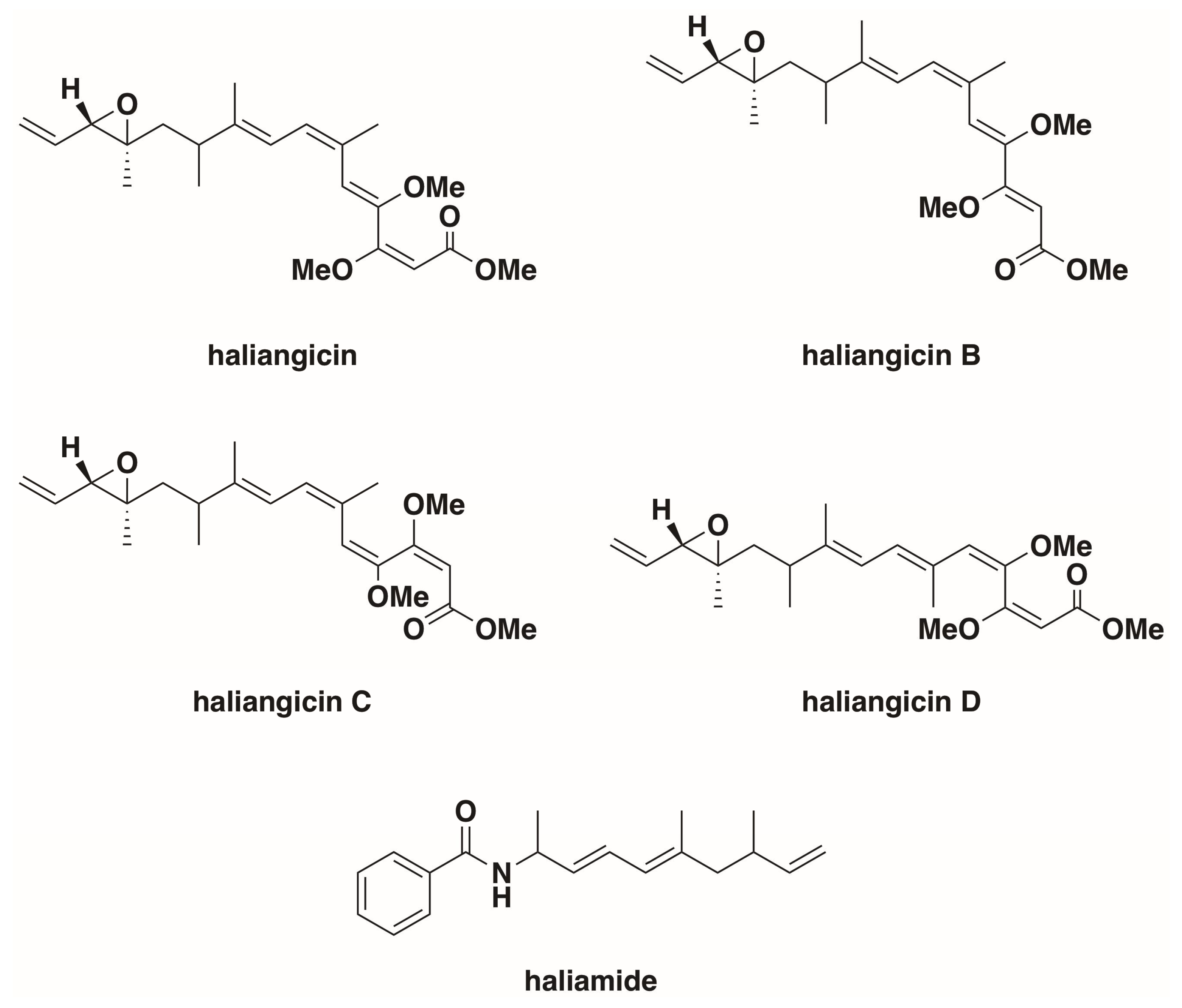
3. Haliangium ochraceum
4. Enhygromyxa salina
5. Plesiocystis pacifica
6. Haliangium tepidum
7. Potential Marine Myxobacteria
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reichenbach, H. The ecology of the myxobacteria. Environ. Microbiol. 1999, 1, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanier, R.Y. A Note on the Elasticotaxis in Myxobacteria. J. Bacteriol. 1942, 44, 405–412. [Google Scholar] [PubMed]
- Stanier, R.Y. Studies on nonfruiting myxobacteria. I. Cytophaga johnsonae n. sp., a chitin-decomposing myxobacterium. J. Bacteriol. 1947, 53, 297–315. [Google Scholar] [PubMed]
- Veldkamp, H. A Study on Two Marine Agar-Decomposing, Facultatively Anaerobic Myxobacteria. Microbiology 1961, 26, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, T.G.; Hendrie, M.S.; Shewan, J.M. The Taxonomy, Differentiation and Identification of Cytophaga Species. J. Appl. Bacteriol. 1969, 32, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Brockman, E.R. Fruiting myxobacteria from the South Carolina coast. J. Bacteriol. 1967, 94, 1253–1254. [Google Scholar] [PubMed]
- Rückert, G. Investigations on the distribution of myxobacteria in substrates influenced by seawater with special reference to the island of Helgoland. Helogländer Meeresuntersuchungen 1984, 38, 179–184. [Google Scholar] [CrossRef]
- Gray, J.P.; Herwig, R.P. Phylogenetic analysis of the bacterial communities in marine sediments. Appl. Environ. Microbiol. 1996, 62, 4049–4059. [Google Scholar] [PubMed]
- Ravenschlag, K.; Sahm, K.; Pernthaler, J.; Amann, R. High bacterial diversity in permanently cold marine sediments. Appl. Environ. Microbiol. 1999, 65, 3982–3989. [Google Scholar] [PubMed]
- Bernardet, J.F.; Segers, P.; Vancanneyt, M.; Berthe, F.; Kersters, K.; Vandamme, P. Cutting a Gordian Knot: Emended Classification and Description of the Genus Flavobacterium, Emended Description of the Family Flavobacteriaceae, and Proposal of Flavobacterium hydatis nom. nov. (Basonym, Cytophaga aquatilis Strohl and Tait 1978). Int. J. Syst. Bacteriol. 1996, 46, 128–148. [Google Scholar] [CrossRef]
- Moyer, C.L.; Dobbs, F.C.; Karl, D.M. Phylogenetic diversity of the bacterial community from a microbial mat at an active, hydrothermal vent system, Loihi Seamount, Hawaii. Appl. Environ. Microbiol. 1995, 61, 1555–1562. [Google Scholar] [PubMed]
- Iizuka, T.; Jojima, Y.; Fudou, R.; Yamanaka, S. Isolation of myxobacteria from the marine environment. FEMS Microbiol. Lett. 1998, 169, 317–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Q.; Li, Y.Z.; Wang, B.; Wu, Z.H.; Zhang, C.Y.; Gong, X.; Qui, Z.J.; Zhang, Y. Characteristics and living patterns of marine myxobacterial isolates. Appl. Environ. Microbiol. 2005, 71, 3331–3336. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.F.; Li, X.; Liu, H.; Liu, X.; Han, K.; Wu, Z.H.; Hu, W.; Li, F.F.; Li, Y.Z. Genome sequence of the halotolerant marine bacterium Myxococcus fulvus HW-1. J. Bacteriol. 2011, 193, 5015–5016. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boury-Esnault, N.; Brandão, S.N.; Costello, M.J.; Gofas, S.; Hernandez, F.; Mees, J.; et al. World Register of Marine Species; WoRMS Editorial Board: Ostend, Belgium, 2018; Available online: http://www.marinespecies.org (accessed on 1 May 2018). [CrossRef]
- Pye, C.R.; Bertin, M.J.; Lokey, R.S.; Gerwick, W.H.; Linington, R.G. Retrospective analysis of natural products provides insights for future discovery trends. Proc. Natl. Acad. Sci. USA 2017, 114, 5601–5606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, J.; Fayad, A.A.; Müller, R. Natural Products from myxobacteria: Novel metabolites and bioactivities. Nat. Prod. Rep. 2017, 34, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Dávila-Céspedes, A.; Hufendiek, P.; Crüsemann, M.; Schäberle, T.F.; König, G.M. Marine-derived myxobacteria of the suborder Nannocystineae: An underexplored source of structurally intriguing and biologically active metabolites. Beilstein J. Org. Chem. 2016, 12, 969–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäberle, T.F.; Lohr, F.; Schmitz, A.; König, G.M. Antibiotics from myxobacteria. Nat. Prod. Rep. 2014, 31, 953–972. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, M.S.; Santos, H.; Galinski, E.A. An overview of the role and diversity of compatible solute in Bacteria and Archaea. Adv. Biochem. Eng. Biotechnol. 1998, 61, 117–153. [Google Scholar] [PubMed]
- Burg, M.B.; Ferraris, J.D. Intracellular organic osmolytes: Function and regulation. J. Biol. Chem. 2008, 283, 7309–7313. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, J.A.; Boehringer, N.; Burdziak, A.; Kunte, H.; Galinski, E.A.; Schäberle, T.F. Different strategies of osmoadaptation in the closely related marine myxobacteria Enhygromyxa salina SWB007 and Plesiocystis pacifica SIR-1. Microbiology 2016, 162, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, D.; Søgaard-Anderson, L. Regulation of Cell Polarity in Motility and Cell Division in Myxococcus xanthus. Annu. Rev. Microbiol. 2017, 71, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Dorado, J.; Marcos-Torres, F.J.; Garćia-Bravo, E.; Moraleda-Muñoz, A.; Pérez, J. Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Front. Microbiol. 2016, 7, 781. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Hu, W.; Liu, H.; Zhang, C.Y.; Zhao, J.Y.; Jiang, D.M.; Wu, Z.H.; Li, Y.Z. Adaptation of Salt-tolerant Myxococcus Strains and their Motility Systems to the Ocean Conditions. Microb. Ecol. 2007, 54, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Jojima, Y.; Hayakawa, A.; Fujii, T.; Yamanaka, S.; Fudou, R. Pseudenhygromyxa salsuginis gen. nov., sp. nov., a myxobacterium isolated from an estuarine marsh. Int. J. Syst. Evol. Microbiol. 2013, 63, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Jojima, Y.; Fudou, R.; Tokura, M.; Hiraishi, A.; Yamanaka, S. Enhygromyxa salina gen. nov., sp. nov., a slightly halophilic myxobacterium isolated from the coastal areas of Japan. Syst. Appl. Microbiol. 2003, 26, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Jojima, Y.; Fudou, R.; Hiraishi, A.; Ahn, J.W.; Yamanaka, S. Plesiocystis pacifica gen. nov., sp. nov., a marine myxobacterium that contains dehydrogenated menaquinone, isolated from the Pacific coasts of Japan. Int. J. Syst. Evol. Microbiol. 2003, 53, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Fudou, R.; Jojima, Y.; Iizuka, T.; Yamanaka, S. Haliangium ochraceum gen. nov., sp. nov. and Haliangium tepidum sp. nov.: Novel moderately halophilic myxobacteria isolated from coastal saline environments. J. Gen. Appl. Microbiol. 2002, 48, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Schäberle, T.F.; Goralski, E.; Neu, E.; Erol, O.; Hölzl, G.; Dörmann, P.; Bierbaum, G.; König, G.M. Marine myxobacteria as a source of antibiotics—Comparison of physiology, polyketide-type genes and antibiotic production of three new isolates of Enhygromyxa salina. Mar. Drugs 2010, 8, 2466–2479. [Google Scholar] [CrossRef] [PubMed]
- Brinkhoff, T.; Fischer, D.; Vollmers, J.; Voget, S.; Beardsley, C.; Thole, S.; Mussmann, M.; Kunze, B.; Wagner-Döbler, I.; Daniel, R.; et al. Biogeography and phylogenetic diversity of a cluster of exclusively marine myxobacteria. ISME J. 2012, 6, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.M.; Kato, C.; Zhou, X.W.; Wu, Z.H.; Sato, T.; Li, Y.Z. Phylogeographic separation of marine and soil myxobacteria at high levels of classification. ISME J. 2010, 4, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, N.; Daum, C.; Lang, E.; Abt, B.; Kopitz, M.; Saunders, E.; Lapidus, A.; Lucas, S.; Glavina Del Rio, T.; Nolan, M.; et al. Complete genome sequence of Haliangium ochraceum type strain (SMP-2). Stand. Genom. Sci. 2010, 2, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Fudou, R.; Iizuka, T.; Yamanaka, S. Haliangicin, a novel antifungal metabolite produced by a marine myxobacterium. 1. Fermentation and biological characteristics. J. Antibiot. (Tokyo) 2001, 54, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Fudou, R.; Iizuka, T.; Sato, S.; Ando, T.; Shimba, N.; Yamanaka, S. Haliangicin, a novel antifungal metabolite produced by a marine myxobacterium. 2. Isolation and structural elucidation. J. Antibiot. (Tokyo) 2001, 54, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Kundim, B.A.; Itou, Y.; Sakagami, Y.; Fudou, R.; Iizuka, T.; Yamanaka, S.; Ojika, M. New haliangicin isomers, potent antifungal metabolites produced by a marine myxobacterium. J. Antibiot. (Tokyo) 2003, 56, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Feng, Z.; Tomura, T.; Suzuki, A.; Miyano, S.; Tsuge, T.; Mori, H.; Suh, J.-W.; Iizuka, T.; Fudou, R.; et al. Isolation and Biosynthetic Analysis of Haliamide, a New PKS-NRPS Hybrid Metabolite from the Marine Myxobacterium Haliangium ochraceum. Molecules 2016, 21, 59. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Feng, Z.; Tomura, T.; Suzuki, A.; Miyano, S.; Tsuge, T.; Mori, H.; Suh, J.; Iizuka, T.; Fudou, R.; et al. Heterologous Production of the Marine Myxobacterial Antibiotic Haliangicin and Its Unnatural Analogues Generated by Engineering of the Biochemical Pathway. Sci. Rep. 2016, 6, 22091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, M.L.; Paudel, Y.P.; Ross, A.C. Investigating the Biosynthesis of Natural Products from Marine Proteobacteria: A Survey of Molecules and Strategies. Mar. Drugs 2017, 15, 235. [Google Scholar] [CrossRef] [PubMed]
- Amiri Moghaddam, J.; Poehlein, A.; Fisch, K.; Alanjary, M.; Daniel, R.; König, G.M.; Schäberle, T.F. Draft Genome Sequences of the Obligatory Marine Myxobacterial Strains Enhygromyxa salina SWB005 and SWB007. Genome Announc. 2018, 6, e00324-18. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; Gerth, K.; Stadler, M.; Dogma, I.J.; Müller, R. Expanded phylogeny of myxobacteria and evidence for cultivation of the ‘unculturables’. Mol. Phylogenet. Evol. 2010, 57, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Felder, S.; Felder, S.; Dreisigacker, S.; Kehraus, S.; Neu, E.; Bierbaum, G.; Wright, P.R.; Menche, D.; Schäberle, T.F.; König, G.M. Salimabromide: Unexpected chemistry from the obligate marine myxobacterium Enhygromyxa salina. Chemistry 2013, 19, 9319–9324. [Google Scholar] [CrossRef] [PubMed]
- Felder, S.; Kehraus, S.; Neu, E.; Bierbaum, G.; Schäberle, T.F.; König, G.M. Salimyxins and enhygrolides: Antibiotic, sponge-related metabolites from the obligate marine myxobacterium Enhygromyxa salina. ChemBioChem 2013, 14, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Fudou, R.; Iizuka, T.; Nakajima, D.; Okazaki, K.; Shibata, D.; Ojika, M.; Harayama, S. PCR detection of type I polyketide synthase genes in myxobacteria. Appl. Environ. Microbiol. 2008, 74, 5571–5574. [Google Scholar] [CrossRef] [PubMed]
- Bogdanović, X.; Hesseler, M.; Palm, G.J.; Bornscheuer, U.T.; Hinrichs, W. Crystallization and preliminary X-ray diffraction studies of the putative haloalkane dehalogenase DppA from Plesiocystis pacifica SIR-1. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 828–830. [Google Scholar] [CrossRef] [PubMed]
- Hesseler, M.; Bogdanović, X.; Hidalgo, A.; Berenguer, J.; Palm, G.J.; Hinrichs, W.; Bornscheuer, U.T. Cloning, functional expression, biochemical characterization, and structural analysis of a haloalkane dehalogenase from Plesiocystis pacifica SIR-1. Appl. Microbiol. Biotechnol. 2011, 91, 1049–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomura, T.; Nagashima, S.; Yamazaki, S.; Iizuka, T.; Fudou, R.; Ojika, M. An Unusual Diterpene-Enhygromic Acid and Deoxyenhygrolides from a Marine Myxobacterium, Enhygromyxa sp. Mar. Drugs 2017, 15, 109. [Google Scholar] [CrossRef] [PubMed]
- Ojika, M.; Inukai, Y.; Kito, Y.; Hirata, M.; Iizuka, T.; Fudou, R. Miuraenamides: Antimicrobial cyclic depsipeptides isolated from a rare and slightly halophilic myxobacterium. Chem. Asian J. 2008, 3, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Fudou, R.; Jojima, Y.; Ogawa, S.; Yamanaka, S.; Inukai, Y.; Ojika, M. Miuraenamides A and B, Novel Antimicrobial Cyclic Depsipeptides from a New Slightly Halophilic Myxobacterium: Taxonomy, Production, and Biological Properties. J. Antibiot. 2006, 59, 385–391. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Gene | NCBI Reference Sequence | Length (aa) | Highest Homology | Identity (%) |
|---|---|---|---|---|---|
| H. ochraceum | ectC | WP_012827762.1 | 142 | ectoine synthase (Hydrogenophaga crassostreae) | 67% |
| betT | WP_012829907.1 | 538 | BCCT family transporter (Desulfovermiculus halophilus) | 64% | |
| sodium/proline symporter | WP_012825704.1 | 598 | hypothetical protein (Hymenobacter terrenus) | 49% | |
| E. salina | ectC | WP_106088059.1 | 126 | ectoine synthase (Blastopirellula marina) | 63% |
| ectD | AMH38938.1 | 298 | ectoine hydroxylase (Blastopirellula marina) | 59% | |
| betT | AMH38943.1 | 492 | BCCT family transporter (Spingomonas sp. Leaf30) | 57% | |
| sodium/proline symporter | WP_106088061.1 | 481 | sodium/proline symporter (Rubinisphaera brasiliensis) | 59% | |
| sodium/glutamate symporter | KIG18073.1 | 469 | hypothetical protein (P. pacifica) | 64% | |
| P. pacifica | betT (BCCT family transporter) | EDM75025.1 | 512 | BCCT family transporter (Spingomonas sp. Leaf10) | 40% |
| sodium/proline symporter | WP_006976305.1 | 484 | sodium/proline symporter (E. salina) | 56% | |
| sodium/glutamate symporter (hypothetical protein) | WP_006969752.1 | 478 | sodium/glutamate symporter (E. salina) | 64% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albataineh, H.; Stevens, D.C. Marine Myxobacteria: A Few Good Halophiles. Mar. Drugs 2018, 16, 209. https://doi.org/10.3390/md16060209
Albataineh H, Stevens DC. Marine Myxobacteria: A Few Good Halophiles. Marine Drugs. 2018; 16(6):209. https://doi.org/10.3390/md16060209
Chicago/Turabian StyleAlbataineh, Hanan, and D. Cole Stevens. 2018. "Marine Myxobacteria: A Few Good Halophiles" Marine Drugs 16, no. 6: 209. https://doi.org/10.3390/md16060209
APA StyleAlbataineh, H., & Stevens, D. C. (2018). Marine Myxobacteria: A Few Good Halophiles. Marine Drugs, 16(6), 209. https://doi.org/10.3390/md16060209