Cultivation of Sponge-Associated Bacteria from Agelas sventres and Xestospongia muta Collected from Different Depths

 ,
,  and
and
Abstract
:1. Introduction
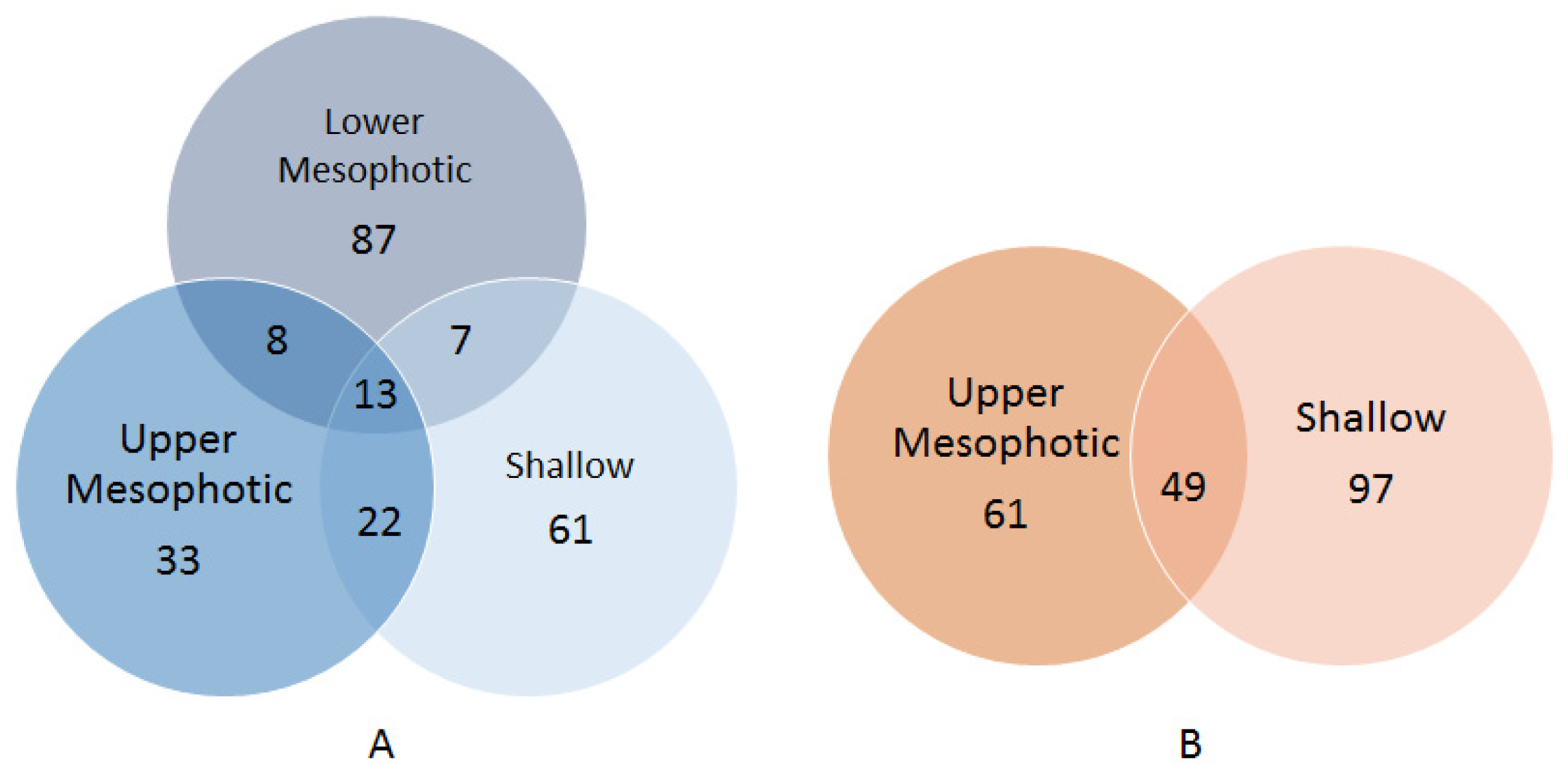
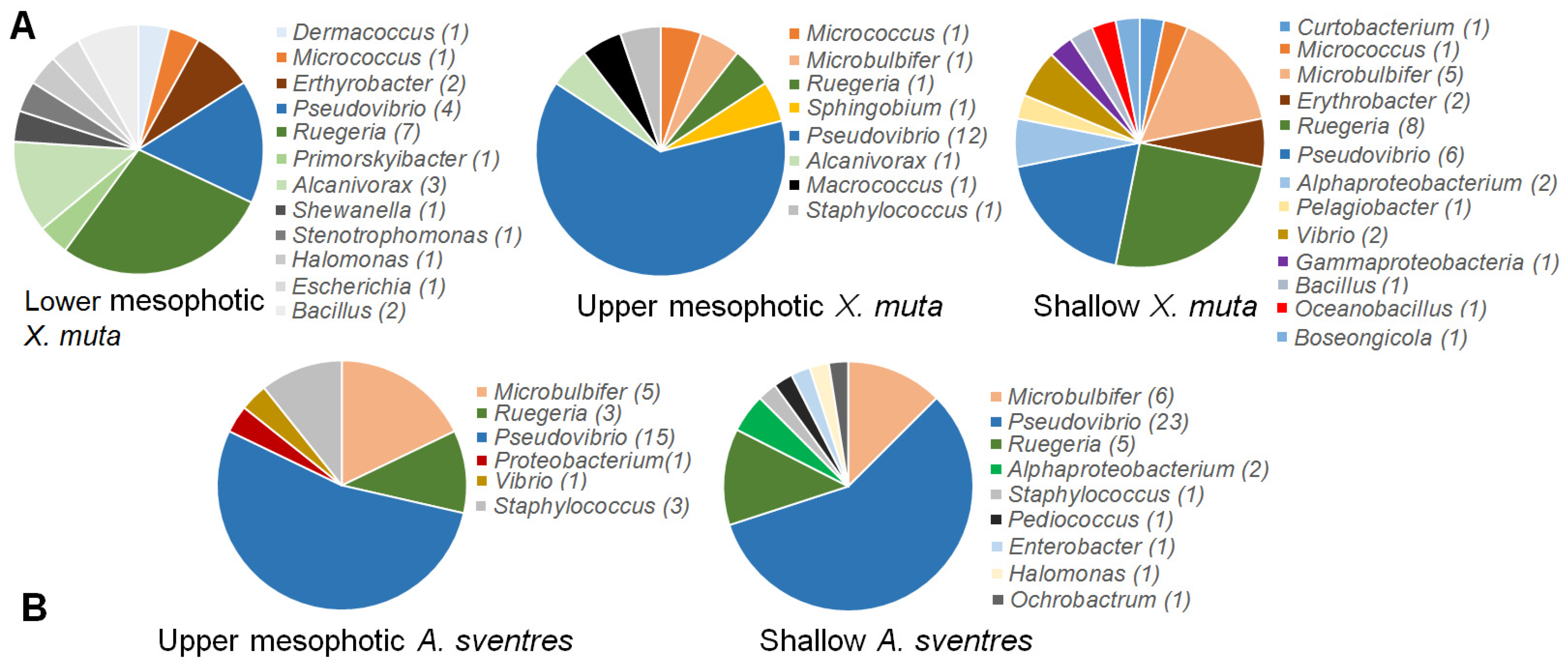
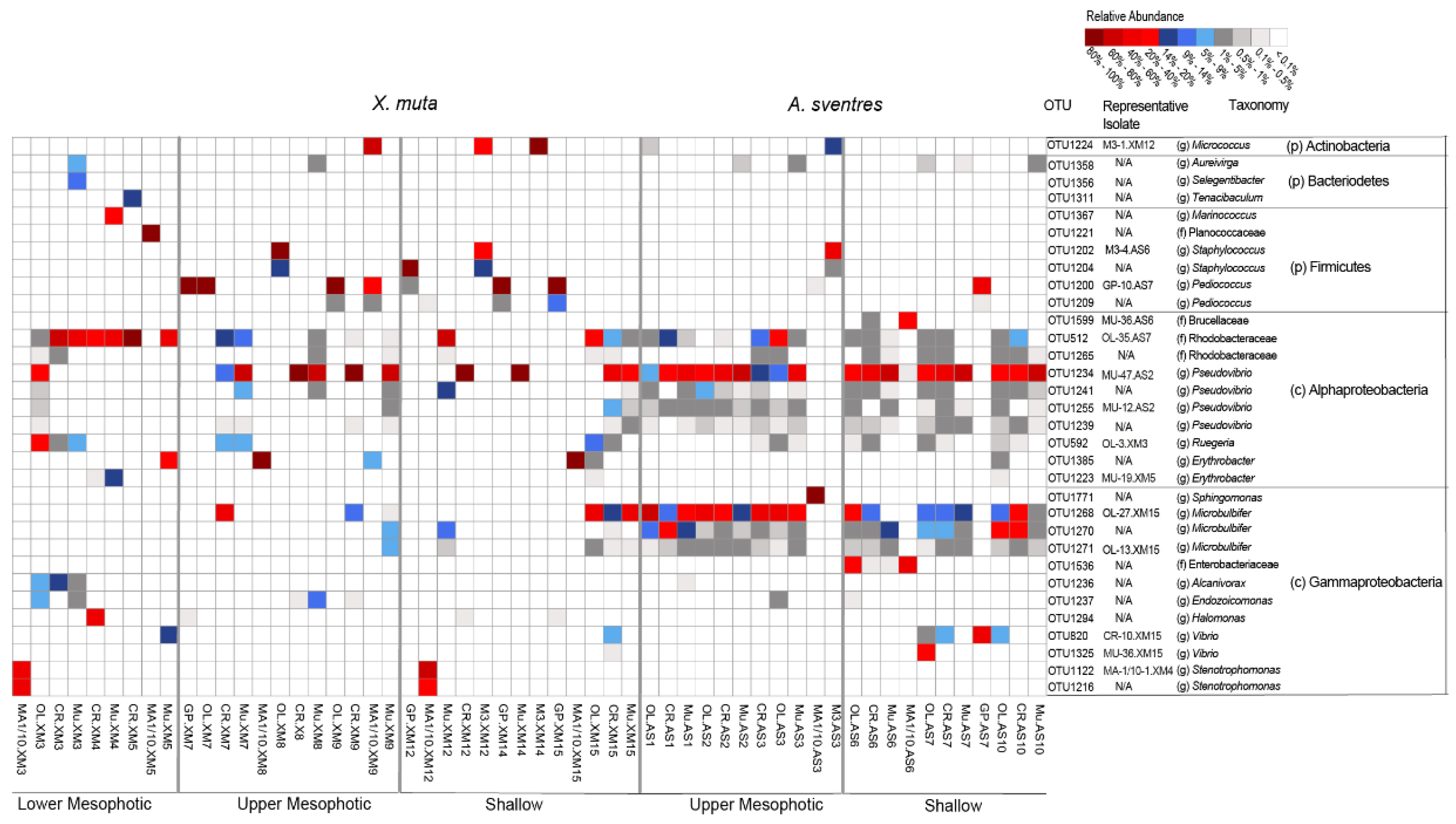
2. Results
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Cultivation Conditions
4.3. DNA Extraction
4.4. Prokaryotic Community Profiling Using 16S rRNA Gene Amplicon Sequencing
4.5. Sequence Data Processing
4.6. Prokaryotic Diversity Analyses
4.7. Regrowth and Identification of Picked Isolates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [PubMed]
- Sipkema, D.; Franssen, M.C.R.; Osinga, R.; Tramper, J.; Wijffels, R.H. Marine sponges as pharmacy. Mar. Biotechnol. 2005, 7, 142–162. [Google Scholar] [CrossRef] [PubMed]
- Laport, M.S.; Santos, O.C.S.; Muricy, G. Marine sponges: Potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol. 2009, 10, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.L.; Peraud, O.; Kasanah, N.; Sims, J.W.; Kothalawala, N.; Anderson, M.A.; Abbas, S.H.; Rao, K.V.; Jupally, V.R.; Kelly, M.; et al. An analysis of the sponge Acanthostrongylophora igens’ microbiome yields an actinomycete that produces the natural product manzamine A. Front. Mar. Sci 2014, 1, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.C.; Mori, T.; Ruckert, C.; Uria, A.R.; Helf, M.J.; Takada, K.; Gernert, C.; Steffens, U.A.E.; Heycke, N.; Schmitt, S.; et al. An environmental bacterial taxon with a large and distinct metabolic repertoire. Nature 2014, 506, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2009, 26, 338–362. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. R 2007, 71, 295–347. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nature Rev. Microbiol. 2012, 10, 641–675. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.M.; Saito, K.; Tal, Y.; Hill, R.T. Diversity of aerobic and anaerobic ammonia-oxidizing bacteria in marine sponges. ISME J. 2010, 4, 38–48. [Google Scholar] [CrossRef]
- Hoffmann, F.; Radax, R.; Woebken, D.; Holtappels, M.; Lavik, G.; Rapp, H.T.; Schläppy, M.-L.; Schleper, C.; Kuypers, M.M.M. Complex nitrogen cycling in the sponge Geodia barretti. Environ. Microbiol. 2009, 11, 2228–2243. [Google Scholar] [CrossRef]
- Tian, R.-M.; Wang, Y.; Bougouffa, S.; Gao, Z.-M.; Cai, L.; Bajic, V.; Qian, P.-Y. Genomic analysis reveals versatile heterotrophic capacity of a potentially symbiotic sulfur-oxidizing bacterium in sponge. Environ. Microbiol. 2014, 16, 3548–3561. [Google Scholar] [CrossRef] [PubMed]
- Lavy, A.; Keren, R.; Yu, K.; Thomas, B.C.; Alvarez-Cohen, L.; Banfield, J.F.; Ilan, M. A novel Chromatiales bacterium is a potential sulfide oxidizer in multiple orders of marine sponges. Environ. Microbiol. 2018, 20, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Blasiak, L.C.; Karolin, J.O.; Powell, R.J.; Geddes, C.D.; Hill, R.T. Phosphorus sequestration in the form of polyphosphate by microbial symbionts in marine sponges. Proc. Natl. Acad. Sci. USA 2015, 112, 4381–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schippers, K.J.; Sipkema, D.; Osinga, R.; Smidt, H.; Pomponi, S.A.; Martens, D.E.; Wijffels, R.H. Cultivation of sponges, sponge cells and symbionts: Achievements and future prospects. Adv. Mar. Biol. 2012, 62, 273–337. [Google Scholar] [PubMed]
- Joint, I.; Muhling, M.; Querellou, J. Culturing marine bacteria—An essential prerequisite for biodiscovery. Microb. Biotechnol. 2010, 3, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Sipkema, D.; Schippers, K.; Maalcke, W.J.; Yang, Y.; Salim, S.; Blanch, H.W. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (gellius) sp. Appl. Environ. Microb 2011, 77, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Santos, O.C.S.; Pontes, P.V.M.L.; Santos, J.F.M.; Muricy, G.; Giambiagi-deMarval, M.; Laport, M.S. Isolation, characterization and phylogeny of sponge-associated bacteria with antimicrobial activities from Brazil. Res. Microbiol. 2010, 161, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Schmid, M.; Wagner, M.; Fieseler, L.; Gernert, C.; Hacker, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001, 35, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the mediterranean sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [CrossRef]
- Matobole, R.M.; van Zyl, L.J.; Parker-Nance, S.; Davies-Coleman, M.T.; Trindade, M. Antibacterial activities of bacteria isolated from the marine sponges Isodictya compressa and Higginsia bidentifera collected from Algoa Bay, South Africa. Mar. Drugs 2017, 15, 47. [Google Scholar] [CrossRef]
- Subramani, R.; Sipkema, D. Marine rare Actinomycetes: A promising source of structurally diverse and unique novel natural products. Mar. Drugs. 2019, 17, 249. [Google Scholar] [CrossRef] [PubMed]
- Indraningrat, A.A.G.; Smidt, H.; Sipkema, D. Bioprospecting sponge-associated microbes for antimicrobial compounds. Mar. Drugs 2016, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.B.; Gao, X.M. Characterizing the bacterial associates of three Caribbean sponges along a gradient from shallow to mesophotic depths. FEMS Microbiol. Ecol. 2013, 85, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Morrow, K.M.; Fiore, C.L.; Lesser, M.P. Environmental drivers of microbial community shifts in the giant barrel sponge, Xestospongia muta, over a shallow to mesophotic depth gradient. Environ. Microbiol. 2016, 18, 2025–2038. [Google Scholar] [CrossRef] [PubMed]
- Steinert, G.; Taylor, M.W.; Deines, P.; Simister, R.L.; de Voogd, N.J.; Hoggard, M.; Schupp, P.J. In four shallow and mesophotic tropical reef sponges from Guam the microbial community largely depends on host identity. PeerJ. 2016, 4, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Beazley, L.I.; Kenchington, E.L.; Murillo, F.J.; Sacau, M.D. Deep-sea sponge grounds enhance diversity and abundance of epibenthic megafauna in the Northwest Atlantic. ICES J. Mar. Sci 2013, 70, 1471–1490. [Google Scholar] [CrossRef]
- Thoms, C.; Schupp, P. Biotechnological potential of marine sponges and their associated bacteria as producers of new pharmaceuticals (part II). J. Int. Biotechnol. Law 2005, 2, 257–264. [Google Scholar] [CrossRef]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Bjork, J.R.; Easson, C.; Astudillo-Garcia, C.; Olson, J.B.; Erwin, P.M.; Lopez-Legentil, S.; Luter, H.; et al. Diversity, structure and convergent evolution of the global sponge microbiome. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Olson, J.B.; Kellogg, C.A. Microbial ecology of corals, sponges, and algae in mesophotic coral environments. FEMS Microbiol. Ecol. 2010, 73, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Slattery, M.; Lesser, M.P.; Brazeau, D.; Stokes, M.D.; Leichter, J.J. Connectivity and stability of mesophotic coral reefs. J. Exp. Mar. Biol. Ecol. 2011, 408, 32–41. [Google Scholar] [CrossRef]
- Webster, N.S.; Taylor, M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Borchert, E.; Jackson, S.A.; O’Gara, F.; Dobson, A.D.W. Diversity of natural product biosynthetic genes in the microbiome of the deep sea sponges Inflatella pellicula, Poecillastra compressa, and Stelletta normani. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sipkema, D. Marine biotechnology: Diving deeper for drugs. Microb. Biotechnol. 2017, 10, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Parra-Velandia, F.J.; Zea, S.; Van Soest, R.W.M. Reef sponges of the genus Agelas (Porifera: Demospongiae) from the Greater Caribbean. Zootaxa 2014, 3794, 301–343. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, R.; Meesters, E.H.W.G.; Becking, L.E. Deep-water sponges (Porifera) from Bonaire and Klein Curaçao, Southern Caribbean. Zootaxa 2014, 3878, 404–443. [Google Scholar] [CrossRef]
- Zhou, X.F.; Xu, T.H.; Yang, X.W.; Huang, R.M.; Yang, B.; Tang, L.; Liu, Y.H. Chemical and biological aspects of marine sponges of the genus Xestospongia. Chem. Biodiv 2010, 7, 2201–2227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Dong, M.L.; Chen, J.W.; Wang, H.; Tenney, K.; Crews, P. Bioactive secondary metabolites from the marine sponge genus Agelas. Mar. Drugs 2017, 15, 351. [Google Scholar] [CrossRef] [PubMed]
- Montalvo, N.F.; Davis, J.; Vicente, J.; Pittiglio, R.; Ravel, J.; Hill, R.T. Integration of culture-based and molecular analysis of a complex sponge-associated bacterial community. Plos One 2014, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Indraningrat, A.A.G.; Steinert, G.; Becking, L.E.; Mueller, B.; de Goeij, J.; Smidt, H.; Sipkema, D. Depth affects sponge prokaryotic communities and their antimicrobial activities in two Demosponges, Xestospongia muta and Agelas sventres. Unpublished work. Manuscript in preparation.
- Versluis, D.; McPherson, K.; Passel, M.; Smidt, H.; Sipkema, D. Recovery of previously uncultured bacterial genera from three Mediterranean sponges. Mar. Biotechnol. 2017, 19, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, C.C.P.; Cardinale, M.; Cucio, A.C.B.; Esteves, A.I.S.; Berg, G.; Xavier, J.R.; Cox, C.J.; Costa, R. Effects of sample handling and cultivation bias on the specificity of bacterial communities in keratose marine sponges. Front. Microbiol. 2014, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Graça, A.P.; Bondoso, J.; Gaspar, H.; Xavier, J.R.; Monteiro, M.C.; de la Cruz, M.; Oves-Costales, D.; Vicente, F.; Lage, O.M. Antimicrobial activity of heterotrophic bacterial communities from the marine sponge Erylus discophorus (Astrophorida, Geodiidae). PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chaib De Mares, M.; Jiménez, D.J.; Palladino, G.; Gutleben, J.; Lebrun, L.A.; Muller, E.E.L.; Wilmes, P.; Sipkema, D.; van Elsas, J.D. Expressed protein profile of a Tectomicrobium and other microbial symbionts in the marine sponge Aplysina aerophoba as evidenced by metaproteomics. Sci. Rep. 2018, 8, 11795. [Google Scholar] [CrossRef]
- Karimi, E.; Slaby, B.M.; Soares, A.R.; Blom, J.; Hentschel, U.; Costa, R. Metagenomic binning reveals versatile nutrient cycling and distinct adaptive features in alphaproteobacterial symbionts of marine sponges. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Ramos, M.; Gonçalves, J.M.S.; Xavier, J.R.; Reis, M.P.; Costa, R. Comparative metagenomics reveals the distinctive adaptive features of the Spongia officinalis endosymbiotic consortium. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Gutleben, J.; Chaib De Mares, M.; van Elsas, J.D.; Smidt, H.; Overmann, J.; Sipkema, D. The multi-omics promise in context: From sequence to microbial isolate Crit. Rev. Microbiol. 2018, 44, 212–229. [Google Scholar] [CrossRef]
- Slaby, B.M.; Hackl, T.; Horn, H.; Bayer, K.; Hentschel, U. Metagenomic binning of a marine sponge microbiome reveals unity in defense but metabolic specialization. ISME J. 2017, 11, 2465–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Q.; Wang, J.; Gu, L.; Zheng, T.; Shao, Z. Alcanivorax marinus sp. nov., isolated from deep-sea water. Int. J. Syst. Evol. Microbiol. 2013, 63, 4428–4432. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Barrett, N.H.; McCarthy, P.J. Draft genome sequence of deep-sea Alteromonas sp. strain V450 isolated from the marine sponge Leiodermatium sp. Genome. Announc 2017, 5, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Kyoung Kwon, K.; Hye Oh, J.; Yang, S.-H.; Seo, H.-S.; Lee, J.-H. Alcanivorax gelatiniphagus sp. nov., a marine bacterium isolated from tidal flat sediments enriched with crude oil. Int. J. Syst. Evol. Microbiol. 2015, 65, 2204–2208. [Google Scholar] [CrossRef]
- Lesser, M.P.; Slattery, M.; Leichter, J.J. Ecology of mesophotic coral reefs. J. Exp. Mar. Biol. Ecol. 2009, 375, 1–8. [Google Scholar] [CrossRef]
- Liu, Q.; Fang, J.; Li, J.; Zhang, L.; Xie, B.-B.; Chen, X.-L.; Zhang, Y.-Z. Depth-resolved variations of cultivable bacteria and their extracellular enzymes in the water column of the New Britain Trench. Front. Microbiol. 2018, 9, 135. [Google Scholar] [CrossRef] [PubMed]
- Kai, W.; Peisheng, Y.; Rui, M.; Wenwen, J.; Zongze, S. Diversity of culturable bacteria in deep-sea water from the South Atlantic Ocean. Bioengineered 2017, 8, 572–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enticknap, J.J.; Kelly, M.; Peraud, O.; Hill, R.T. Characterization of a culturable Alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. App. Env. Microbiol. 2006, 72, 3724–3732. [Google Scholar] [CrossRef] [PubMed]
- Romano, S. Ecology and biotechnological potential of bacteria belonging to the genus. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Margassery, L.M.; Kennedy, J.; O’Gara, F.; Dobson, A.D.; Morrissey, J.P. Diversity and antibacterial activity of bacteria isolated from the coastal marine sponges Amphilectus fucorum and Eurypon major. Lett. App. Microbiol. 2012, 55, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, C.; Kearns, P.; Evans-Illidge, E.; Kurtböke, D. Diversity and bioactivity of marine bacteria associated with the sponges Candidaspongia flabellata and Rhopaloeides odorabile from the Great Barrier Reef in Australia. Diversity 2017, 9, 39. [Google Scholar] [CrossRef]
- Mohamed, N.M.; Cicirelli, E.M.; Kan, J.; Chen, F.; Fuqua, C.; Hill, R.T. Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ. Microbiol. 2008, 10, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Zan, J.; Cicirelli, E.M.; Mohamed, N.M.; Sibhatu, H.; Kroll, S.; Choi, O.; Uhlson, C.L.; Wysoczynski, C.L.; Murphy, R.C.; Churchill, M.E.A.; et al. A complex LuxR–LuxI type quorum sensing network in a roseobacterial marine sponge symbiont activates flagellar motility and inhibits biofilm formation. Mol. Microbiol. 2012, 85, 916–933. [Google Scholar] [CrossRef] [PubMed]
- Quévrain, E.; Domart-Coulon, I.; Pernice, M.; Bourguet-Kondracki, M.-L. Novel natural parabens produced by a Microbulbifer bacterium in its calcareous sponge host Leuconia nivea. Environ. Microbiol. 2009, 11, 1527–1539. [Google Scholar] [CrossRef]
- Esteves, A.I.S.; Amer, N.; Nguyen, M.; Thomas, T. Sample processing impacts the viability and cultivability of the sponge microbiome. Front. Microbiol. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Munroe, S.; Martens, D.E.; Sipkema, D.; Pomponi, S.A. Comparison of cryopreservation techniques for cells of the marine sponge Dysidea etheria. Cryoletters 2018, 39, 269–278. [Google Scholar] [PubMed]
- Olson, J.B.; Lord, C.C.; McCarthy, P.J. Improved Recoverability of Microbial Colonies from Marine Sponge Samples. Microbiol. Ecol. 2000, 40, 139–147. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Jin, Y.; Jin, M.; Yu, X. A comparative study on the phylogenetic diversity of culturable actinobacteria isolated from five marine sponge species. Antonie van Leeuwenhoek 2008, 93, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Santavy, D.L.; Colwell, R.R. Comparison of bacterial communities associated with the Caribbean Sclerosponge Ceratoporella nicholsoni and ambient seawater. Mar. Ecol. Prog. Ser. 1990, 67, 73–82. [Google Scholar] [CrossRef]
- Olson, J.B.; McCarthy, P.J. Associated bacterial communities of two deep-water sponges. Aquat. Microb. Ecol. 2005, 39, 47–55. [Google Scholar] [CrossRef]
- LibreTexts. Available online: https://bio.libretexts.org/Ancillary_Materials/Laboratory_Experiments/Microbiology_Labs/Microbiology_Labs_I/08%3A_Bacterial_Colony_Morphology (accessed on 30 April 2016).
- Van Lingen, H.J.; Edwards, J.E.; Vaidya, J.D.; van Gastelen, S.; Saccenti, E.; van den Bogert, B.; Bannink, A.; Smidt, H.; Plugge, C.M.; Dijkstra, J. Diurnal dynamics of gaseous and dissolved metabolites and microbiota composition in the bovine rumen. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Ramiro-Garcia, J.; Hermes, G.; Giatsis, C.; Sipkema, D.; Zoetendal, E.; Schaap, P.; Smidt, H. NG-Tax, a highly accurate and validated pipeline for analysis of 16S rRNA amplicons from complex biomes. F1000Research 2016, 5, 1791. [Google Scholar] [CrossRef]
- Dat, T.T.H.; Steinert, G.; Thi Kim Cuc, N.; Smidt, H.; Sipkema, D. Archaeal and bacterial diversity and community composition from 18 phylogenetically divergent sponge species in Vietnam. PeerJ. 2018, 6, 1–23. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic. Acids. Res 2014, 42, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Lahti, L.; Shetty, A.S.; Blake, T.; Salojarvi, J. Microbiome R Package. 2017. Available online: https://bioconductor.org/packages/devel/bioc/html/microbiome.html (accessed on 31 July 2018).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.M.; et al. Vegan: Community Ecology Package. 2018. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 31 July 2018).
- Kolde, R. pheatmap: Pretty Heatmaps. R Package Version 1.0.8. 2015. Available online: https://cran.r-project.org/web/packages/pheatmap/pheatmap.pdf (accessed on 31 July 2018).
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Total number of colonies from X. muta samples | |||||||||||
| Scraping isolates | X. muta lower mesophotic | X. muta upper mesophotic | X. muta shallow | Total per medium | |||||||
| XM3 | XM4 | XM5 | XM7 | XM8 | XM9 | XM12 | XM14 | XM15 | |||
| MA1/10 agar | 2 | 0 | 13 | 0 | 1 | 22 | 1 | 0 | 1 | 40 | |
| M3 agar | 0 | 0 | 0 | 0 | 1 | 0 | 43 | 3 | 0 | 47 | |
| OLIGO agar | 56 | 0 | 0 | 1 | 1 | 82 | 0 | 0 | 24 | 164 | |
| GP agar | 0 | 0 | 0 | 1 | 1 | 0 | 7 | 3 | 0 | 12 | |
| Mucin agar | 87 | 3 | 12 | 66 | 13 | 5 | 5 | 1 | 22 | 214 | |
| Crenarchaeota agar | 61 | 4 | 11 | 51 | 1 | 3 | 2 | 0 | 40 | 173 | |
| Total per sample | 206 | 7 | 36 | 119 | 18 | 112 | 58 | 7 | 87 | 650 | |
| Total per depth | 249 | 249 | 152 | ||||||||
| B. Total number of colonies from A. sventres sample | |||||||||||
| Scraping isolates | A. sventres upper mesophotic | A. sventres shallow | Total per medium | ||||||||
| AS1 | AS2 | AS3 | AS6 | AS7 | AS10 | ||||||
| MA1/10 agar | 3 | 0 | 2 | 6 | 0 | 0 | 11 | ||||
| M3 agar | 0 | 0 | 4 | 1 | 0 | 2 | 7 | ||||
| OLIGO agar | 19 | 513 | 131 | 45 | 69 | 126 | 903 | ||||
| GP agar | 0 | 0 | 0 | 0 | 4 | 0 | 4 | ||||
| Mucin agar | 57 | 377 | 127 | 19 | 53 | 399 | 1032 | ||||
| Crenarchaeota agar | 32 | 668 | 130 | 49 | 63 | 125 | 1087 | ||||
| Total per sample | 111 | 1558 | 394 | 120 | 189 | 652 | 3024 | ||||
| Total per depth | 2063 | 961 | |||||||||
| Samples | Parameter | OTUs | df | PERMANOVA | Betadisper | ||
|---|---|---|---|---|---|---|---|
| R2 | p-Value | F | p-Value | ||||
| All scraped bacterial colonies (excluding inocula of X. muta and A. sventres) | Sponge (X. muta and A. sventres) | 371 | 1 | 0.10 | 0.001 | 23.84 | 0.001 |
| Media type (MA1/10, M3, OLIGO, GP, Mucin, Crenarchaeota) | 371 | 5 | 0.28 | 0.001 | 2.43 | 0.06 | |
| Depth (lower mesophotic, upper mesophotic, shallow) | 371 | 2 | 0.08 | 0.003 | 0.16 | 0.84 | |
| Scraped bacterial colonies of X. muta (excluding inoculum of X. muta) | Depth (lower mesophotic, upper mesophotic and shallow) | 220 | 2 | 0.10 | 0.038 | 0.84 | 0.45 |
| Media type (MA1/10, M3, OLIGO, GP, Mucin, Crenarchaeota) | 220 | 5 | 0.31 | 0.001 | 1.20 | 0.33 | |
| Scraped bacterial colonies of A. sventres (excluding inoculum of A. sventres) | Depth (upper mesophotic and shallow) | 151 | 1 | 0.06 | 0.146 | 0.05 | 0.83 |
| Media type (MA1/10, M3, OLIGO, GP, Mucin, Crenarchaeota) | 151 | 5 | 0.60 | 0.001 | 4.00 | 0.04 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Indraningrat, A.A.G.; Micheller, S.; Runderkamp, M.; Sauerland, I.; Becking, L.E.; Smidt, H.; Sipkema, D. Cultivation of Sponge-Associated Bacteria from Agelas sventres and Xestospongia muta Collected from Different Depths. Mar. Drugs 2019, 17, 578. https://doi.org/10.3390/md17100578
Indraningrat AAG, Micheller S, Runderkamp M, Sauerland I, Becking LE, Smidt H, Sipkema D. Cultivation of Sponge-Associated Bacteria from Agelas sventres and Xestospongia muta Collected from Different Depths. Marine Drugs. 2019; 17(10):578. https://doi.org/10.3390/md17100578
Chicago/Turabian StyleIndraningrat, Anak Agung Gede, Sebastian Micheller, Mandy Runderkamp, Ina Sauerland, Leontine E. Becking, Hauke Smidt, and Detmer Sipkema. 2019. "Cultivation of Sponge-Associated Bacteria from Agelas sventres and Xestospongia muta Collected from Different Depths" Marine Drugs 17, no. 10: 578. https://doi.org/10.3390/md17100578
APA StyleIndraningrat, A. A. G., Micheller, S., Runderkamp, M., Sauerland, I., Becking, L. E., Smidt, H., & Sipkema, D. (2019). Cultivation of Sponge-Associated Bacteria from Agelas sventres and Xestospongia muta Collected from Different Depths. Marine Drugs, 17(10), 578. https://doi.org/10.3390/md17100578