Cognitive Impairment in Chronic Obstructive Pulmonary Disease (COPD): Possible Utility of Marine Bioactive Compounds


Abstract
:1. Introduction
2. Mild Cognitive Impairment
2.1. Association of Chronic Obstructive Pulmonary Disease with Mild Cognitive Impairment and Dementia
- i. Abnormal microRNA pattern. MicroRNAs (miRNAs), a class of small non-coding RNAs, are involved in post-transcriptional gene repression. Alterations in miRNA abundance occurs in lung tissue, inflammatory cells, and freely circulating cells in blood, and are thought to function both as drivers and modifiers of disease [61,62]. In COPD patients, -miR-124-3p, miR-34a, miR-124, miR-29c and miR-126 are upregulated; -miR-181c, miR-21, miR-146a, miR-98-5bp are downregulated [61]. In MCI/AD, miRNAs (miR-124-3p, miR-34a, miR-124, miR-181c, miR-21, miR-146a, miR-98-5bp) contribute to the development, differentiation, and synaptic plasticity of neuronal cells, and are involved in many neurodegenerative diseases, including AD [62].
- ii. Activation of PI3K-mTOR signaling. PI3K-AKT-mammalian target of rapamycin (mTOR) pathway is critical for cellular senescence and aging. In parallel, mTOR is a negative regulator of autophagia. There is evidence for PI3K activation in the lungs and cells of COPD patients, as shown by increased expression of the downstream kinase phosphorylated Akt, which in turn activates mTOR [36]. mTOR has a critical role during cognitive function and memory and affects genetic pathways that lead to cognitive loss. An mTOR upstream signaling pathway, the PI3K/Akt axis, is observed in AD brain. Persistent activation of neuronal mTOR signaling is found in MCI and AD brains [63].
- iii. Altered autophagy. Refers to a pathway of cellular self-digestion controlling the degradation of subcellular constituents, including misfolded proteins and damaged organelles. Increasing numbers of autophagic vacuoles are observed in COPD lung tissues under electron microscopy, whereas low vacuole formation is observed in control tissues. Autophagy and mitophagy play a complex role in the lungs, and its related phenomena can have both protective and injurious effects on the progression of COPD. Currently, there is no unifying explanation for the discrepancies between various studies [64]. Although the exact pathological role of autophagy in AD remains to be elucidated, autophagy inducers might provide a new effective therapeutic strategy by degrading aggregates in the early stages of AD. By contrast, the activation of autophagy might enhance disease severity during the late stages of AD, by accelerating Aβ-amyloid production.The autophagy–lysosome pathway is unable to “keep up” with the misfolded protein load that is built up, and becomes defective, causing the aggregation of protein [65]. A mutation in sequestosome1 (SQSTM1), a marker for autophagy that binds cargoes, is identified in patients with familial AD [66].
- iv. Decreased anti-aging molecules. Many endogenous antiaging molecules counteract the mechanisms of senescence, and a reduction in their expression may accelerate the aging process [36]. Sirtuins, are recognized as antiaging molecules that regulate lifespan. Sirtuins are highly conserved NAD+-dependent deacetylases enzymes that play a role in resistance to stress, genomic stability, and energy metabolism. Defective sirtuins are proposed as a mechanism for accelerated lung aging in COPD. SIRT1, 120 kDa (actual size), levels are decreased in patients with COPD, as a result of oxidative stress [67]. SIRT1 decreases with increasing severity of lung emphysema and with a clinical history of frequent COPD exacerbations. Over-expression of a miR-34a causes a significant reduction in both mRNA and protein of SIRT1/-6 in COPD. The aging-suppressor gene, Klotho, is downregulated in COPD [67].There is no direct proof of different levels of SIRT1 in human AD, but overexpression of miR-34a decreases SIRT1 levels. Klotho protein is predominantly secreted by the choroid plexus of the brain, and protects hippocampal neurons from amyloid and glutamate toxicity via the activation of an antioxidant enzymatic system, suggesting Klotho is necessary for oligodendrocyte maturation and myelin integrity.The Klotho KL-VS variant is associated with an increase in the incidence of dementia in older men, in a dose-dependent fashion [68].
- v. Defective DNA damage repair. Effective repair of DNA damage is essential for the survival of cells, and most individual organisms and species. Ineffective repair can result in cell death, cancer, and neurological disease. COPD and AD are associated with excessive DNA damage [69,70]. Several types of DNA damage are associated with neurodegeneration, including bulky adducts, abasic sites, DNA single-strand breaks (SSBs), DNA double-strand breaks (DSBs), base mismatches, insertions, and deletions. DNA repair inefficiency is common in COPD, and is correlated to progression. Poly (ADP-ribose) polymerase (PARP) activation is associated with the progression of COPD [71]. BRCA1 (originally breast cancer 1; currently BRCA1) critically contributes to DSB repair in central neurons and neuronal reductions [69]. BRCA1 causes increased persistence of DSBs, abnormal chromatin remodeling, cellular dysfunction, and cognitive deficits. Depletion of BRCA1 is found in brains of patients with MCI or AD. Depletion of BRCA1 is caused by the pathological accumulation of Aβ, which may promote the proteasomal degradation of BRCA1 through overactivation of extrasynaptic N-methyl-d-aspartate (NMDA) receptor [69].
- vi. Cellular senescence. Senescence is a state of irreversible cell cycle arrest. Senescent cells accumulate in the lung of COPD patients leading to persistent secretory phenotype (SASP) factors, and contributing to increased tissue dysfunction and COPD severity [72]. p16 and p21 are upregulated in cells of COPD patients. Compared with controls, p21 level is significantly decreased in lymphocytes of AD patients, while p53 is increased [72].
- vii. Epigenetic changes. They include DNA methylation, covalent modifications of histone proteins and non-coding RNAs, and increases or decreases in gene transcription. In patients with COPD, there is emerging evidence showing aberrant expression of epigenetic marks, such as DNA methylation, histone modifications, and microRNAs in blood, sputum, and lung tissue [73]. AD entails dramatic losses of histone H4 lysine 16 acetylated isoform (H4K16ac) in the proximity of genes linked to aging and AD. A targeted proteomics approach in human brains showed reduction of histone H3 lysine18 acetylation (H3K18ac) and histone H3 lysine 23 acetylation (H3K23ac) in AD [74].
- viii. Immunosenescence. Immunosenescence affects both innate and adaptive immunity, leading to a loss of function, and is implicated in chronic inflammatory diseases. In COPD, there is reduced immunity, which results in a low-grade chronic inflammatory response. Cluster of differentiation 8 + 28 (CD8 + CD28) null cells from COPD patients have reduced histone deacetylase 2 (HDAC2) expression and are corticosteroid resistant [36]. Patients with mild AD show decreased percentages of naïve cells, elevated memory cells, and increased proportions of CD4+, but not CD8+ cells lacking the important costimulatory receptor CD28 [75].
- ix. Mitochondrial dysfunction. The key function of mitochondria is to produce ATP through the coupling of oxidative phosphorylation with cellular respiration. Mitochondria play a crucial role in the maintenance of intracellular Ca2+ homeostasis, because they can take up substantial amounts of cytosolic Ca2+. COPD is linked to increased mitochondrial reactive oxygen species (ROS) production, decreased intracellular antioxidants, and reduced numbers of mitochondria. The mitochondrial stress markers Parkin and PTEN-induced protein kinase-1 (PINK1) are increased in COPD patients [36]. Mitochondrial deficiency has been suggested to be a hallmark of AD as the patients display early metabolic changes prior to the emergence of any histopathological or clinical abnormalities, showing reduced metabolism, disruption of Ca2+ homeostasis, increased levels of ROS, lipid peroxidation and apoptosis, as well as hyperphosphorylation, aggregation, and damage of axonal transport leading to abnormal mitochondrial distribution. These pathological features impair mitochondrial dynamics by regulating mitochondrial fission/fusion proteins, causing mitochondrial dysfunction and neuronal damage [76].
- x. Stem cell exhaustion. Airway basal progenitor cells are crucial for lung health and resilience because of their ability to repair injured airways. Basal progenitor count, self-renewal, and multipotentiality are all reduced. COPD progenitors produce an epithelium with increased basal and mucous cells and decreased ciliated cells, replicating the COPD phenotype [77].
- xi. Telomere shortening. Studies analyzing leukocyte telomere length (LTL) at the population level have provided ample evidence for the hypothesis that LTL shortening is associated with aging and with age-related chronic diseases (cardiovascular and metabolic disease, cancer), although some inconsistencies have been observed.The exact mechanisms leading to telomere shortening in association with COPD are not yet understood [36]. Increased oxidative stress impairs telomerase activity and, thus, may directly result in telomere shortening. Telomere shortening, in turn, leads to the activation of p21, resulting in cellular senescence and the release of proinflammatory mediators, such as interleukin (IL)-6 and chemokine CXCL8. The telomere length and its rate of shortening did not relate to clinical and lung function parameters [36]. Intermediate values in the aMCI subjects, and the lowest values in the AD patients, suggest a definite relationship between telomere reduction and AD development. Association between APOE genotypes and LTL is observed [78].
2.2. Marine Bioactive Compounds and COPD and MCI/AD
3. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Global Strategy for the Diagnosis, Management and Prevention of Chronic Obstructive Pulmonary Disease. Available online: https://goldcopd.org/gold-2017-global-strategy-diagnosis-management-prevention-copd (accessed on 4 July 2018).
- Vanfleteren, L.E.; Spruit, M.A.; Wouters, E.F.; Franssen, F.M. Management of chronic obstructive pulmonary disease beyond the lungs. Lancet Respir. Med. 2016, 4, 911–924. [Google Scholar] [CrossRef]
- Soriano, J.B.; Abajobir, A.A.; Abate, K.H.; Abera, S.F.; Agrawal, A.; Ahmed, M.B.; Aichour, A.N.; Aichour, I.; Aichour, M.T.E.; Alam, K.; et al. Global, regional, and national deaths, prevalence, disability-adjusted life years, and years lived with disability for chronic obstructive pulmonary disease and asthma, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Respir. Med. 2017, 5, 691–706. [Google Scholar] [CrossRef]
- Agustí, A.; Faner, R. COPD beyond smoking: New paradigm, novel opportunities. Lancet Respir. Med. 2018, 6, 324–326. [Google Scholar] [CrossRef]
- Stoller, J.K.; Lacbawan, F.L.; Aboussouan, L.S. Alpha-1 Antitrypsin Deficiency. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 2006. [Google Scholar]
- Singh, B.; Mielke, M.M.; Parsaik, A.K.; Cha, R.H.; Roberts, R.O.; Scanlon, P.D.; Geda, Y.E.; Christianson, T.J.; Pankratz, V.S.; Petersen, R.C. A prospective study of chronic obstructive pulmonary disease and the risk for mild cognitive impairment. JAMA Neurol. 2014, 71, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Reisberg, B.; Ferris, S.H.; de Leon, M.J.; Franssen, E.S.E.; Kluger, A.; Mir, P.; George, A.E.; Shulman, E.; Steinberg, G.; et al. Stage-specific behavioral, cognitive, and in vivo changes in community residing subjects with age-associated memory impairment and primary degenerative dementia of the Alzheimer type. Drug Dev. Res. 1988, 15, 101–114. [Google Scholar]
- Dubois, B.; Albert, M.L. Amnestic MCI or prodromal Alzheimer’s disease? Lancet Neurol. 2004, 3, 246–248. [Google Scholar] [CrossRef]
- Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013.
- Petersen, R.C.; Negash, S. Mild cognitive impairment: An overview. CNS Spectr. 2008, 13, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Ganguli, M.; Dodge, H.H.; Shen, C.; DeKosky, S.T. Mild cognitive impairment, amnestic type: An epidemiological study. Neurology 2004, 63, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, K. Mild cognitive impairment: An epidemiological perspective. Dialogues Clin. Neurosci. 2004, 6, 401–408. [Google Scholar] [PubMed]
- Apostolo, J.; Holland, C.; O’Connell, M.D.; Feeney, J.; Tabares-Seisdedos, R.; Tadros, G.; Campos, E.; Santos, N.; Robertson, D.A.; Marcucci, M.; et al. Mild cognitive decline. A position statement of the Cognitive Decline Group of the European Innovation Partnership for Active and Healthy Ageing (EIPAHA). Maturitas 2016, 83, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, M.S.; DeKosky, S.T.; Dickson, D.; Dubois, B.; Feldman, H.H.; Fox, N.C.; Gamst, A.; Holtzman, D.M.; Jagust, W.J.; Petersen, R.C.; et al. The diagnosis of mild cognitive impairment due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011, 7, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Hansson, O.; Zetterberg, H.; Buchhave, P.; Londos, E.; Blennow, K.; Minthon, L. Association between CSF biomarkers and incipient Alzheimer’s disease in patients with mild cognitive impairment: A follow-up study. Lancet Neurol. 2006, 5, 228–234. [Google Scholar] [CrossRef]
- Fennema-Notestine, C.; Hagler, D.J.; McEvoy, L.K.; Fleisher, A.S.; Wu, E.H.; Karow, D.S.; Dale, A.M. Structural MRI biomarkers for preclinical and mild Alzheimer’s disease. Hum. Brain Mapp. 2009, 30, 3238–3253. [Google Scholar] [CrossRef] [PubMed]
- Herholz, K. Cerebral glucose metabolism in preclinical and prodromal Alzheimer’s disease. Expert Rev. Neurother. 2010, 10, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Corder, E.H.; Saunders, A.M.; Strittmatter, W.J.; Schmechel, D.E.; Gaskell, P.C.; Small, G.W.; Roses, A.D.; Haines, J.L.; Pericak-Vance, M.A. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset families. Science 1993, 261, 921–923. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, W.J.; Saunders, A.M.; Schmechel, D.; Pericak-Vance, M.; Enghild, J.; Salvesen, G.S.; Roses, A.D. Apolipoprotein E: High-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer disease. Proc. Natl. Acad. Sci. USA 1993, 90, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Bonanni, L.; Thomas, A.; Onofrj, M. Diagnosis and management of dementia with Lewy bodies: Third report of the DLB Consortium. Neurology 2006, 66, 1455. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, S.; Wicklund, A.H.; Salmon, D.P. The neuropsychological profile of Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006171. [Google Scholar] [CrossRef] [PubMed]
- Seelaar, H.; Kamphorst, W.; Rosso, S.M.; Azmani, A.; Masdjedi, R.; de Koning, I.; Maat-Kievit, J.A.; Anar, B.; Donker Kaat, L.; Breedveld, G.J.; et al. Distinct genetic forms of frontotemporal dementia. Neurology 2008, 71, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Kane, R.L.; Butler, M.; Fink, H.A.; Brasure, M.; Davila, H.; Desai, P.; Jutkowitz, E.; McCreedy, E.; Nelson, V.A.; McCarten, J.R.; et al. Interventions to Prevent Age-Related Cognitive Decline, Mild Cognitive Impairment, and Clinical Alzheimer’s-Type Dementia. In Comparative Effectiveness Reviews; Agency for Healthcare Research and Quality: Washington, DC, USA, 2017; Volume 188. [Google Scholar]
- Lee, L.K.; Shahar, S.; Chin, A.V.; Yusoff, N.A. Docosahexaenoic acid-concentrated fish oil supplementation in subjects with mild cognitive impairment (MCI): A 12-month randomised, double-blind, placebo-controlled trial. Psychopharmacology 2013, 225, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Gavrilova, S.I.; Preuss, U.W.; Wong, J.W.; Hoerr, R.; Kaschel, R.; Bachinskaya, N.; GIMCI Plus Study Group. Efficacy and safety of Ginkgo biloba extract EGb 761 in mild cognitive impairment with neuropsychiatric symptoms: A randomized, placebo-controlled, double-blind, multi-center trial. Int. J. Geriatr. Psychiatry 2014, 29, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- DeKosky, S.T.; Williamson, J.D.; Fitzpatrick, A.L.; Kronmal, R.A.; Ives, D.G.; Saxton, J.A.; Lopez, O.L.; Burke, G.; Carlson, M.C.; Fried, L.P.; et al. Ginkgo biloba for prevention of dementia: A randomized controlled trial. JAMA 2008, 300, 2253–2262. [Google Scholar] [CrossRef] [PubMed]
- Cherrier, M.M.; Anderson, K.; Shofer, J.; Millard, S.; Matsumoto, A.M. Testosterone treatment of men with mild cognitive impairment and low testosterone levels. Am. J. Alzheimers Dis. Other Demen. 2015, 30, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Kato-Kataoka, A.; Sakai, M.; Ebina, R.; Nonaka, C.; Asano, T.; Miyamori, T. Soybean-derived phosphatidylserine improves memory function of the elderly Japanese subjects with memory complaints. J. Clin. Biochem. Nutr. 2010, 47, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Naeini, A.M.; Elmadfa, I.; Djazayery, A.; Barekatain, M.; Ghazvini, M.R.; Djalali, M.; Feizi, A. The effect of antioxidant vitamins E and C on cognitive performance of the elderly with mild cognitive impairment in Isfahan, Iran: A double-blind, randomized, placebo-controlled trial. Eur. J. Nutr. 2014, 53, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Smith, S.M.; de Jager, C.A.; Whitbread, P.; Johnston, C.; Agacinski, G.; Oulha, A.; Bradley, K.M.; Jacoby, R.; Refsum, H. Homocysteine-lowering by B vitamins slows the rate of accelerated brain atrophy in mild cognitive impairment: A randomized controlled trial. PLoS ONE 2010, 5, e12244. [Google Scholar] [CrossRef] [PubMed]
- De Jager, C.A.; Oulhaj, A.; Jacoby, R.; Refsum, H.; Smith, A.D. Cognitive and clinical outcomes of homocysteine-lowering B-vitamin treatment in mild cognitive impairment: A randomized controlled trial. Int. J. Geriatr. Psychiatry 2012, 27, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Douaud, G.; Refsum, H.; de Jager, C.A.; Jacoby, R.; Nichols, T.E.; Smith, S.M.; Smith, A.D. Preventing Alzheimer’s disease-related gray matter atrophy by B-vitamin treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 9523–9528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, R.C.; Thomas, R.G.; Grundman, M.; Bennett, D.; Doody, R.; Ferris, S.; Galasko, D.; Jin, S.; Kaye, J.; Levey, A.; et al. Vitamin E and donepezil for the treatment of mild cognitive impairment. N. Engl. J. Med. 2005, 352, 2379–2388. [Google Scholar] [CrossRef] [PubMed]
- Starr, J.M.; Whalley, L.J.; Deary, I.J. The effects of antihypertensive treatment on cognitive function: Results from the HOPE study. J. Am. Geriatr. Soc. 1996, 44, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Starr, J.M.; Whalley, L.J. Differential cognitive outcomes in the Hypertensive Old People in Edinburgh study. J. Neurol. Sci. 2005, 229–230, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Cellular and molecular mechanisms of asthma and COPD. Clin. Sci. 2017, 131, 1541–1558. [Google Scholar] [CrossRef] [PubMed]
- Barnard, N.D.; Bunner, A.E.; Agarwal, U. Saturated and trans fats and dementia: A systematic review. Neurobiol. Aging 2014, 35, S65–S73. [Google Scholar] [CrossRef] [PubMed]
- McGeer, P.L.; Schulzer, M.; McGeer, E.G. Arthritis and anti-inflammatory agents as possible protective factors for Alzheimer’s disease: A review of 17 epidemiologic studies. Neurology 1996, 47, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Small, G.W.; Siddarth, P.; Silverman, D.H.; Ercoli, L.M.; Miller, K.J.; Lavretsky, H.; Bookheimer, S.Y.; Huang, S.C.; Barrio, J.R.; Phelps, M.E. Cognitive and cerebral metabolic effects of celecoxib versus placebo in people with age-related memory loss: Randomized controlled study. Am. J. Geriatr. Psychiatry 2008, 16, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Petersen, R.C.; Grundman, M.; Jin, S.; Gamst, A.; Ward, C.P.; Sencakova, D.; Doody, R.S.; Thal, L.J. Longitudinal MRI findings from the vitamin E and donepezil treatment study for MCI. Neurobiol. Aging 2008, 29, 1285–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildreth, K.L.; Van Pelt, R.E.; Moreau, K.L.; Grigsby, J.; Hoth, K.F.; Pelak, V.; Anderson, C.A.; Parnes, B.; Kittelson, J.; Wolfe, P.; et al. Effects of pioglitazone or exercise in older adults with mild cognitive impairment and insulin resistance: A pilot study. Dement. Geriatr. Cogn. Dis. Extra 2015, 5, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Luchsinger, J.A.; Perez, T.; Chang, H.; Mehta, P.; Steffener, J.; Pradabhan, G.; Ichise, M.; Manly, J.; Devanand, D.P.; Bagiella, E. Metformin in Amnestic Mild Cognitive Impairment: Results of a Pilot Randomized Placebo Controlled Clinical Trial. J. Alzheimers Dis. 2016, 51, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.E.; Yaffe, K. The projected effect of risk factor reduction on Alzheimer’s disease prevalence. Lancet Neurol. 2011, 10, 819–828. [Google Scholar] [CrossRef]
- Newhouse, P.; Kellar, K.; Aisen, P.; White, H.; Wesnes, K.; Coderre, E.; Pfaff, A.; Wilkins, H.; Howard, D.; Levin, E.D. Nicotine treatment of mild cognitive impairment: A 6-month double-blind pilot clinical trial. Neurology 2012, 78, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forlenza, O.V.; Diniz, B.S.; Radanovic, M.; Santos, F.S.; Talib, L.L.; Gattaz, W.F. Disease-modifying properties of long-term lithium treatment for amnestic mild cognitive impairment: Randomised controlled trial. Br. J. Psychiatry 2011, 198, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Knopman, D.S.; Petersen, R.C. Mild cognitive impairment and mild dementia: A clinical perspective. Mayo Clin. Proc. 2014, 89, 1452–1459. [Google Scholar] [CrossRef] [PubMed]
- Langa, K.M.; Levine, D.A. The diagnosis and management of mild cognitive impairment: A clinical review. JAMA 2014, 312, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Fink, H.A.; Jutkowitz, E.; McCarten, J.R.; Hemmy, L.S.; Butler, M.; Davila, H.; Ratner, E.; Calvert, C.; Barclay, T.R.; Brasure, M.; et al. Pharmacologic Interventions to Prevent Cognitive Decline, Mild Cognitive Impairment, and Clinical Alzheimer-Type Dementia: A Systematic Review. Ann. Intern. Med. 2018, 168, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C.; Lopez, O.; Armstrong, M.J.; Getchius, T.S.D.; Ganguli, M.; Gloss, D.; Gronseth, G.S.; Marson, D.; Pringsheim, T.; Day, G.S.; et al. Practice guideline update summary: Mild cognitive impairment: Report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology. Neurology 2018, 90, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, S.; Pepin, V.; Rahayel, S.; Bertrand, J.A.; de Lorimier, M.; Rizk, A.; Desjardins, C.; Parenteau, S.; Beaucage, F.; Joncas, S.; et al. Mild cognitive impairment in moderate to severe COPD: A preliminary study. Chest 2012, 142, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Parsaik, A.K.; Mielke, M.M.; Roberts, R.O.; Scanlon, P.D.; Geda, Y.E.; Pankratz, V.S.; Christianson, T.; Yawn, B.P.; Petersen, R.C. Chronic obstructive pulmonary disease and association with mild cognitive impairment: The Mayo Clinic Study of Aging. Mayo Clin. Proc. 2013, 88, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Yohannes, A.M.; Chen, W.; Moga, A.M.; Leroi, I.; Connolly, M.J. Cognitive Impairment in Chronic Obstructive Pulmonary Disease and Chronic Heart Failure: A Systematic Review and Meta-analysis of Observational Studies. J. Am. Med. Dir. Assoc. 2017, 18, 451.e1–451.e11. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, D.R.; Lavoie, K.L. Recognition, diagnosis, and treatment of cognitive and psychiatric disorders in patients with COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2017, 12, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, Y.; Fei, G.H. The evaluation of cognitive impairment and relevant factors in patients with chronic obstructive pulmonary disease. Respiration 2013, 85, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Schrijvers, E.M.; Koudstaal, P.J.; Hofman, A.; Breteler, M.M. Plasma clusterin and the risk of Alzheimer disease. JAMA 2011, 305, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Nuutinen, T.; Suuronen, T.; Kauppinen, A.; Salminen, A. Clusterin: A forgotten player in Alzheimer’s disease. Brain Res. Rev. 2009, 61, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Jongbloed, W.; van Dijk, K.D.; Mulder, S.D.; van de Berg, W.D.; Blankenstein, M.A.; van der Flier, W.; Veerhuis, R. Clusterin Levels in Plasma Predict Cognitive Decline and Progression to Alzheimer’s Disease. J. Alzheimers Dis. 2015, 46, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.R.; Zoubeidi, A. Clusterin as a therapeutic target. Expert Opin. Ther. Targets. 2017, 21, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, S.; Luppi, F.; D’Arca, D.; Caporali, A.; Ruggieri, M.P.; Vettori, M.V.; Caglieri, A.; Astancolle, S.; Panico, F.; Davalli, P.; et al. Clusterin decreases oxidative stress in lung fibroblasts exposed to cigarette smoke. Am. J. Respir. Crit. Care Med. 2006, 174, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. The Continuum of Aging and Age-Related Diseases: Common Mechanisms but Different Rates. Front. Med. 2018, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Salimian, J.; Mirzaei, H.; Moridikia, A.; Harchegani, A.B.; Sahebkar, A.; Salehi, H. Chronic obstructive pulmonary disease: MicroRNAs and exosomes as new diagnostic and therapeutic biomarkers. J. Res. Med. Sci. 2018, 23, 27. [Google Scholar] [CrossRef] [PubMed]
- Zendjabil, M. Circulating microRNAs as novel biomarkers of Alzheimer’s disease. Clin. Chim. Acta 2018, 484, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, F.; Barone, E.; Perluigi, M.; Butterfield, D.A. The Triangle of Death in Alzheimer’s Disease Brain: The Aberrant Cross-Talk Among Energy Metabolism, Mammalian Target of Rapamycin Signaling, and Protein Homeostasis Revealed by Redox Proteomics. Antioxid. Redox Signal. 2017, 26, 364–387. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S. Autophagy in the respiratory diseases. Respir. Investig. 2016, 54, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Fujikake, N.; Shin, M.; Shimizu, S. Association Between Autophagy and Neurodegenerative Diseases. Front. Neurosci. 2018, 12, 255. [Google Scholar] [CrossRef] [PubMed]
- White, J.A.; Banerjee, R.; Gunawardena, S. Axonal Transport and Neurodegeneration: How Marine Drugs Can Be Used for the Development of Therapeutics. Mar. Drugs 2016, 14, 102. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, S.; Papaioannou, A.I.; Papaporfyriou, A.; Baker, J.R.; Vuppusetty, C.; Loukides, S.; Barnes, P.J.; Ito, K. Decreased Serum Sirtuin-1 in COPD. Chest 2017, 152, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Almeida, O.P.; Morar, B.; Hankey, G.J.; Yeap, B.B.; Golledge, J.; Jablensky, A.; Flicker, L. Longevity Klotho gene polymorphism and the risk of dementia in older men. Maturitas 2017, 101, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Suberbielle, E.; Djukic, B.; Evans, M.; Kim, D.H.; Taneja, P.; Wang, X.; Finucane, M.; Knox, J.; Ho, K.; Devidze, N.; et al. DNA repair factor BRCA1 depletion occurs in Alzheimer brains and impairs cognitive function in mice. Nat. Commun. 2015, 6, 8897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milic, M.; Frustaci, A.; Del Bufalo, A.; Sánchez-Alarcón, J.; Valencia-Quintana, R.; Russo, P.; Bonassi, S. DNA damage in non-communicable diseases: A clinical and epidemiological perspective. Mutat. Res. 2015, 776, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.S.; Dharwal, V.; Naura, A.S. Poly(ADP-Ribose)Polymerase-1 in Lung Inflammatory Disorders: A Review. Front. Immunol. 2017, 8, 1172. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Chand, H.S.; Bruse, S.; Doyle-Eisele, M.; Royer, C.; McDonald, J.; Qualls, C.; Klingelhutz, A.J.; Lin, Y.; Mallampalli, R.; et al. Connective Tissue Growth Factor Promotes Pulmonary Epithelial Cell Senescence and Is Associated with COPD Severity. COPD 2017, 14, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, T.; Aoshiba, K.; Nagai, A. Alveolar cell senescence in patients with pulmonary emphysema. Am. J. Respir. Crit. Care Med. 2006, 174, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Nativio, R.; Donahue, G.; Berson, A.; Lan, Y.; Amlie-Wolf, A.; Tuzer, F.; Toledo, J.B.; Gosai, S.J.; Gregory, B.D.; Torres, C.; et al. Dysregulation of the epigenetic landscape of normal aging in Alzheimer’s disease. Nat. Neurosci. 2018, 21, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Larbi, A.; Pawelec, G.; Witkowski, J.M.; Schipper, H.M.; Derhovanessian, E.; Goldeck, D.; Fulop, T. Dramatic shifts in circulating CD4 but not CD8 T cell subsets in mild Alzheimer’s disease. J. Alzheimers Dis. 2009, 17, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Bai, F. The Association of Tau With Mitochondrial Dysfunction in Alzheimer’s Disease. Front. Neurosci. 2018, 12, 163. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Miller, Y.E.; Nakachi, I.; Kwon, J.B.; Barón, A.E.; Brantley, A.E.; Merrick, D.T.; Franklin, W.A.; Keith, R.L.; Vandivier, R.W. Exhaustion of Airway Basal Progenitor Cells in Early and Established Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit Care Med. 2018, 197, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Scarabino, D.; Broggio, E.; Gambina, G.; Corbo, R.M. Leukocyte telomere length in mild cognitive impairment and Alzheimer’s disease patients. Exp. Gerontol. 2017, 98, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Lamonaca, P.; Prinzi, G.; Kisialiou, A.; Cardaci, V.; Fini, M.; Russo, P. Metabolic Disorder in Chronic Obstructive Pulmonary Disease (COPD) Patients: Towards a Personalized Approach Using Marine Drug Derivatives. Mar. Drugs 2017, 15, 81. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Gao, W.Y.; Yan, Y.Q.; Wang, J.; Xu, Z.H.; Zheng, W.J.; Xiao, P.G. The rediscovery of ancient Chinese herbal formulas. Phytother. Res. 2004, 18, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Nastrucci, C.; Cesario, A.; Russo, P. Anticancer drug discovery from the marine environment. Recent Pat. Anticancer Drug Discov. 2012, 7, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Cesario, A. New anticancer drugs from marine cyanobacteria. Curr. Drug Targets 2012, 13, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Del Bufalo, A.; Fini, M. Deep sea as a source of novel-anticancer drugs: Update on discovery and preclinical/clinical evaluation in a systems medicine perspective. EXCLI J. 2015, 14, 228–236. [Google Scholar] [PubMed]
- Russo, P.; Kisialiou, A.; Lamonaca, P.; Moroni, R.; Prinzi, G.; Fini, M. New Drugs from Marine Organisms in Alzheimer’s Disease. Mar. Drugs 2016, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.Y.; Park, W.S.; Lee, D.S.; Choi, G.; Yim, M.J.; Lee, J.M.; Jung, W.K.; Park, S.G.; Seo, S.K.; Park, S.J.; et al. Fucoxanthin inhibits profibrotic protein expression in vitro and attenuates bleomycin-induced lung fibrosis in vivo. Eur. J. Pharmacol. 2017, 811, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.; Wang, H.D.; Gao, R.; Chang, Y.C.; Hu, F.; Meng, X.; Huang, S.Y. Marine-derived protein kinase inhibitors for neuroinflammatory diseases. Biomed. Eng. Online 2018, 17, 46. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.J.; Chen, W.F.; Guo, B.L.; Feng, C.W.; Hung, H.C.; Yang, W.Y.; Sung, C.S.; Tsui, K.H.; Chu, H.; Chen, N.F.; et al. The 1-Tosylpentan-3-one Protects against 6-Hydroxydopamine-Induced Neurotoxicity. Int. J. Mol. Sci. 2017, 18, 1096. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.C.; Cho, K.H.; Ko, W.; Yoon, C.S.; Sohn, J.H.; Yim, J.H.; Kim, Y.C.; Oh, H. Anti-Inflammatory and Cytoprotective Effects of TMC-256C1 from Marine-Derived Fungus Aspergillus sp. SF-6354 via up-Regulation of Heme Oxygenase-1 in Murine Hippocampal and Microglial Cell Lines. Int. J. Mol. Sci. 2016, 17, 529. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Luo, Y.; Tang, R.; Zhang, H.; Ye, Y.; Xiang, L.; Qi, J. Neuritogenic monoglyceride derived from the constituent of a marine fish for activating the PI3K/ERK/CREB signalling pathways in PC12 cells. Int. J. Mol. Sci. 2013, 14, 24200–24210. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.K.; Kamei, Y. Sargaquinoic acid supports the survival of neuronal PC12D cells in a nerve growth factor-independent manner. Eur. J. Pharmacol. 2004, 488, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Lee, J.H.; Chand, H.S.; Lee, J.S.; Lin, Y.; Weathington, N.; Mallampalli, R.; Jeon, Y.J.; Nyunoya, T. APO-9′-Fucoxanthinone Extracted from Undariopsis peteseniana Protects Oxidative Stress-Mediated Apoptosis in Cigarette Smoke-Exposed Human Airway Epithelial Cells. Mar. Drugs 2016, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wu, F.; Wen, M.; Yanagita, T.; Xue, C.; Zhang, T.; Wang, Y. The Protective Effect of Antarctic Krill Oil on Cognitive Function by Inhibiting Oxidative Stress in the Brain of Senescence-Accelerated Prone Mouse Strain 8 (SAMP8) Mice. J. Food Sci. 2018, 83, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Lin, Y.C.; Chen, B.R.; Wu, S.C.; Lee, B.H. The neuronal protection of a zinc-binding protein isolated from oyster. Food Chem. Toxicol. 2018, 114, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Leirós, M.; Alonso, E.; Rateb, M.E.; Houssen, W.E.; Ebel, R.; Jaspars, M.; Alfonso, A.; Botana, L.M. Gracilins: Spongionella-derived promising compounds for Alzheimer disease. Neuropharmacology 2015, 93, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Leirós, M.; Sánchez, J.A.; Alonso, E.; Rateb, M.E.; Houssen, W.E.; Ebel, R.; Jaspars, M.; Alfonso, A.; Botana, L.M. Spongionella secondary metabolites protect mitochondrial function in cortical neurons against oxidative stress. Mar. Drugs 2014, 12, 700–718. [Google Scholar] [CrossRef] [PubMed]
- Mokdad, A.H.; Ballestros, K.; Echko, M.; Glenn, S.; Olsen, H.E.; Mullany, E.; Lee, A.; Khan, A.R.; Ahmadi, A.; Ferrari, A.J.; et al. The State of US Health, 1990–2016: Burden of Diseases, Injuries, and Risk Factors Among US States. JAMA 2018, 319, 1444–1472. [Google Scholar] [PubMed]
- Annuario Statistico Italiano 2017; Istat: Roma, Italy, 2017.
- Calverley, P.M.; Anderson, J.A.; Celli, B.; Ferguson, G.T.; Jenkins, C.; Jones, P.W.; Yates, J.C.; Vestbo, J.; TORCH investigators. Salmeterol and fluticasone propionate and survival in chronic obstructive pulmonary disease. N. Engl. J. Med. 2007, 356, 775–789. [Google Scholar] [PubMed]
- Germini, F.; Veronese, G.; Marcucci, M.; Coen, D.; Ardemagni, D.; Montano, N.; Fabbri, A.; SIMEU Study Group. COPD exacerbations in the emergency department: Epidemiology and related costs. A retrospective cohort multicentre study from the Italian Society of Emergency Medicine (SIMEU). Eur. J. Intern. Med. 2018, 51, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Grant, I.; Heaton, R.K.; McSweeny, A.J.; Adams, K.M.; Timms, R.M. Neuropsychologic findings in hypoxemic chronic obstructive pulmonary disease. Arch. Intern. Med. 1982, 142, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.S.; Chen, S.; McAvay, G.J.; Tinetti, M.E. Effect of coexisting chronic obstructive pulmonary disease and cognitive impairment on health outcomes in older adults. J. Am. Geriatr. Soc. 2012, 60, 1839–1846. [Google Scholar] [CrossRef] [PubMed]
- Dodd, J.W.; Charlton, R.A.; van den Broek, M.D.; Jones, P.W. Cognitive dysfunction in patients hospitalized with acute exacerbation of COPD. Chest 2013, 144, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Baird, C.; Lovell, J.; Johnson, M.; Shiell, K.; Ibrahim, J.E. The impact of cognitive impairment on self-management in chronic obstructive pulmonary disease: A systematic review. Respir. Med. 2017, 129, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.C.; Baxter, M. A comparison of four tests of cognition as predictors of inability to perform spirometry in old age. Age Ageing 2009, 38, 537–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkeshi, E.; Zelenukha, D.; Vaes, B.; Andreeva, E.; Frolova, E.; Degryse, J.M. Predictors of poor-quality spirometry in two cohorts of older adults in Russia and Belgium: A cross-sectional study. NPJ Prim. Care Respir. Med. 2015, 25, 15048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knorr-Held, L.; Best, N.G. A shared component model for detecting joint and selective clustering of two diseases. J. Royal Statist. Soc. 2001, 164, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Loscalzo, J.; Kohane, I.; Barabasi, A.L. Human disease classification in the postgenomic era: A complex systems approach to human pathobiology. Mol. Syst. Biol. 2007, 3, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lococo, F.; Cesario, A.; Del Bufalo, A.; Ciarrocchi, A.; Prinzi, G.; Mina, M.; Bonassi, S.; Russo, P. Novel therapeutic strategy in the management of COPD: A systems medicine approach. Curr. Med. Chem. 2015, 22, 3655–3675. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, R.; Zhao, S.; Chung, S.W.; Mager, D.E.; Gallo, J.M. Merging systems biology with pharmacodynamics. Sci. Transl. Med. 2012, 4, 126ps7. [Google Scholar] [CrossRef] [PubMed]
- Harrold, J.M.; Ramanathan, M.; Mager, D.E. Network-based approaches in drug discovery and early development. Clin. Pharmacol. Ther. 2013, 94, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, a Marine Carotenoid, Reverses Scopolamine-Induced Cognitive Impairments in Mice and Inhibits Acetylcholinesterase in Vitro. Mar. Drugs 2016, 14, 77. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Adki, K.M. Marine natural products for multi-targeted cancer treatment: A future insight. Biomed. Pharmacother. 2018, 105, 233–245. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Markers | Methodology | References |
|---|---|---|
| AD PATHOLOGY | ||
| Biomarkers of amyloid (Aβ) deposition | -Cerebrospinal fluid (CSF) concentrations of Aβ42 (decreased CSF Aβ42 levels) -Positron emission tomography (PET) amyloid imaging | [14,15] |
| Biomarkers of neuronal injury | -CSF concentrations of /phosphorylated(increased CSF/p levels) -Hippocampal volume or medial temporal atrophy or rate of brain atrophy measured by structural MRI -Decreased glucose metabolism in temporoparietal regions on fluorodeoxyglucose PET imaging | [15,16,17] |
| Presence of AD genetic risk factors | Variation in apolipoprotein E gene located on chromosome 19 | [18,19] |
| LEWY BODY PATHOLOGY | ||
| -Visual hallucinations -Parkinsonism -Motor features (bradykinesia, rigidity) -Rapid eye movement during sleep -Abnormalities suggesting pathological processes associated with dementia and with Lewy bodies (DLB) | [20] | |
| VASCULAR DISEASE | ||
| Multiple vascular risk factors suggesting pathological processes associated with vascular dementia | -Presence of extensive cerebrovascular disease evident by structural MRI -“Step-wise” decline | [21] |
| FRONTOTEMPORAL DEGENERATION | ||
| -Frontal lobe atrophy evident by structural MRI -Decreased glucose metabolism in the frontal and temporal lobes evident by PET -Mutations in the microtubule-associatedprotein gene on chromosome 17 | [22] | |
| Intervention | Rationale | Key Message | References |
|---|---|---|---|
| Nutraceuticals | Targeted pathways include: -reducing oxidative stress and chronic inflammation -improving vascular function -supplementing macronutrients found in brain tissue and used in brain function | Few studies examined the effects of nutraceuticals on adults with MCI (i.e., omega-3, fatty acids, ginkgo biloba) | [24,25,26] |
| Hormone therapy | Speculation of the relationship between the pituitary endocrine axis and aging | -Low-strength evidence suggests that estrogen therapy may slightly increase the risk of probable MCI -Low-strength evidence suggests that faloxifene may decrease the risk of MCI compared to placebo -No effect of soybean-derived phosphatidylserine -Hormone therapy has been associated with serious adverse events, including increased risk of certain cancers and cardiovascular disease | [27,28] |
| Vitamin(s) | In the case of B vitamins the targeted pathway may involve lowering of homocysteine levels | -Moderate-strength evidence shows no benefit in cognitive performance for vitamin E in women -B vitamins show mixed findings -Low-strength evidence shows no benefit in cognitive performance for multivitamins, vitamin C (in women), vitamin D with calcium (in women), or -carotene (in women) -Low-strength evidence shows no benefit in incident MCI for multivitamins or vitamin D with calcium -In adults with MCI, low-strength evidence shows no benefit for vitamin E | [29,30,31,32,33] |
| Antihypertensive | Hypertension is thought to contribute to risk of both vascular and AD dementia through unclear vascular mechanisms. Presumably hypertension is the cause or result of vascular changes in the blood supply to the brain that can adversely affect its function. It remains unclear whether this is a direct effect or the result of other factors that affect both the vasculature and the brain. | Generally, low-strength evidence shows that 3 to 4.7 years of antihypertensive treatment regimens versus placebo appears to have no benefit on cognitive test performance in adults MCI | [34,35,36] |
| Lipid lowering treatment | Saturated fat intake is positively associated with MCI, or cognitive decline. | Evidence was insufficient to assess the effect of 5 years statin treatment on preventing MCI | [37] |
| Non-steroidal antiinflammatory drugs (NSAIDs) | Numerous epidemiological studies have shown an association between NSAID use and a reduced prevalence of dementia, specifically AD. In vitro and animal models of AD pathology show that NSAIDs reduce plaque-related inflammation and improve function, both at a cellular and behavioral level | No evidence is available for the effect of low-dose aspirin on MCI | [38,39] |
| Anti-dementia | The acetylcholinesterase inhibitors (AChEIs) have consistently demonstrated a modest but positive benefit to cognition in persons with mild through severe stages. They may likewise provide benefit to persons with age-related cognitive decline or MCI through the same mechanisms of action by increasing the duration of action of acetylcholine in the synapse through inhibition of its breakdown by AChE. The drugs have been approved by the FDA/EMA for people with mild to moderate AD but not for people with age-related cognitive decline or MCI. | -Low-strength evidence shows AChEIs do not reduce the incidence of AD in persons with MCI for over 3 years -Low-strength evidence shows AChEIs for 3 years have no significant effect on cognitive performance in adults with MCI | [33,40] |
| Diabetes medication | Diabetes may increase risk of AD through: -vascular mechanisms -direct effects of elevated blood glucose -insulin-resistance associated inflammation, and/or a pathway in which peripheral hyperinsulinemia inhibits brain insulin production, which then results in impaired brain Aclearance | No studies report on the effect of diabetes treatment on the risk of incident clinical diagnoses of MCI. | [41,42,43] |
| Other drugs | Evidence was insufficient for lithium, or for nicotine patch | [44,45] |
| Drug | Mechanism | References | |
|---|---|---|---|
| COPD | MCI/AD | ||
| Activation PI3K-mTOR | |||
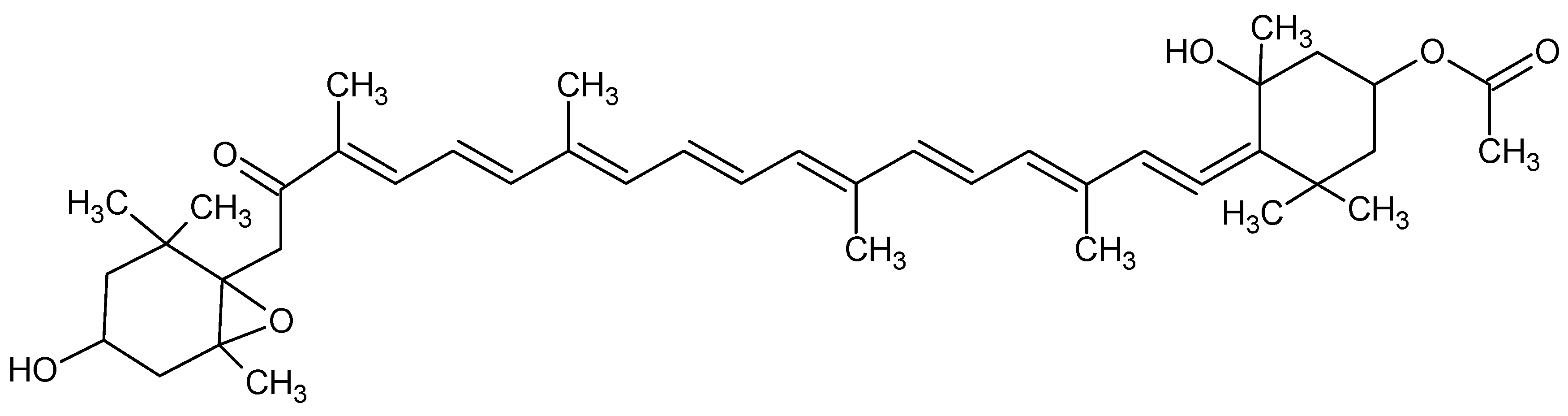
| Fucoxanthin | Inhibition of mice bleomicyn-induced lung fibrosis | Neuroprotective | [85,86] |
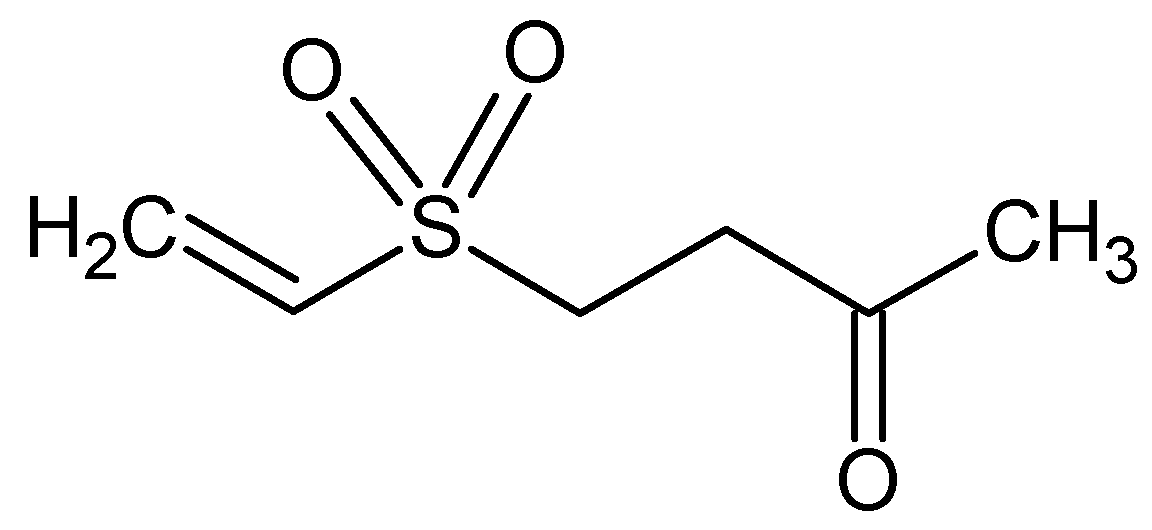
| Austrasulfone | NA | Anti-apoptotic | [87] |
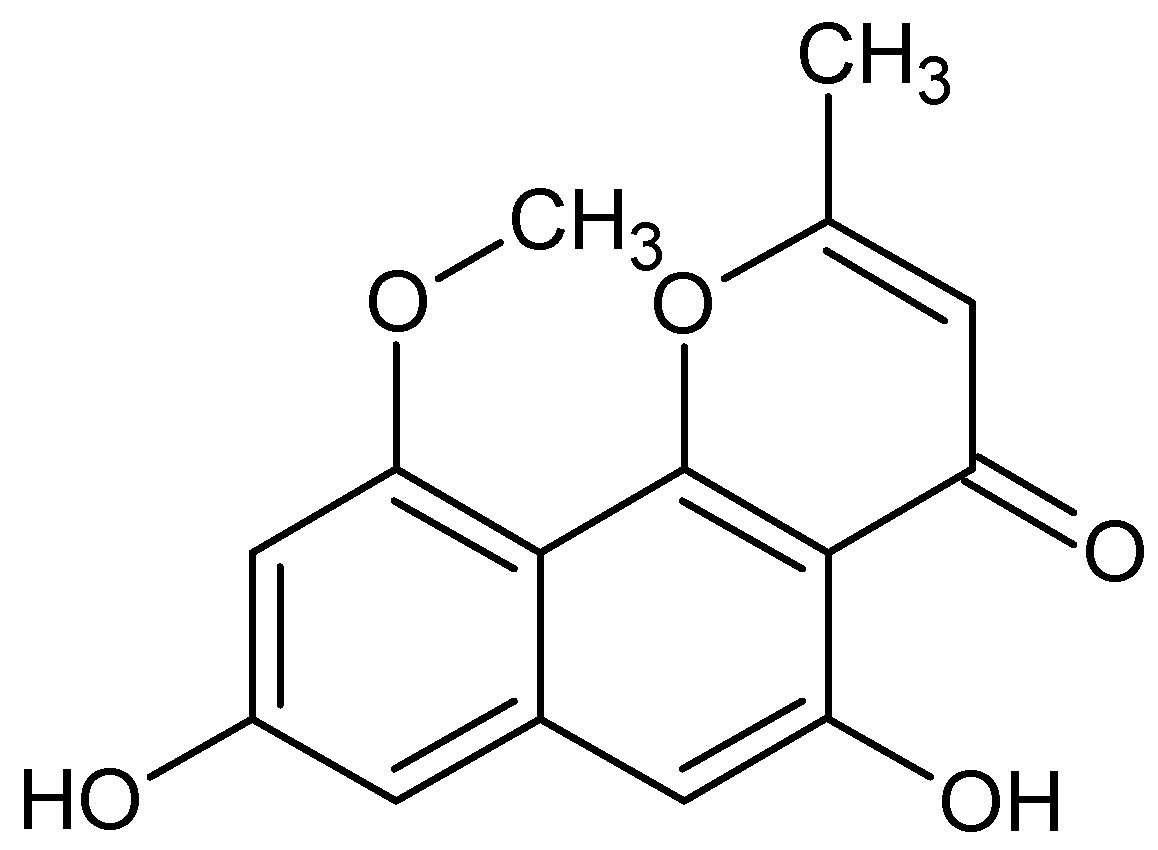
| TMC-256C1 | NA | Activation of kinases | [88] |
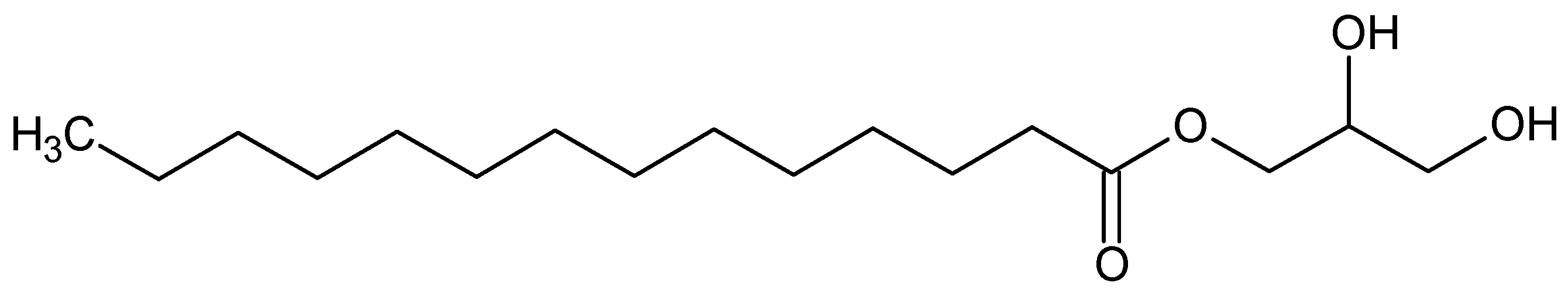
| 1-O-(Myristoyl) glycerol (MG) | NA | Neurite outgrowth | [89] |
| Sargaquinoic acid | NA | Enhancement of neuriteregeneration | [90] |
| Altered autophagy | |||
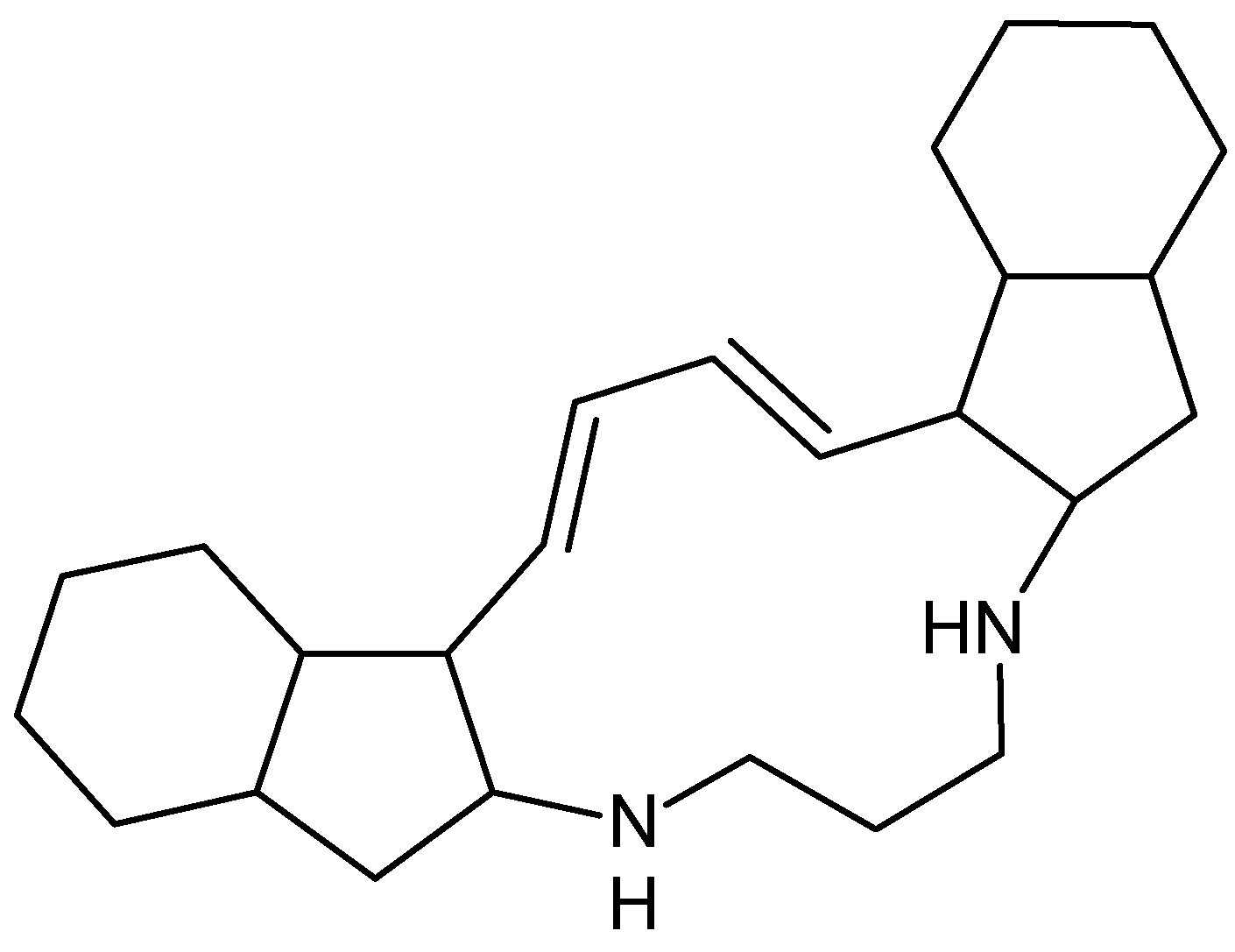
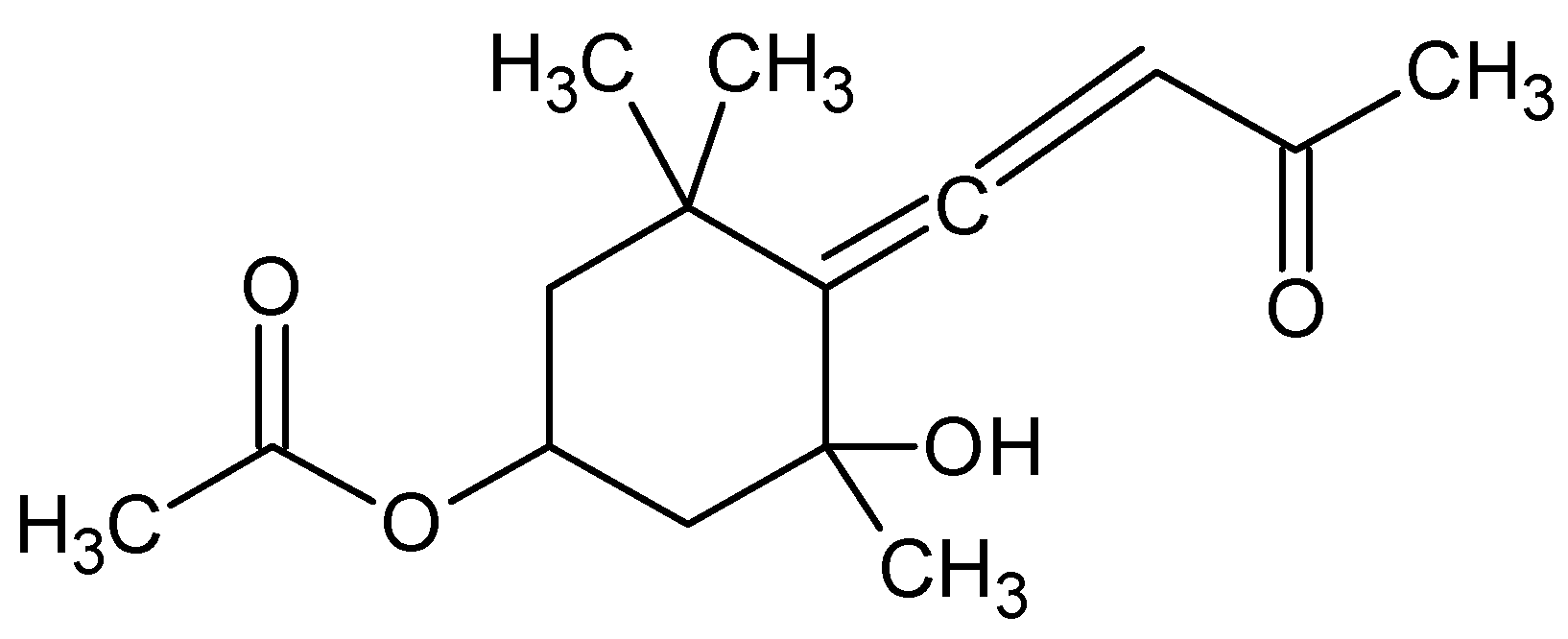
| Bafilomycins, coibamide A, manzamine A and papuamine | NA | Inhibits autophagy Induces autophagy Induces autophagy | [66] |
| Defective DNA damage repair | |||
| Apo-9′-fucoxanthinone | Decreases cigarette smoke extract-induced DNA damage | [91] | |
| Cellular senescence | |||
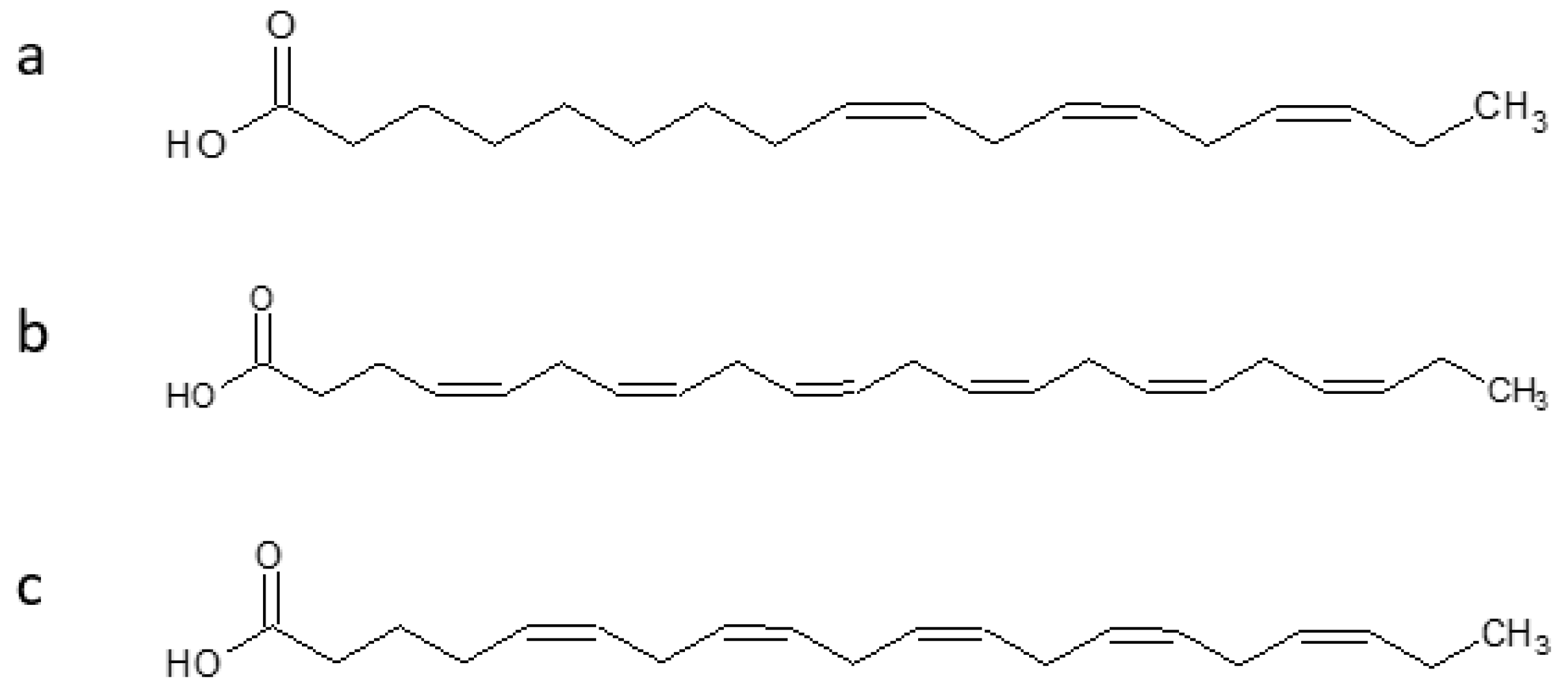
| Antarctic krill oil (AKO) | NA | -Protective effect against AD senescence -Ameliorates learning and memory deficits and eases the anxiety -Reduces Aβ accumulation | [92] |
| Mitochondrial dysfunction | |||
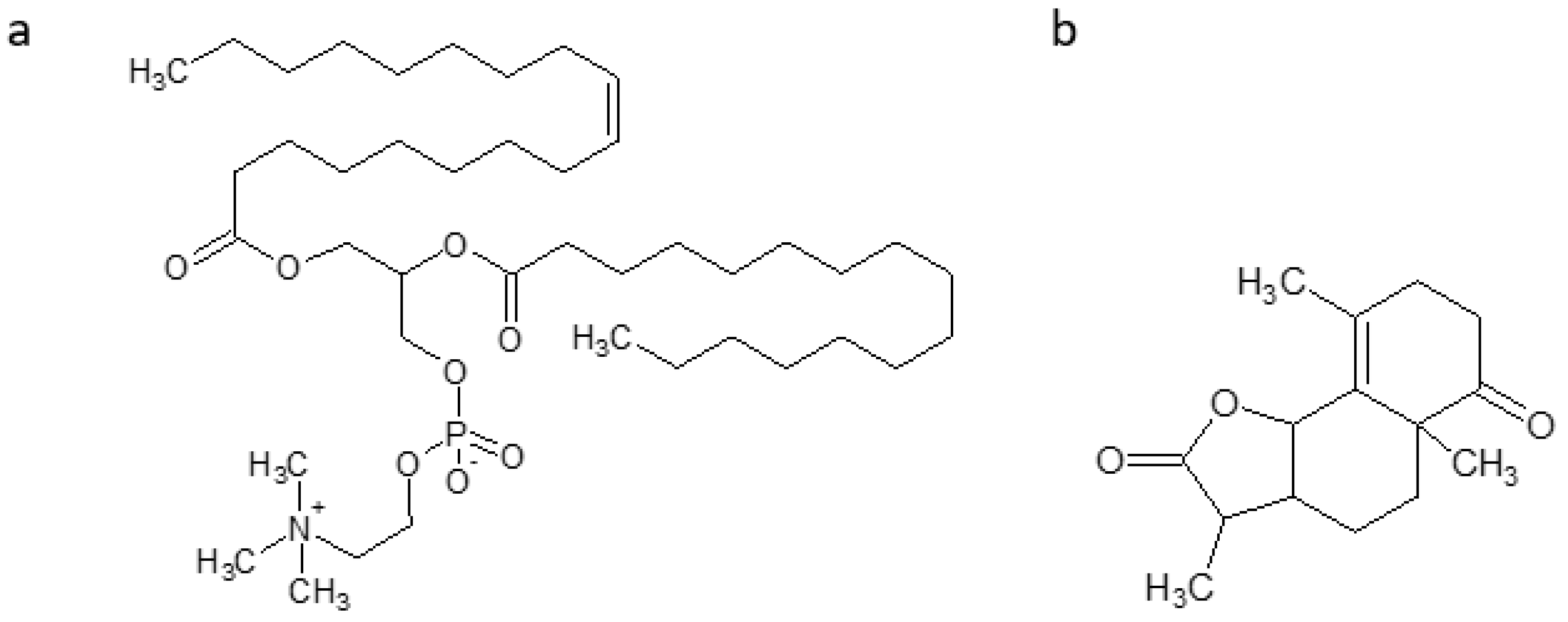
| 43 kD protein | NA | -Attenuates neuronal cell death. -Alleviates mitochondrial inactivation -Recovers metallathionein-3 (MT-3) -Decreases and inhibits β- and γ-secretase, as well as Aβ accumulation | [93] |
| Gracilins | NA | -Positive trend on learning and spatial memory of treated mice. --Decreases Aβ42 and hyperphosphorylated levels -Preserves neurons against oxidative damage | [94,95] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prinzi, G.; Santoro, A.; Lamonaca, P.; Cardaci, V.; Fini, M.; Russo, P. Cognitive Impairment in Chronic Obstructive Pulmonary Disease (COPD): Possible Utility of Marine Bioactive Compounds. Mar. Drugs 2018, 16, 313. https://doi.org/10.3390/md16090313
Prinzi G, Santoro A, Lamonaca P, Cardaci V, Fini M, Russo P. Cognitive Impairment in Chronic Obstructive Pulmonary Disease (COPD): Possible Utility of Marine Bioactive Compounds. Marine Drugs. 2018; 16(9):313. https://doi.org/10.3390/md16090313
Chicago/Turabian StylePrinzi, Giulia, Alessia Santoro, Palma Lamonaca, Vittorio Cardaci, Massimo Fini, and Patrizia Russo. 2018. "Cognitive Impairment in Chronic Obstructive Pulmonary Disease (COPD): Possible Utility of Marine Bioactive Compounds" Marine Drugs 16, no. 9: 313. https://doi.org/10.3390/md16090313
APA StylePrinzi, G., Santoro, A., Lamonaca, P., Cardaci, V., Fini, M., & Russo, P. (2018). Cognitive Impairment in Chronic Obstructive Pulmonary Disease (COPD): Possible Utility of Marine Bioactive Compounds. Marine Drugs, 16(9), 313. https://doi.org/10.3390/md16090313