Photosynthetic Accumulation of Lutein in Auxenochlorella protothecoides after Heterotrophic Growth

Abstract
:1. Introduction
2. Results and Discussion
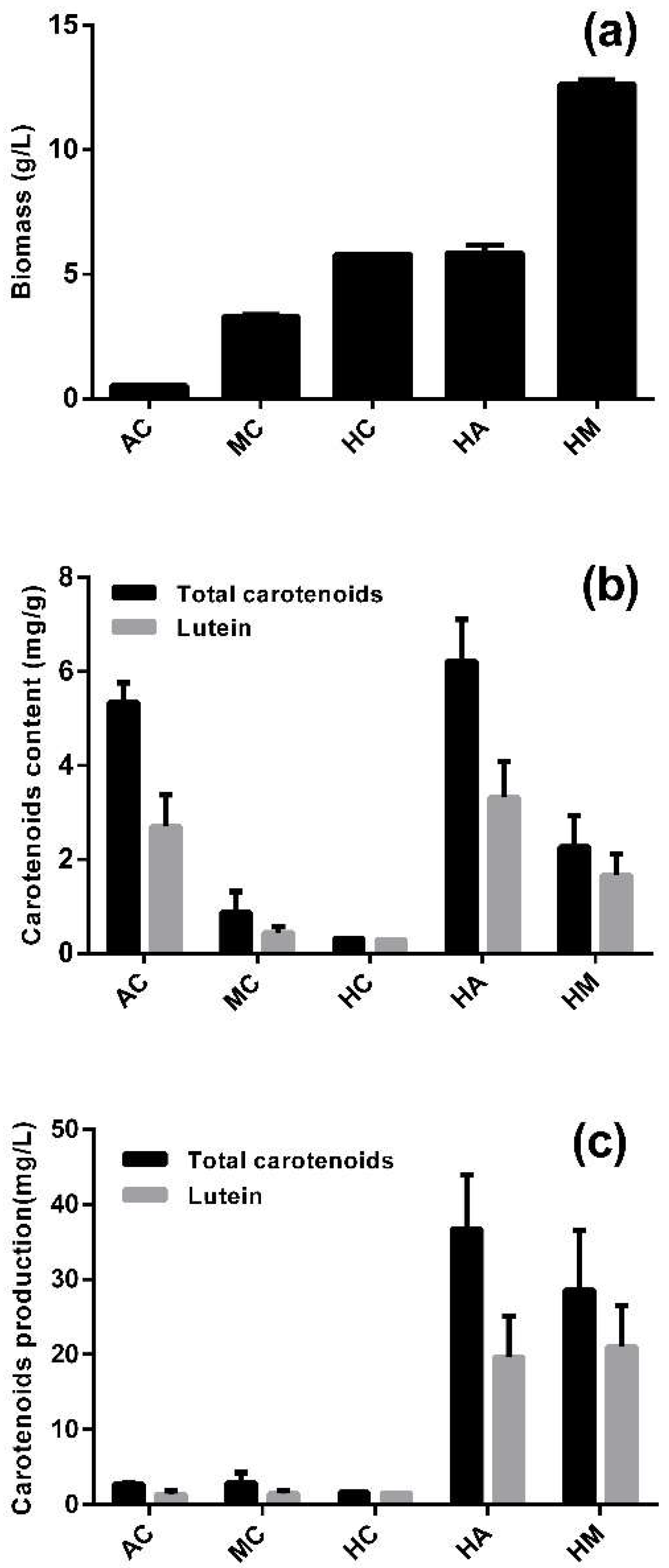
2.1. Lutein Lccumulation in Different Culture Modes
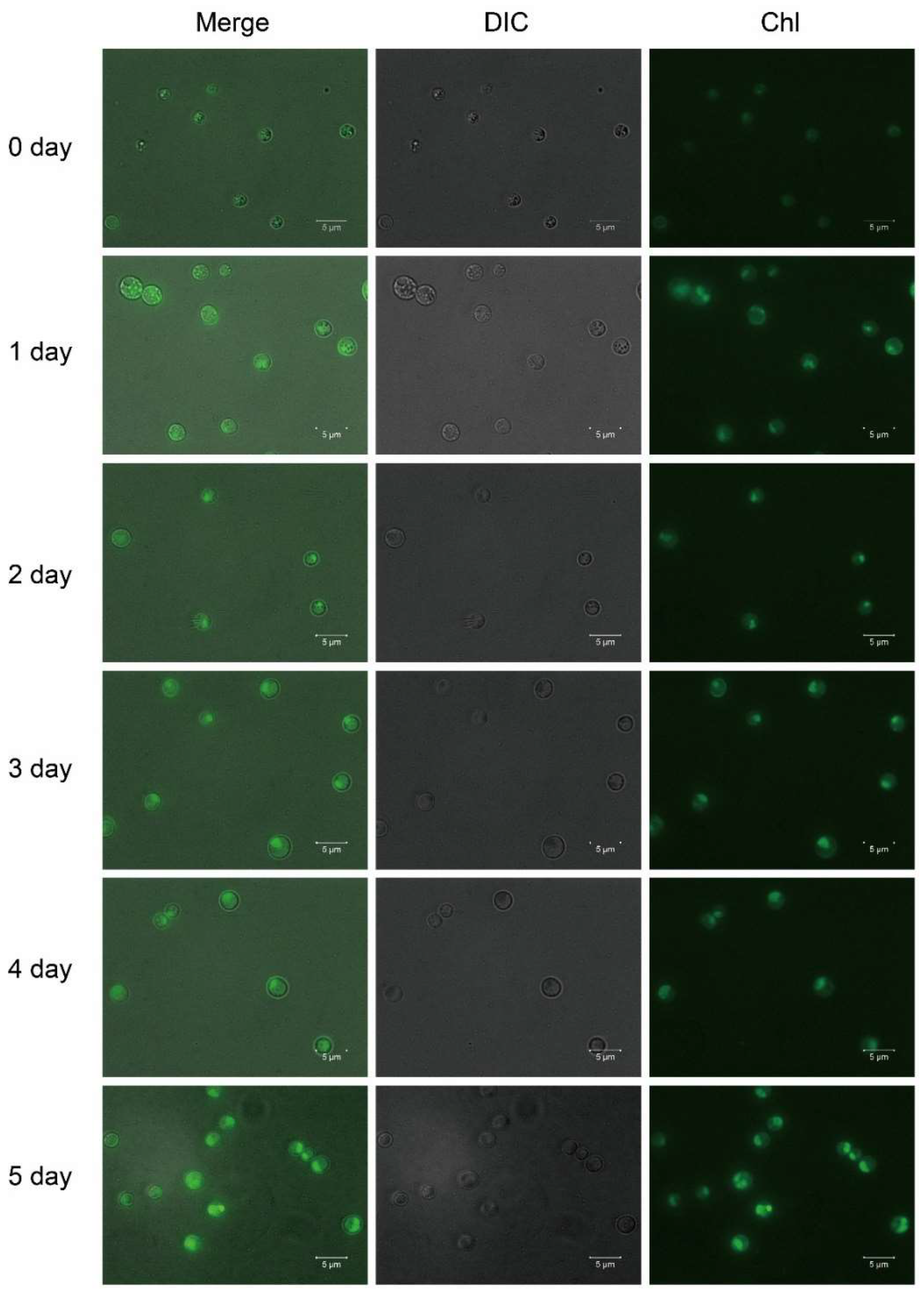
2.2. Lutein Accumulation in the Heterotrophy-Photoautotrophy Culture Mode
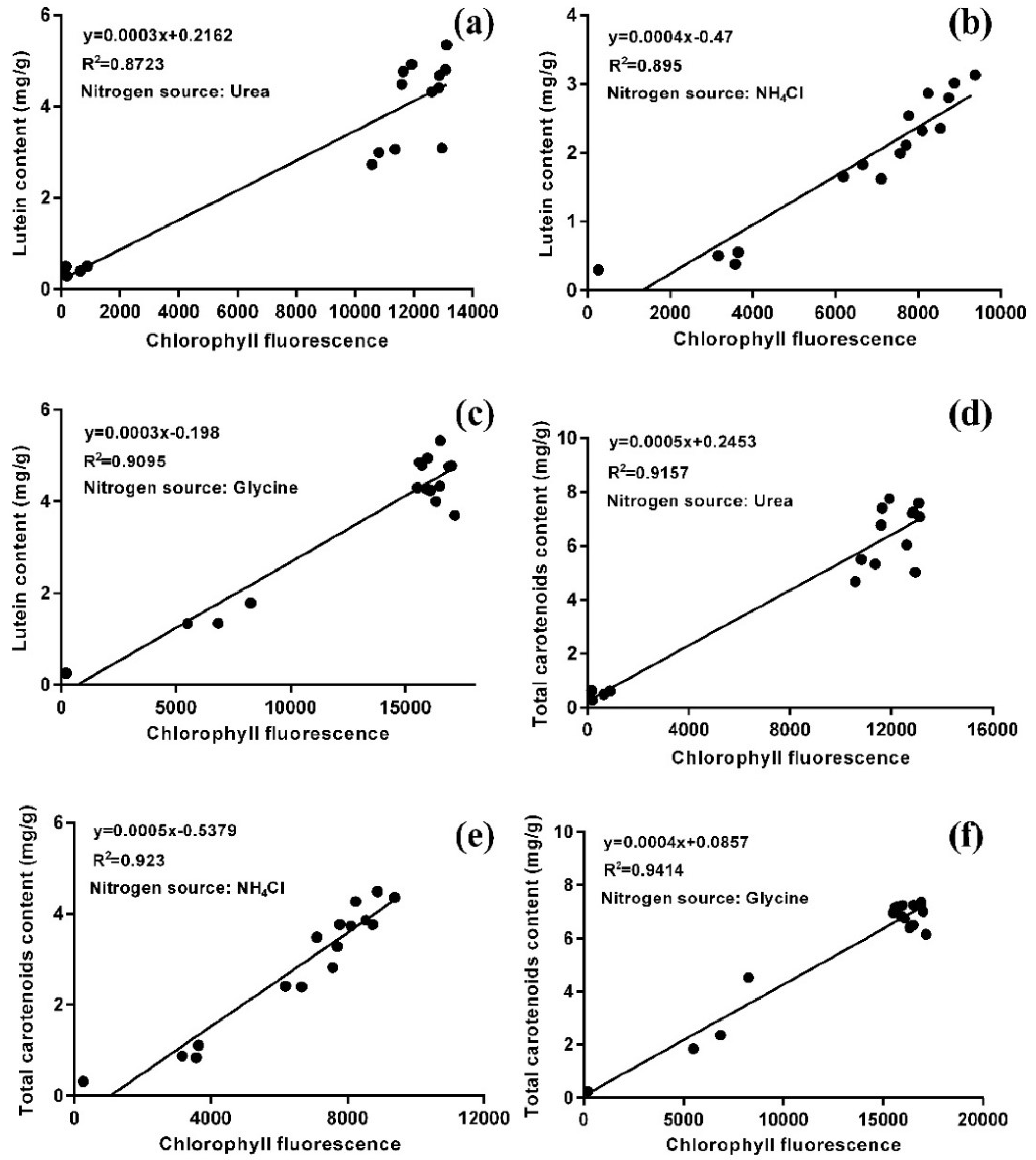
2.3. Lutein Biosynthesis and Chloroplast Regeneration under Nitrogen-Replete Conditions
2.4. Comparative Transcriptomic Analysis of the Genes Involved in Lutein Biosynthesis and Photosynthesis
3. Materials and Methods
3.1. Strain and Culture Conditions
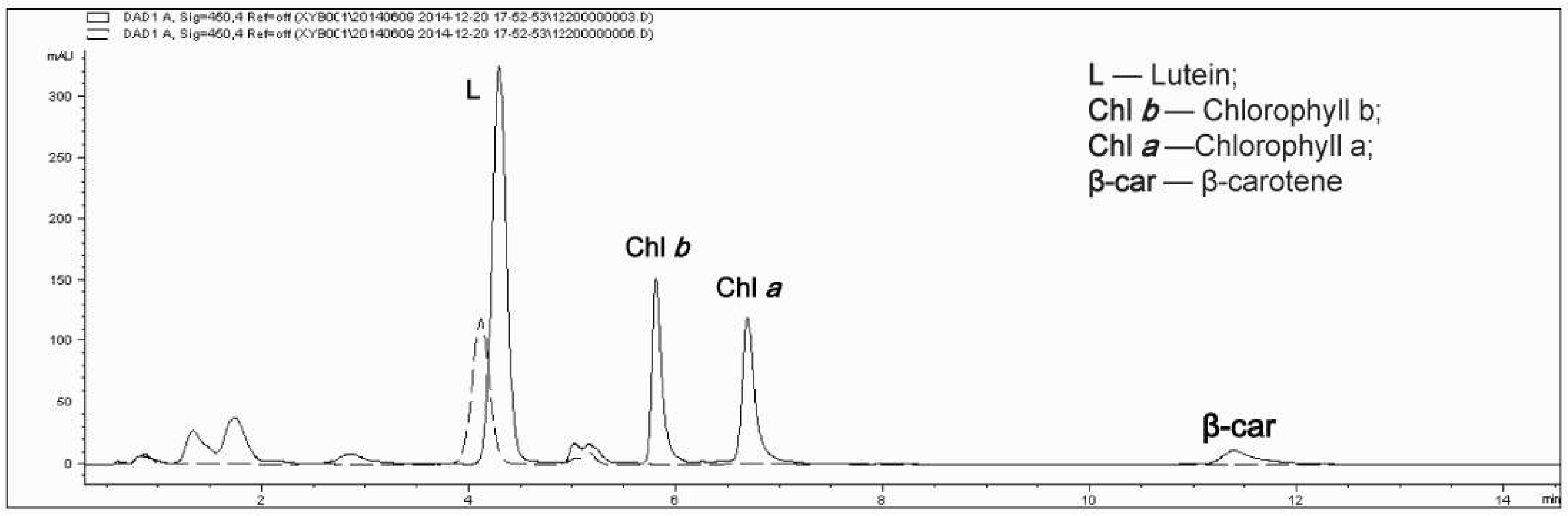
3.2. Determination of Biomass, Total Carotenoids and Lutein Content
3.3. RNA Extraction, Library Preparation, Sequencing and Differential Gene Expression Analysis
3.4. Fluorescence Microscope Analysis
3.5. Flow Cytometry Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chan, M.C.; Ho, S.H.; Lee, D.J.; Chen, C.Y.; Huang, C.C.; Chang, J.S. Characterization, extraction and purification of lutein produced by an indigenous microalga Scenedesmus obliquus CNW-N. Biochem. Eng. J. 2013, 78, 24–31. [Google Scholar] [CrossRef]
- Vidhyavathi, R.; Venkatachalam, L.; Sarada, R.; Ravishankar, G.A. Regulation of carotenoid biosynthetic genes expression and carotenoid accumulation in the green alga. Haematococcus pluvialis under nutrient stress conditions. J. Exp. Bot. 2008, 59, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.X.; Wang, J.X.; Shi, M.L.; Niu, X.F.; Yu, X.H.; Gao, L.J.; Zhang, X.Q.; Chen, L.; Zhang, W.W. Metabolomic and network analysis of astaxanthin-producing Haematococcus pluvialis under various stress conditions. Bioresour. Technol. 2014, 170, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, M.; Moreno, J.; Manzano, J.C.; Florencio, F.J.; Guerrero, M.G. Production of Dunaliella salina biomass rich in 9-cis-beta-carotene and lutein in a closed tubular photobioreactor. J. Biotechnol. 2005, 115, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Moreno, J.; Rodriguez, H.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: factors determining lutein accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef]
- Sujak, A.; Gabrielska, J.; Grudzinski, W.; Borc, R.; Mazurek, P.; Gruszecki, W.I. Lutein and zeaxanthin as protectors of lipid membranes against oxidative damage: The structural aspects. Arch. Biochem. Biophys. 1999, 371, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Lidebjer, C.; Leanderson, P.; Ernerudh, J.; Jonasson, L. Low plasma levels of oxygenated carotenoids in patients with coronary artery disease. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Moeller, S.M.; Voland, R.; Tinker, L.; Blodi, B.A.; Klein, M.L.; Gehrs, K.M.; Johnson, E.J.; Snodderly, D.M.; Wallace, R.B.; Chappell, R.J.; et al. Associations between age-related nuclear cataract and lutein and zeaxanthin in the diet and serum in the Carotenoids in the Age-Related Eye Disease Study (CAREDS), an ancillary study of the women’s health initiative. Arch. Ophthalmol. 2008, 126, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Lin, X.M. Effects of lutein and zeaxanthin on aspects of eye health. J. Sci. Food Agric. 2010, 90, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Molnar, P.; Deli, J.; Tanaka, T.; Kann, Y.; Tani, S.; Gyemant, N.; Molnar, J.; Kawase, M. Carotenoids with anti-Helicobacter pylori Activity from Golden Delicious Apple. Phytother. Res. 2010, 24, 644–648. [Google Scholar] [PubMed]
- Graziani, G.; Schiavo, S.; Nicolai, M.A.; Buono, S.; Fogliano, V.; Pinto, G.; Pollio, A. Microalgae as human food: chemical and nutritional characteristics of the thermo-acidophilic microalga Galdieria sulphuraria. Food Funct. 2013, 4, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Raposo, M.F.D.J.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Sashidhar, B.; Adholeya, A. The isolation and identification of new microalgal strains producing oil and carotenoid simultaneously with biofuel potential. Bioresour. Technol. 2016, 211, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Li, X.F.; Xiang, J.Y.; Wu, Q.Y. High-density fermentation of microalga Chlorella protothecoides in bioreactor for microbio-diesel production. Appl. Microbiol. Biotechnol. 2008, 78, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Araya, B.; Gouveia, L.; Nobre, B.; Reis, A.; Chamy, R.; Poirrier, P. Evaluation of the simultaneous production of lutein and lipids using a vertical alveolar panel bioreactor for three Chlorella species. Algal Res. 2014, 6, 218–222. [Google Scholar] [CrossRef]
- Neijat, M.; Ojekudo, O.; House, J.D. Effect of flaxseed oil and microalgae DHA on the production performance, fatty acids and total lipids of egg yolk and plasma in laying hens. Prostaglandins Leukot. Essent. Fatty Acids 2016, 115, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Brasil, B.D.S.A.F.; de Siqueira, F.G.; Salum, T.F.C.; Zanette, C.M.; Spier, M.R. Microalgae and cyanobacteria as enzyme biofactories. Algal Res. 2017, 25, 76–89. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Dai, J.B.; Wu, Q.Y. Photosynthesis-fermentation hybrid system to produce lipid feedstock for algal biofuel. Environ. Technol. 2013, 34, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Cui, Y.; Gan, Z.B.; Shi, C.L.; Shi, X.M. Isolation and Analysis of the Cppsy Gene and Promoter from Chlorella protothecoides CS-41. Mar. Drugs 2015, 13, 6620–6635. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lu, I.C.; Nagarajan, D.; Chang, C.H.; Ng, I.S.; Lee, D.J.; Chang, J.S. A highly efficient two-stage cultivation strategy for lutein production using heterotrophic culture of Chlorella sorokiniana MB-1-M12. Bioresour. Technol. 2018, 253, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Liu, C.C. Optimization of lutein production with a two-stage mixotrophic cultivation system with Chlorella sorokiniana MB-1. Bioresour. Technol. 2018, 262, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Sun, H.; Zhao, W.Y.; Cheng, K.W.; Chen, F.; Liu, B. A Hetero-Photoautotrophic Two-Stage Cultivation Process for Production of Fucoxanthin by the Marine Diatom Nitzschia laevis. Mar. Drugs 2018, 16, 219. [Google Scholar] [CrossRef] [PubMed]
- Salvini, M.; Bernini, A.; Fambrini, M.; Pugliesi, C. cDNA cloning and expression of the phytoene synthase gene in sunflower. J. Plant Physiol. 2005, 162, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, J.; Linden, H. Regulation of two carotenoid biosynthesis genes coding for phytoene synthase and carotenoid hydroxylase during stress-induced astaxanthin formation in the green alga Haematococcus pluvialis. Plant Physiol. 2001, 125, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Cheng, D.J.; Wang, L.; Gao, J.; Zhao, Q.Y.; Wei, W.; Sun, Y.H. Comparative transcriptomic analysis reveals phenol tolerance mechanism of evolved Chlorella strain. Bioresour. Technol. 2017, 227, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Rodriguez, H.; Moreno, J.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Accumulation of astaxanthin and lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Dai, J.; Wu, Q. Characterization of an ammonium transporter in the oleaginous alga Chlorella protothecoides. Appl. Microbiol. Biotechnol. 2013, 97, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.S.; Lee, D.J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Dai, J.; Wu, Q. Autophagy-like processes are involved in lipid droplet degradation in Auxenochlorella protothecoides during the heterotrophy-autotrophy transition. Front Plant Sci. 2014, 5, 400. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Suga, M.; Kuang, T.; Shen, J.R. Photosynthesis. Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 2015, 348, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Nerantzis, E. Microalgae for high-value compounds and biofuels production: A review with focus on cultivation under stress conditions. Biotechnol. Adv. 2013, 31, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; DellaPenna, D. Defining the primary route for lutein synthesis in plants: The role of Arabidopsis carotenoid beta-ring hydroxylase CYP97A3. Proc. Natl. Acad. Sci. USA 2006, 103, 3474–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallin, E.I.; Guo, K.; Akerlund, H.E. Violaxanthin de-epoxidase disulphides and their role in activity and thermal stability. Photosynth. Res. 2015, 124, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coesel, S.; Obornik, M.; Varela, J.; Falciatore, A.; Bowler, C. Evolutionary Origins and Functions of the Carotenoid Biosynthetic Pathway in Marine Diatoms. PLoS ONE 2008, 3, e2896. [Google Scholar] [CrossRef] [PubMed]
- Jahns, P.; Latowski, D.; Strzalka, K. Mechanism and regulation of the violaxanthin cycle: The role of antenna proteins and membrane lipids. Biochim. Biophys. Acta 2009, 1787, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid Metabolism in Plants: The Role of Plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M. Carotenoid oxidation products as stress signals in plants. Plant J. 2014, 79, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Xiong, W.; Dai, J.; Wu, Q. Genome-Based Metabolic Mapping and 13C Flux Analysis Reveal Systematic Properties of an Oleaginous Microalga Chlorella protothecoides. Plant Physiol. 2015, 167, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.L.; Brackenridge, A.E.; Vivier, M.A.; Oberholster, A. High-performance liquid chromatography profiling of the major carotenoids in Arabidopsis thaliana leaf tissue. J. Chromatogr. A 2006, 1121, 83–91. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Dai, J.; Wu, Q. Identification of Sporopollenin as the Outer Layer of Cell Wall in Microalga Chlorella protothecoides. Front. Microbiol. 2016, 7, 1047. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Enzyme | log2(0–3) | log2(0–6) | log2(0–12) | log2(0–24) | log2(0–72) |
|---|---|---|---|---|---|---|
| PsbB | Photosystem II CP47 chlorophyll apoprotein | −2.91 | −3.35 | −2.87 | −1.84 | NS |
| −0.71 | −2.44 | NS | −0.71 | −2.31 | ||
| PsbE | Photosystem II cytochrome b559 subunit alpha | −0.21 | −0.01 | −0.24 | −1.42 | NS |
| PsbH | Photosystem II PsbH protein | −1.52 | NS | −0.17 | NS | −0.85 |
| PsbO | Photosystem II oxygen-evolving enhancer protein 1 | −4.64 | −2.98 | −2.79 | −4.90 | −3.92 |
| PsbP | Photosystem II oxygen-evolving enhancer protein 2 | −0.52 | −0.21 | −1.35 | −2.19 | −1.48 |
| PsbQ | Photosystem II oxygen-evolving enhancer protein 3 | −3.93 | −2.52 | −1.51 | −2.07 | −1.62 |
| PsbR | Photosystem II 10kDa protein | −2.27 | −2.42 | −1.60 | −2.90 | −2.70 |
| PsbS | Photosystem II 22kDa protein | −8.43 | −8.01 | −7.43 | −8.14 | −7.73 |
| Psb27 | Photosystem II Psb27 protein | −4.80 | −1.67 | −2.18 | −2.11 | −1.56 |
| Psb28 | Photosystem II 13kDa protein | −0.41 | −1.36 | −3.21 | −2.77 | −2.13 |
| PsaC | Photosystem I subunit VII | NS | NS | NS | NS | NS |
| PsaD | Photosystem I subunit II | −2.32 | −1.88 | −1.81 | −2.15 | −2.45 |
| PsaE | Photosystem I subunit IV | −1.78 | −1.07 | −0.91 | −2.51 | −1.48 |
| PsaF | Photosystem I subunit III | −1.98 | −1.68 | −1.14 | −2.84 | −2.48 |
| PsaG | Photosystem I subunit V | −4.33 | −2.43 | −1.91 | −5.10 | −3.93 |
| PsaK | Photosystem I subunit X | −4.49 | −2.03 | −1.30 | −3.40 | −3.12 |
| PsaL | Photosystem I subunit XI | −2.15 | −1.98 | −1.96 | −4.40 | −4.21 |
| PsaN | Photosystem I subunit PsaN | −4.60 | −3.63 | −3.05 | −4.78 | −3.56 |
| PsaO | Photosystem I subunit PsaO | −2.82 | −1.41 | −0.03 | −1.76 | −1.22 |
| PetC | Cytochrome b6-f complex iron-sulfur subunit | −0.89 | −1.36 | −0.95 | −1.97 | −2.11 |
| PetN | Cytochrome b6-f complex subunit 8 | 1.31 | 1.31 | 1.29 | 0.34 | −1.66 |
| PetE | Plastocyanin | −6.16 | −6.73 | −5.35 | −6.14 | −5.77 |
| PetF | Ferredoxin | −1.23 | −1.04 | −1.64 | −2.37 | −3.47 |
| PetH | Ferredoxin--NADP+ reductase | 0.66 | 0.47 | 0.12 | 0.13 | −1.40 |
| LHCA1 | Light-harvesting complex I chlorophyll a/b binding protein 1 | −6.22 | −3.93 | −2.76 | −5.95 | −5.77 |
| LHCA3 | Light-harvesting complex I chlorophyll a/b binding protein 3 | −3.44 | −2.02 | −0.42 | −2.58 | −3.02 |
| LHCA5 | Light-harvesting complex I chlorophyll a/b binding protein 5 | −6.94 | −6.36 | −6.46 | −8.07 | −4.88 |
| −4.49 | −2.98 | −1.29 | −3.14 | −3.58 | ||
| LHCB3 | Light-harvesting complex II chlorophyll a/b binding protein 3 | −0.76 | −0.65 | −0.81 | −3.50 | −7.99 |
| LHCB4 | Light-harvesting complex II chlorophyll a/b binding protein 4 | −2.60 | −1.58 | −1.46 | −3.14 | −2.86 |
| LHCB5 | Light-harvesting complex II chlorophyll a/b binding protein 5 | −3.33 | −2.81 | −1.06 | −3.07 | −5.82 |
| PDS | 15-cis-phytoene desaturase | −0.71 | −1.01 | −1.13 | −1.60 | −1.19 |
| ZDS | Zeta-carotene desaturase | −1.51 | −1.36 | −1.43 | −2.18 | −1.46 |
| ZEP | Zeaxanthin epoxidase | −5.34 | −5.22 | −1.69 | −3.00 | −1.50 |
| VDE | Violaxanthin de-epoxidase | −6.90 | −7.60 | −5.30 | −5.49 | −3.87 |
| CrtL-e | Lycopene epsilon-cyclase | −1.46 | −0.29 | −0.80 | −1.61 | −1.24 |
| CrtL-b | Lycopene beta-cyclase | −3.05 | −2.00 | −2.03 | −2.46 | −1.46 |
| CrtR-b | Beta-carotene 3-hydroxylase | 1.06 | 0.41 | −0.14 | −1.29 | −0.63 |
| LUT1 | Carotene epsilon-monooxygenase | −2.73 | −0.78 | −0.54 | −1.58 | −1.23 |
| LUT5 | Beta-ring hydroxylase | 2.11 | 1.84 | 2.19 | 0.96 | −0.27 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; He, X.; Ma, Q.; Lu, Y.; Bai, F.; Dai, J.; Wu, Q. Photosynthetic Accumulation of Lutein in Auxenochlorella protothecoides after Heterotrophic Growth. Mar. Drugs 2018, 16, 283. https://doi.org/10.3390/md16080283
Xiao Y, He X, Ma Q, Lu Y, Bai F, Dai J, Wu Q. Photosynthetic Accumulation of Lutein in Auxenochlorella protothecoides after Heterotrophic Growth. Marine Drugs. 2018; 16(8):283. https://doi.org/10.3390/md16080283
Chicago/Turabian StyleXiao, Yibo, Xi He, Qi Ma, Yue Lu, Fan Bai, Junbiao Dai, and Qingyu Wu. 2018. "Photosynthetic Accumulation of Lutein in Auxenochlorella protothecoides after Heterotrophic Growth" Marine Drugs 16, no. 8: 283. https://doi.org/10.3390/md16080283
APA StyleXiao, Y., He, X., Ma, Q., Lu, Y., Bai, F., Dai, J., & Wu, Q. (2018). Photosynthetic Accumulation of Lutein in Auxenochlorella protothecoides after Heterotrophic Growth. Marine Drugs, 16(8), 283. https://doi.org/10.3390/md16080283