The Protective Effects of Clams on Hypercholesterolemia in Late-Stage Triple-Transgenic Alzheimer’s Diseased Mice Hearts

 ,
,
Abstract
:1. Introduction
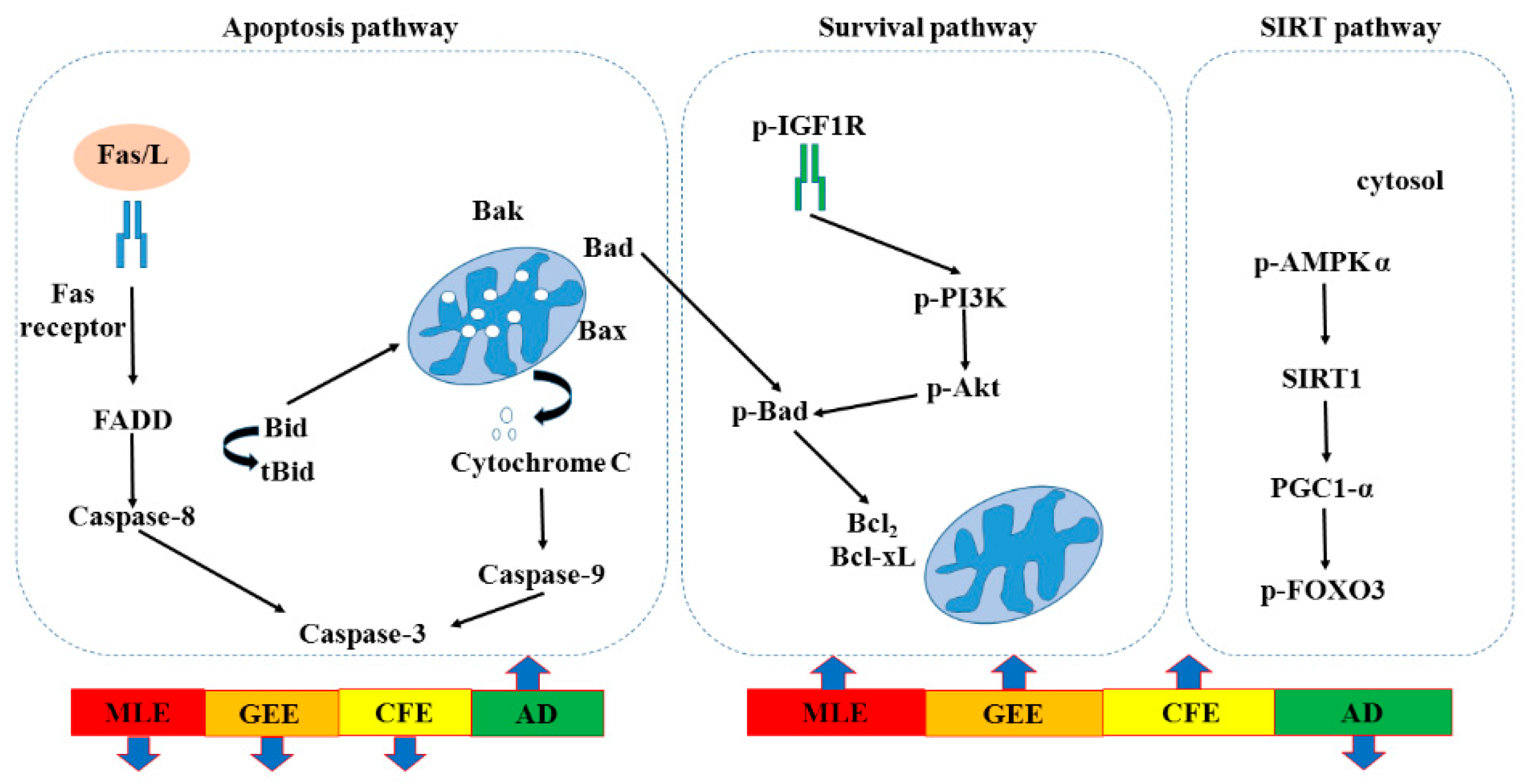
2. Results
3. Discussion
4. Materials and methods
4.1. Materials
4.2. Preparation of Clams
4.3. Animal Test
4.4. Determination of Fatty Acid Profiles
4.5. Tissue Grinding
4.6. Lowry Protein Assay
4.7. Western Blotting
4.8. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AKT | Protein kinase B |
| AMPK | 5’ AMP-activated protein kinase |
| ANOVA | Analysis of variance |
| Bak | Bcl2-antagonist/killer |
| Bcl2 | B-cell lymphoma 2 |
| Bcl-XL | B-cell lymphoma-extra large |
| BSA | bovine serum albumin |
| Caspase | cysteine-aspartic proteases |
| CFE | Corbicula fluminea extract |
| CVD | Cardiovascular disease |
| DHA | Docosahexaenoic acid |
| EPA | Eicosapentaenoic acid |
| FADD | Fas-associated death domain |
| FADD | Fas-associated death domain |
| FAMEs | Fatty acid methyl esters |
| Fas/L | Fas ligand |
| GC | gas chromatography |
| GEE | Geloina eros extract |
| IGF1 | Insulin-like growth factor 1 |
| IGF1R | Insulin-like growth factor 1 receptor |
| MLE | Meretrix lusoria extract |
| MUFA | Monounsaturated fatty acid |
| PI3K | Phosphoinositide 3-kinase |
| PUFA | polyunsaturated fatty acids |
| SFA | Saturated fatty acid |
References
- Yoon, H.; Flores, L.F.; Kim, J. MicroRNAs in brain cholesterol metabolism and their implications for Alzheimer’s disease. BBA Mol. Cell Biol. Lipids 2016, 1861, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Jeyapalan, J.C.; Sedivy, J.M. Cellular senescence and organismal aging. Mech. Ageing Dev. 2008, 129, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D. Evidence of oxidative stress in Alzheimer’s disease brain and antioxidant therapy. Ann. N. Y. Acad. Sci. 2008, 1147, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, L.; Gonzalez de Aguilar, J.L.; Oudart, H.; de Tapia, M.; Barbeito, L.; Loeffler, J.P. Mitochondria in amyotrophic lateral sclerosis: A trigger and a target. Neurodegener. Dis. 2004, 1, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Castillo, E.M.; Goodman-Gruen, D.; Kritz-Silverstein, D.; Morton, D.J.; Wingard, D.L.; Barrett-Connor, E. Sarcopenia in Elderly Men and Women: The Rancho Bernardo study. Am. J. Prev. Med. 2003, 25, 226–231. [Google Scholar] [CrossRef]
- Henchcliffe, C.; Beal, M.F. Mitochondrial biology and oxidative stress in Parkinson disease pathogenesis. Nat. Clin. Pract. Neurol. 2008, 4, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Sindhu, K.K. Oxidative stress and metabolic syndrome. Life Sci. 2009, 84, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Lubos, E.; Handy, D.E.; Loscalzo, J. Role of oxidative stress and nitric oxide in atherothrombosis. Front Biosci. 2008, 13, 5323–5344. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E.; Kamendulis, L.M. The role of oxidative stress in carcinogenesis. Ann. Rev. Pharmacol. Toxicol. 2004, 44, 239–267. [Google Scholar] [CrossRef] [PubMed]
- Mayeux, R.; Stern, Y. Epidemiology of Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2012, 2, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association. Alzheimer’s Association Report 2014 Alzheimer’s disease facts and figures. Alzheimers Dement. 2014, 10, e47–e92. [Google Scholar] [CrossRef]
- Finegolda, J.A.; Asariab, P.; Francisa, D.P. Mortality from ischaemic heart disease by country, region, and age: Statistics from World Health Organisation and United Nations. Int. J. Cardiol. 2013, 168, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Weil, B.R.; Young, R.F.; Shen, X.; Suzuki, G.; Qu, J.; Malhotra, S.; Canty, J.M. Brief myocardial ischemia produces cardiac troponin I release and focal myocyte apoptosis in the absence of pathological infarction in swine. JACC: Basic Transl. Sci. 2017, 2, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Lordan, R.; Demuru, M.; Shiels, K.; Kumar Saha, S.; Nasopoulou, C.; Zabetakis, I. Structural Elucidation of Irish Organic Farmed Salmon (Salmo salar) Polar Lipids with Antithrombotic Activities. Mar. Drugs 2018, 16, 176. [Google Scholar] [CrossRef] [PubMed]
- Tolppanen, A.M.; Kettunen, R.; Ahonen, R.; Soininen, H.; Hartikainen, S. Incident ischaemic heart disease in persons with Alzheimer’s disease in a Finnish nationwide exposure-matched cohort. Int. J. Cardiol. 2013, 170, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yao, L.; Liu, J.; Jiang, Y.; Ma, G. Cardiovascular disease contributes to Alzheimer’s disease: Evidence from large-scale genome-wide association studies. Neurobiol. Aging 2014, 35, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Zulli, R.; Nicosia, F.; Borroni, B.; Agosti, C.; Prometti, P.; Donati, P.; Vecchi, M.D.; Turini, D.; Romanelli, G.; Grassi, V.; et al. Increased prevalence of silent myocardial ischaemia and severe ventricular arrhythmias in untreated patients with Alzheimer’s disease and mild cognitive impairment without overt coronary artery disease. Clin. Neurol. Neurosurg. 2008, 110, 791–796. [Google Scholar] [PubMed]
- Martinić-Popović, I.; Lovrenčić-Huzjan, A.; Demarin, V. Advanced Asymptomatic Carotid Disease and Cognitive Impairment: An Understated Link? Stroke Res. Treat. 2012, 2012, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corder, E.H.; Ervin, J.F.; Lockhart, E.; Szymanski, M.H.; Schmechel, D.E.; Hulette, C.M. Cardiovascular Damage in Alzheimer Disease: Autopsy Findings From the Bryan ADRC. J. Biomed. Biotechnol. 2005, 2, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Chen, L.M.; Chang, M.H.; Chen, W.K.; Tsai, F.J.; Tsai, C.H.; Lai, T.Y.; Kuo, W.W.; Huang, C.Y.; Liu, C.J. Aging and disease: Connections to sirtuins. Aging Cell 2010, 9, 285–290. [Google Scholar]
- Luo, X.Y.; Qu, S.L.; Tang, Z.H.; Zang, Y.; Liu, M.H.; Peng, J.; Tang, H.; Yu, K.L.; Zhang, C.; Ren, Z.; et al. SIRT1 in cardiovascular aging. Clin. Chim. Acta 2014, 437, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Huang, Y.T.; Chang, C.I.; Ho, C.T.; Pan, B.S. Apoptotic-inducing epidioxysterols identified in hard clam (Meretrix lusoria). Food Chem. 2007, 102, 788–795. [Google Scholar] [CrossRef]
- Pan, M.H.; Huang, Y.T.; Ho, C.T.; Chang, C.I.; Hsu, P.C.; Pan, B.S. Induction of apoptosis by Meretrix lusoria through reactive oxygen species production, glutathione depletion, and caspase activation in human leukemia cells. Life Sci. 2006, 79, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.H.; Hsieh, Y.L.; Lee, Y.T.; Hu, C.C. Protective effects of Geloina eros extract against carbon tetrachloride-induced hepatotoxicity in rats. Food Res. Int. 2012, 48, 551–558. [Google Scholar]
- Rovere, M.T.L.; Christensen, J.H. The autonomic nervous system and cardiovascular disease: Role of n-3 PUFAs. Vascul. Pharmacol. 2015, 71, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Loopuijt, L.D.; Sebens, J.B. Loss of dopamine receptors in the olfactory bulb of patients with Alzheimer’s disease. Brain Res. 1990, 529, 239–244. [Google Scholar] [CrossRef]
- Fabian, C.J.; Kimler, B.F.; Hursting, S.D. Omega-3 fatty acids for breast cancer prevention and survivorship. Breast Cancer Res. 2015, 17, 62. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.Y.; Snyder, P.J.; Wu, W.-C.; Zhang, M.; Echeverria, A.; Alber, J. Pathophysiologic relationship between alzheimer’s disease, cerebrovascular disease, and cardiovascular risk: A review and synthesis. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2017, 7, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Schneider, J.A.; Tangney, C.; Tremblay-Mercier, J.; Fortier, M.; Bennett, D.A. Plasma and brain fatty acid profiles in mild cognitive impairment and Alzheimer’s disease. J. Alzheimers Dis. 2012, 29, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Lebbadi, M.; Julien, C.; Phivilay, A.; Tremblay, C.; Emond, V.; Kang, J.X. Endogenous conversion of omega-6 into omega-3 fatty acids improve neuropathology in an animal model of Alzheimer’s disease. J. Alzheimers Dis. 2011, 27, 853–869. [Google Scholar] [CrossRef] [PubMed]
- Uslu, S.; Akarkarasu, Z.E.; Ozbabalik, D.; Ozkan, S.; Colak, O.; Demirkan, E.S. Levels of amyloid beta-42, interleukin-6 and tumor necrosis factor-alpha in Alzheimer’s disease and vascular dementia. Neurochem. Res. 2012, 37, 1554–1559. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.A.; An, J.; Peller, D.; Bowser, R. Total protein is an effective loading control for cerebrospinal fluid western blots. J. Neurosci. Meth. 2015, 251, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Teshima, T.H.N.; Ianez, R.C.F.; Coutinho-Camillo, C.M.; Tucker, A.S.; Lourenço, S.V. Apoptosis-associated protein expression in human salivary gland morphogenesis. Arch. Oral Biol. 2016, 69, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.M.; Ho, T.J.; Yang, A.L.; Chen, I.J.; Kao, C.L.; Wu, F.N.; Lin, J.A.; Kuo, C.H.; Ou, H.C.; Huang, C.Y.; Lee, S.D. Exercise training enhances cardiac IGFI-R/PI3K/Akt and Bcl-2 family associated pro-survival pathways in streptozotocin-induced diabetic rats. Int. J. Cardiol. 2012, 167, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhao, M.; An, W. Increased hepatic apoptosis in high-fat diet-induced NASH in rats may be associated with downregulation of hepatic stimulator substance. J. Mol. Med. 2011, 892, 1207. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, I.L.; Kelley, D.S.; Klasing, K.C.; Erickson, K.L. Regulation of cellular differentiation and apoptosis by fatty acids and their metabolites. Nutr. Res. 2001, 21, 381–393. [Google Scholar] [CrossRef]
- Zheng, S.; Ren, X.; Han, T.; Huiying, Y.C.; Liu, Q.W.; Hu, Y. Fenofibrate attenuates fatty acid-induced islet β-cell dysfunction and apoptosis via inhibiting the NF-κB/MIF dependent inflammatory pathway. Metabolism 2017, 77, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wu, Y.; Zhu, S.; He, Z.; Chen, Y.Q. Omega-3 free fatty acids inhibit tamoxifen-induced cell apoptosis. Biochem. Biophysi. Res. Commun. 2015, 459, 294–299. [Google Scholar]
- Tanaka, Y.; Meguro, K.; Yamaguchi, S.; Ishii, H.; Watanuki, S.; Funaki, Y. Decreased striatal D2 receptor density associated with severe behavioral abnormality in Alzheimer’s disease. Ann. Nucl. Med. 2003, 17, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Amann, M. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef]
- Danaei, G.; Ding, E.L.; Mozaffarian, D.; Taylor, B.; Rehm, J.; Murray, C.J.L.; Ezzati, M. The preventable causes of death in the United States: comparative risk assessment of dietary, lifestyle, and metabolic risk factors. PLoS Med. 2009, 6, e1000058. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.D.; Miller, P.E.; Van Elswyk, M.E.; Kuratko, C.N.; Bylsma, L.C. A meta-analysis of randomized controlled trials and prospective cohort studies of eicosapentaenoic and docosahexaenoic long-chain omega-3 fatty acids and coronary heart disease risk. Mayo Clin. Proc. 2017, 92, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.S.; Lee, S.D.; Kuo, W.W.; Fan, M.J.; Lin, Y.M.; Hu, W.S.; Huang, Y.C.; Velmurugan, B.K.; Tsai, F.J.; Tsai, C.H.; et al. Anti-apoptotic and pro-survival effect of protocatechuic acid on hypertensive hearts. Chem. Biol. Interact. 2014, 209, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.H.; Ho, T.J.; Kuo, W.W.; Day, C.H.; Pai, P.Y.; Chung, L.C.; Liao, P.H.; Lin, F.H.; Wu, E.T.; Huang, C.Y. Exercise training enhanced SIRT1 longevity signaling replaces the IGF1 survival pathway to attenuate aging-induced rat heart apoptosis. AGE 2017, 2014, 9706. [Google Scholar] [CrossRef] [PubMed]
- Zelinkova, Z.; Giri, A.; Wenzl, T. Assessment of critical steps of a GC/MS based indirect analytical method for the determination of fatty acidesters of monochloropropanediols (MCPDEs) and of glycidol (GEs). Food Control 2017, 77, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Pomory, C.M. Color development time of the Lowry protein assay. Anal. Biochem. 2008, 378, 216–217. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.; Datta, K.; Chakraborty, K.; Kulkarni, S.S.; Doiron, K.; Fornace, A.J.J.; Kumar, K.S.; Hauer-Jensen, M.; Ghosh, S.P. Gamma tocotrienol, a potent radioprotector, preferentially upregulates expression of anti-apoptotic genes to promote intestinal cell survival. Food Chem. Toxicol. 2013, 60, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.H.; Lee, Y.T.; Hsieh, H.S.; Hwang, D.F. Dietary caffeic acid, ferulic acid and coumaric acid supplements on cholesterol metabolism and antioxidant activity in rats. J. Food Drug Anal. 2009, 17, 123–132. [Google Scholar]


{kind=link}
| Fatty Acid | MLE (%) | GEE (%) | CFE (%) |
|---|---|---|---|
| Saturated fatty acid (SFA) (% of total fatty acid) | |||
| C12:0 | 0.13 ± 0.02 a | 0.21 ± 0.03 b | 0.15 ± 0.02 a |
| C14:0 | 0.75 ± 0.13 a | 1.23 ± 0.07 b | 0.95 ± 0.08 a |
| C15:0 | 1.56 ± 0.22 c | 0.53 ± 0.10 a | 0.87 ± 0.12 b |
| C16:0 | 12.31 ± 1.56 b | 7.65 ± 1.23 a | 10.56 ± 1.82 a |
| C17:0 | 6.23 ± 0.35 b | 5.76 ± 0.52 b | 3.27 ± 0.32 a |
| C18:0 | 0.21 ± 0.03 a | 0.31 ± 0.02 b | 0.39 ± 0.02 b |
| C20:0 | 1.13 ± 0.06 a | 3.25 ± 0.07 c | 2.87 ± 0.05 b |
| C23:0 | 0.22 ± 0.01 a | 0.32 ± 0.01 c | 1.35 ± 0.03 b |
| Monosaturated fatty acid (MUFA) (% of total fatty acid) | |||
| C14:1 | 0.51 ± 0.02 b | 0.36 ± 0.01 a | 1.56 ± 0.21 c |
| C15:1 | 0.62 ± 0.03 b | 0.21 ± 0.02 a | 0.73 ± 0.01 c |
| C16:1 n-7 | 3.52 ± 0.78 b | 2.36 ± 0.56 a | 4.83 ± 0.32 b |
| C17:1 | 1.23 ± 0.02 c | 0.56 ± 0.03 b | 0.39 ± 0.01 a |
| C18:1 n-11 | 2.32 ± 0.56 b | 1.21 ± 0.21 a | 3.26 ± 0.23 c |
| C18:1 n-9 | 5.13 ± 1.21 a | 6.56 ± 1.65 a | 8.59 ± 1.58 b |
| C18:1 n-7 | 1.16 ± 0.02 b | 2.22 ± 0.03 c | 0.25 ± 0.01 a |
| C20:1 n-7 | 1.33 ± 0.01 a | 2.29 ± 0.01 c | 3.48 ± 0.02 b |
| C20:1 n-9 | 1.25 ± 0.03 b | 0.65 ± 0.02 a | 2.36 ± 0.03 c |
| C20:1 n-11 | 3.65 ± 0.16 b | 2.13 ± 0.11 a | 3.24 ± 0.35 b |
| C22:1 n-9 | 0.26 ± 0.01 a | 2.11 ± 0.02 b | 0.27 ± 0.02 a |
| Polyunsaturated fatty acid (PUFA) (% of total fatty acid) | |||
| C16:3 n-3 | 1.72 ± 0.05 c | 1.21 ± 0.03 a | 1.33 ± 0.05 b |
| C18:2 n-6 | 0.21 ± 0.01 a | 0.36 ± 0.01 b | 0.22 ± 0.02 a |
| C18:3 n-3 | 3.23 ± 0.36 a | 5.63 ± 0.52 b | 3.23 ± 0.65 a |
| C18:3 n-6 | 1.26 ± 0.13 b | 0.82 ± 0.18 a | 1.12 ± 0.15 b |
| C18:4 n-3 | 1.03 ± 0.12 b | 0.75 ± 0.13 a | 1.09 ± 0.15 b |
| C20:1 n-6 | 4.93 ± 0.23 a | 6.31 ± 0.56 b | 4.56 ± 0.62 a |
| C20:3 n-6 | 1.15 ± 0.03 b | 0.53 ± 0.05 a | 2.02 ± 0.07 c |
| C20:3 n-3 | 0.88 ± 0.02 c | 0.62 ± 0.03 b | 0.21 ± 0.02 a |
| C20:4 n-6 | 6.52 ± 0.52 a | 7.53 ± 0.20 b | 5.33 ± 0.67 a |
| C20:4 n-3 | 1.22 ± 0.02 b | 0.76 ± 0.03 a | 1.53 ± 0.06 c |
| C20:5 n-3, EPA | 7.32 ± 0.58 a | 8.50 ± 0.75 b | 6.58 ± 0.23 a |
| C21:5 n-3 | 1.25 ± 0.06 b | 0.12 ± 0.02 a | 1.65 ± 0.01 c |
| C22:4 n-6 | 3.23 ± 0.05 b | 4.52 ± 0.03 c | 2.25 ± 0.03 a |
| C22:5 n-3 | 6.57 ± 0.08 c | 5.26 ± 0.07 b | 3.78 ± 0.07 a |
| C22:5 n-6 | 5.73 ± 0.05 b | 4.95 ± 0.03 a | 6.52 ± 0.06 c |
| C22:6 n-3, DHA | 10.23 ± 0.05 b | 12.21 ± 0.08 c | 9.21 ± 0.03 a |
| Sums of fatty acids | |||
| ƩSFA | 22.54 ± 1.56 b | 19.26 ± 1.52 a | 20.41 ± 1.23 a |
| ƩMUFA | 20.98 ± 2.31 a | 20.66 ± 3.68 a | 28.96 ± 2.23 b |
| ƩPUFA | 56.48 ± 3.62 b | 60.08 ± 2.35 c | 50.63 ± 2.57 a |
| Ʃn-3 | 26.13 ± 1.23 b | 35.06 ± 1.61 b | 28.61 ± 1.20 a |
| Ʃn-6 | 23.03 ± 0.51 b | 25.02 ± 0.31 c | 22.02 ± 0.22 a |
| Fatty Acid | MLE (%) | GEE (%) | CFE (%) |
| Ratios | |||
| PUFA/SFA | 2.50 ± 0.56 a | 3.12 ± 0.52 b | 2.48 ± 0.35 a |
| MUFA/SFA | 0.93 ± 0.01 a | 1.07 ± 0.02 b | 1.42 ± 0.03 c |
| DHA/EPA | 1.40 ± 0.01 a | 1.44 ± 0.02 b | 1.40 ± 0.01 a |
| n-3/n-6 | 1.13 ± 0.02 c | 1.40 ± 0.01 b | 1.30 ± 0.02 a |
| n-6/n-3 | 0.88 ± 0.02 a | 0.71 ± 0.01 b | 0.77 ± 0.01 c |
| Enzyme | Enzyme/β-actin | ||||
|---|---|---|---|---|---|
| Control | AD | AD + MLE | AD + GEE | AD + CFE | |
| Fas receptor | 1.00 ± 0.15 b | 2.65 ± 0.15 a | 1.62 ± 0.18 c | 1.65 ± 0.28 c | 1.86 ± 0.13 c |
| FADD | 1.00 ± 0.13 b | 2.42 ± 0.13 a | 1.53 ± 0.15 c | 1.45 ± 0.12 c | 1.77 ± 0.28 c |
| Caspase-8 | 1.00 ± 0.08 b | 2.75 ± 0.08 a | 1.78 ± 0.27 c | 1.56 ± 0.18 c | 1.89 ± 0.16 c |
| BAK | 1.00 ± 0.07 b | 2.21 ± 0.07 a | 1.35 ± 0.16 c | 1.33 ± 0.19 c | 1.45 ± 0.08 c |
| C-Caspase-9 | 1.00 ± 0.09 b | 2.33 ± 0.09 a | 1.56 ± 0.38 c | 1.68 ± 0.17 c | 1.82 ± 0.28 c |
| Cytochrome-c | 1.00 ± 0.19 b | 2.41 ± 0.89 a | 1.56 ± 0.37 c | 1.42 ± 0.35 c | 1.75 ± 0.33 c |
| Caspase-3 | 1.00 ± 0.32 b | 2.56 ± 0.89 a | 1.46 ± 0.21 c | 1.62 ± 0.28 c | 1.88 ± 0.19 c |
| Enzyme | Enzyme/β-actin | ||||
|---|---|---|---|---|---|
| Control | AD | AD + MLE | AD + GEE | AD + CFE | |
| p-IGF1R | 1.00 ± 0.21 b | 0.76 ± 0.13 c | 3.63 ± 0.36 a | 3.91 ± 0.55 a | 2.32 ± 0.88 a |
| p-PI3K | 1.00 ± 0.15 b | 0.58 ± 0.15 c | 3.37 ± 0.37 a | 2.75 ± 0.65 a | 1.63 ± 0.52 a |
| p-AKT | 1.00 ± 0.16 b | 0.35 ± 0.15 c | 2.55 ± 0.32 a | 2.83 ± 0.53 a | 1.82 ± 0.27 a |
| Bcl-XL | 1.00 ± 0.22 b | 0.25 ± 0.29 c | 1.32 ± 0.27 a | 1.36 ± 0.17 a | 1.25 ± 0.15 a |
| Bcl2 | 1.00 ± 0.11 b | 0.56 ± 0.31 c | 1.35 ± 0.12 a | 1.38 ± 0.13 a | 1.26 ± 0.13 a |
| Enzyme | Enzyme/β-actin | ||||
|---|---|---|---|---|---|
| Control | AD | AD + MLE | AD + GEE | AD + CFE | |
| p-AMPK-α | 1.00 ± 0.13 b | 2.56 ± 0.12 a | 1.23 ± 0.39 b | 1.33 ± 0.27 b | 1.23 ± 0.65 b |
| SIRT1 | 1.00 ± 0.22 b | 2.21 ± 0.23 a | 3.56 ± 0.65 b | 4.46 ± 0.52 b | 3.21 ± 0.53 b |
| PGC1-α | 1.00 ± 0.35 b | 2.65 ± 0.35 a | 1.33 ± 0.29 b | 1.43 ± 0.27 b | 1.31 ± 0.15 b |
| p-FOXO3 | 1.00 ± 0.16 b | 2.36 ± 0.27 a | 1.23 ± 0.19 b | 1.35 ± 0.25 b | 1.13 ± 0.33 b |
| Ingredient | Diets | ||||
|---|---|---|---|---|---|
| Control | AD (%) | AD + MLE (%) | AD + GEE (%) | AD + CFE (%) | |
| Casein | 20 | 20 | 20 | 20 | 20 |
| Methionine | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| Cellulose | 5 | 5 | 5 | 5 | 5 |
| Corn oil | 2 | 2 | 2 | 2 | 2 |
| Cholesterol | 2 | 2 | 2 | 2 | 2 |
| MLE | 0 | 0 | 6 | 0 | 0 |
| GEE | 0 | 0 | 0 | 6 | 0 |
| CFE | 0 | 0 | 0 | 0 | 6 |
| Choline | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Mineral mixa | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin mixb | 1 | 1 | 1 | 1 | 1 |
| Corn starch | 31 | 31 | 25 | 25 | 25 |
| Sucrose | 35 | 35 | 35 | 35 | 35 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, Y.-L.; Teng, H.-J.; Yeh, Y.-H.; Hsieh, C.-H.; Huang, C.-Y. The Protective Effects of Clams on Hypercholesterolemia in Late-Stage Triple-Transgenic Alzheimer’s Diseased Mice Hearts. Mar. Drugs 2018, 16, 263. https://doi.org/10.3390/md16080263
Hsieh Y-L, Teng H-J, Yeh Y-H, Hsieh C-H, Huang C-Y. The Protective Effects of Clams on Hypercholesterolemia in Late-Stage Triple-Transgenic Alzheimer’s Diseased Mice Hearts. Marine Drugs. 2018; 16(8):263. https://doi.org/10.3390/md16080263
Chicago/Turabian StyleHsieh, You-Liang, Hsu-Ju Teng, Yen-Hung Yeh, Cheng-Hong Hsieh, and Chih-Yang Huang. 2018. "The Protective Effects of Clams on Hypercholesterolemia in Late-Stage Triple-Transgenic Alzheimer’s Diseased Mice Hearts" Marine Drugs 16, no. 8: 263. https://doi.org/10.3390/md16080263
APA StyleHsieh, Y. -L., Teng, H. -J., Yeh, Y. -H., Hsieh, C. -H., & Huang, C. -Y. (2018). The Protective Effects of Clams on Hypercholesterolemia in Late-Stage Triple-Transgenic Alzheimer’s Diseased Mice Hearts. Marine Drugs, 16(8), 263. https://doi.org/10.3390/md16080263