Biotechnological Potential of Bacteria Isolated from the Sea Cucumber Holothuria leucospilota and Stichopus vastus from Lampung, Indonesia

 , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolation, Taxonomic Identification, and Antimicrobial Assay
2.2. Illumina MiSEQ Analysis from the Tissue of Sea Cucumber
2.3. Testing the Effect of Bacterial Extracts on the Infectivity of Hepatitis C Virus (HCV)
2.4. Identification of Putative Compounds from Bioactive Fractions
3. Discussion
4. Materials and Methods
4.1. Isolation of Bacteria
4.2. Identification of Bacteria by 16S rRNA Gene Sanger Sequencing and Construction of Phylogenetic Tree
4.3. Preparation of 16S Amplicon Sample Library for Illumina MiSEQ
4.4. Processing of 16S MiSEQ Data
4.5. Cultivation of Bacteria and Their Biomass Extraction
4.6. Antimicrobial Assay
4.7. Inhibitory Effects on Hepatitis C Virus (HCV) Infectivity
4.8. Chemical Analysis of Bioactive Extracts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ding, Y.; Xu, F.; Liu, B.; Kou, Z.; Xiao, W.; Zhu, J. Systems pharmacology-based drug discovery for marine resources: An example using sea cucumber (Holothurians). J. Ethnopharmacol. 2015, 165, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Masre, S.F.; Yip, G.W.; Sirajudeen, K.N.S.; Ghazali, F.C. Wound healing activity of total sulfated glycosaminoglycan (GAG) from Stichopus vastus and Stichopus hermanni integumental tissue in rats. Int. J. Mol. Med. Adv. Sci. 2010, 6, 49–53. [Google Scholar] [CrossRef]
- Abedin, M.Z.; Karim, A.A.; Latiff, A.A.; Gan, C.-Y.; Ghazali, F.C.; Zzaman, W.; Hossain, M.M.; Ahmed, F.; Absar, N.; Sarker, M.Z.I. Physicochemical and biochemical properties of pepsin-solubilized collagen isolated from the integument of sea cucumber (Stichopus vastus). J. Food Process. Preserv. 2014, 38, 2027–2036. [Google Scholar] [CrossRef]
- Abedin, M.; Karim, A.; Gan, C.; Ghazali, F.; Barzideh, Z.; Zzaman, W.; Zaidul, I. Identification of angiotensin I converting enzyme inhibitory and radical scavenging bioactive peptides from sea cucumber (Stichopus vastus) collagen hydrolysates through optimization. Int. Food Res. J. 2015, 22, 1074–1082. [Google Scholar]
- Conand, C.; Purcell, S.; Gamboa, R. Holothuria leucospilota. In The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2013. [Google Scholar]
- Baharara, J.; Amini, E.; Vazifedan, V. Concomitant use of sea cucumber organic extract and radiotherapy on proliferation and apoptosis of cervical (HeLa) cell line. Zahedan J. Res. Med. Sci. 2016, 18, e6442. [Google Scholar] [CrossRef]
- Baharara, J.; Amini, E.; Nikdel, N.; Salek-Abdollahi, F. The cytotoxicity of dacarbazine potentiated by sea cucumber saponin in resistant B16F10 melanoma cells through apoptosis induction. Avicenna J. Med. Biotechnol. 2016, 8, 112–119. [Google Scholar]
- Soltani, M.; Parivar, K.; Baharara, J.; Kerachian, M.A.; Asili, J. Hemolytic and cytotoxic properties of saponin purified from Holothuria leucospilota sea cucumber. Rep. Biochem. Mol. Biol. 2014, 3, 43–50. [Google Scholar]
- Soltani, M.; Parivar, K.; Baharara, J.; Kerachian, M.A.; Asili, J. Putative mechanism for apoptosis-inducing properties of crude saponin isolated from sea cucumber (Holothuria leucospilota) as an antioxidant compound. Iran. J. Basic Med. Sci. 2015, 18, 180–187. [Google Scholar]
- Pangestuti, R.; Arifin, Z. Medicinal and health benefit effects of functional sea cucumbers. J. Tradit. Complement. Med. 2018, 8, 341–351. [Google Scholar] [CrossRef]
- Farjami, B.; Nematollahi, M.A.; Moradi, Y.; Irajian, G.; Nazemi, M.; Ardebili, A.; Pournajaf, A. Antibacterial activity of the sea cucumber Holothuria leucospilota. Int. J. Mol. Clin. Microbiol. 2013, 3, 225–230. [Google Scholar]
- Adibpour, N.; Nasr, F.; Nematpour, F.; Shakouri, A.; Ameri, A. Antibacterial and antifungal activity of Holothuria leucospilota isolated from Persian Gulf and Oman Sea. Jundishapur J. Microbiol. 2014, 7, e8708. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P.; Edrada, R.; Ebel, R. Drugs from the seas–current status and microbiological implications. Appl. Microbiol. Biotechnol. 2002, 59, 125–134. [Google Scholar] [PubMed]
- Haygood, M.G.; Schmidt, E.W.; Davidson, S.K.; Faulkner, D.J. Microbial symbionts of marine invertebrates: Opportunities for microbial biotechnology. J. Mol. Microbiol. Biotechnol. 1999, 1, 33–43. [Google Scholar] [PubMed]
- Osinga, R.; Armstrong, E.; Burgess, J.G.; Hoffmann, F.; Reitner, J.; Schumann-Kindel, G. Sponge–microbe associations and their importance for sponge bioprocess engineering. Hydrobiologia 2001, 461, 55–62. [Google Scholar] [CrossRef]
- Müller, R.; Wink, J. Future potential for anti-infectives from bacteria—How to exploit biodiversity and genomic potential. Int. J. Med. Microbiol. 2014, 304, 3–13. [Google Scholar] [CrossRef]
- Stadler, M.; Dersch, P. How to Overcome the Antibiotic Crisis; Springer: Berlin, Germany, 2017. [Google Scholar]
- Pietschmann, T.; Brown, R.J. Hepatitis C virus. Trends Microbiol. 2019, 27, 379–380. [Google Scholar] [CrossRef]
- Roy, V.; King, L. Betting on hepatitis C: How financial speculation in drug development influences access to medicines. Bmj 2016, 354, i3718. [Google Scholar] [CrossRef]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association—A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef]
- Moitinho-Silva, L.; Nielsen, S.; Amir, A.; Gonzalez, A.; Ackermann, G.L.; Cerrano, C.; Astudillo-Garcia, C.; Easson, C.; Sipkema, D.; Liu, F. The sponge microbiome project. Gigascience 2017, 6, gix077. [Google Scholar] [CrossRef]
- Steinert, G.; Rohde, S.; Janussen, D.; Blaurock, C.; Schupp, P.J. Host-specific assembly of sponge-associated prokaryotes at high taxonomic ranks. Sci. Rep. 2017, 7, 2542. [Google Scholar] [CrossRef] [PubMed]
- Steinert, G.; Taylor, M.W.; Deines, P.; Simister, R.L.; de Voogd, N.J.; Hoggard, M.; Schupp, P.J. In four shallow and mesophotic tropical reef sponges from Guam the microbial community largely depends on host identity. PeerJ 2016, 4, e1936. [Google Scholar] [CrossRef] [PubMed]
- Steinert, G.; Taylor, M.W.; Schupp, P.J. Diversity of actinobacteria associated with the marine ascidian Eudistoma toealensis. Mar. Biotechnol. 2015, 17, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Tawfike, A.; Attia, E.Z.; Desoukey, S.Y.; Hajjar, D.; Makki, A.A.; Schupp, P.J.; Edrada-Ebel, R.; Abdelmohsen, U.R. New bioactive metabolites from the elicited marine sponge-derived bacterium Actinokineospora spheciospongiae sp. nov. AMB Express 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Björk, J.R.; Easson, C.; Astudillo-García, C.; Olson, J.B.; Erwin, P.M.; López-Legentil, S.; Luter, H. Diversity, structure and convergent evolution of the global sponge microbiome. Nat. Commun. 2016, 7, 11870. [Google Scholar] [CrossRef] [Green Version]
- Steinert, G.; Whitfield, S.; Taylor, M.W.; Thoms, C.; Schupp, P.J. Application of diffusion growth chambers for the cultivation of marine sponge-associated bacteria. Mar. Biotechnol. 2014, 16, 594–603. [Google Scholar] [CrossRef]
- Gao, F.; Li, F.; Tan, J.; Yan, J.; Sun, H. Bacterial community composition in the gut content and ambient sediment of sea cucumber Apostichopus japonicus revealed by 16S rRNA gene pyrosequencing. PLoS ONE 2014, 9, e100092. [Google Scholar] [CrossRef]
- Böhringer, N.; Fisch, K.M.; Schillo, D.; Bara, R.; Hertzer, C.; Grein, F.; Eisenbarth, J.-H.; Kaligis, F.; Schneider, T.; Wägele, H. Antimicrobial potential of bacteria associated with marine sea slugs from North Sulawesi, Indonesia. Front. Microbiol. 2017, 8, 1092. [Google Scholar] [CrossRef]
- Tangerina, M.M.; Correa, H.; Haltli, B.; Vilegas, W.; Kerr, R.G. Bioprospecting from cultivable bacterial communities of marine sediment and invertebrates from the underexplored Ubatuba region of Brazil. Arch. Microbiol. 2017, 199, 155–169. [Google Scholar] [CrossRef]
- Zhang, X.; Nakahara, T.; Miyazaki, M.; Nogi, Y.; Taniyama, S.; Arakawa, O.; Inoue, T.; Kudo, T. Diversity and function of aerobic culturable bacteria in the intestine of the sea cucumber Holothuria leucospilota. J. Gen. Appl. Microbiol. 2012, 58, 447–456. [Google Scholar] [CrossRef]
- Dupont, S.; Carre-Mlouka, A.; Domart-Coulon, I.; Vacelet, J.; Bourguet-Kondracki, M.-L. Exploring cultivable bacteria from the prokaryotic community associated with the carnivorous sponge Asbestopluma hypogea. FEMS Microbiol. Ecol. 2014, 88, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Zhang, X.H. Vibrio harveyi: A significant pathogen of marine vertebrates and invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, Z.; Li, Z.; Wang, L.; Li, H.; Wu, F.; Jin, L.; Li, X.; Li, S.; Xu, Y. Effect of bacteriophages onVibrio alginolyticusInfection in the sea cucumber, Apostichopus japonicus (Selenka). J. World Aquac. Soc. 2015, 46, 149–158. [Google Scholar] [CrossRef]
- Huang, J.; Zeng, B.; Liu, D.; Wu, R.; Zhang, J.; Liao, B.; He, H.; Bian, F. Classification and structural insight into vibriolysin-like proteases of Vibrio pathogenicity. Microb. Pathog. 2018, 117, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, Y.; Sheng, Y.; Su, P.; Qiu, Y.; Ke, C.; Feng, D. Antifouling activity towards mussel by small-molecule compounds from a strain of Vibrio alginolyticus bacterium associated with sea anemone Haliplanella sp. J. Microbiol. Biotechnol. 2017, 27, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Lateef, A.; Adelere, I.A.; Gueguim-Kana, E.B. The biology and potential biotechnological applications of Bacillus safensis. Biologia 2015, 70, 411–419. [Google Scholar] [CrossRef]
- Gesheva, V.; Vasileva-Tonkova, E. Production of enzymes and antimicrobial compounds by halophilic Antarctic Nocardioides sp. grown on different carbon sources. World J. Microbiol. Biotechnol. 2012, 28, 2069–2076. [Google Scholar] [CrossRef]
- Pan, H.Q.; Yu, S.Y.; Song, C.F.; Wang, N.; Hua, H.M.; Hu, J.C.; Wang, S.J. Identification and characterization of the antifungal substances of a novel Streptomyces cavourensis NA4. J. Microbiol. Biotechnol. 2015, 25, 353–357. [Google Scholar] [CrossRef]
- Begum, I.F.; Mohankumar, R.; Jeevan, M.; Ramani, K. GC–MS analysis of bio-active molecules derived from Paracoccus pantotrophus FMR19 and the antimicrobial activity against bacterial pathogens and MDROs. Indian J. Microbiol. 2016, 56, 426–432. [Google Scholar] [CrossRef]
- Zhang, F.; Ye, Q.; Chen, Q.; Yang, K.; Zhang, D.; Chen, Z.; Lu, S.; Shao, X.; Fan, Y.; Yao, L. Algicidal activity of novel marine bacterium Paracoccus sp. strain Y42 against a harmful algal-bloom-causing dinoflagellate, Prorocentrum donghaiense. Appl. Environ. Microbiol. 2018, 84, e01015-18. [Google Scholar] [CrossRef]
- Lukman, A.L.; Nordin, N.F.H.; Kamarudin, K.R. Microbial population in the coelomic fluid of Stichopus chloronotus and Holothuria (Mertensiothuria) leucospilota collected from Malaysian waters. Sains Malays. 2014, 43, 1013–1021. [Google Scholar]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Matloub, A.A.; Gomaa, E.Z.; Hassan, A.A.; Elbatanony, M.M.; El-Senousy, W.M. Comparative chemical and bioactivity studies of intra-and extracellular metabolites of endophytic bacteria, Bacillus subtilis NCIB 3610. Int. J. Pept. Res. Ther. 2019. [Google Scholar] [CrossRef]
- Covington, B.C.; McLean, J.A.; Bachmann, B.O. Comparative mass spectrometry-based metabolomics strategies for the investigation of microbial secondary metabolites. Nat. Prod. Rep. 2017, 34, 6–24. [Google Scholar] [CrossRef]
- Yoshioka, T.; Watanabe, A.; Kojima, I.; Shimauchi, Y.; Okabe, M.; Fukagawa, Y.; Ishikura, T. Structures of OA-6129D and E, new carbapenam antibiotics. J. Antibiot. 1984, 37, 211–217. [Google Scholar] [CrossRef]
- Paulo, B.S.; Sigrist, R.; Angolini, C.F.; de Oliveira, L.G. New cyclodepsipeptide derivatives revealed by genome mining and molecular networking. ChemistrySelect 2019, 4, 7785–7790. [Google Scholar] [CrossRef]
- Callow, R.; Taylor, D. 429. The cardio-active glycosides of Strophanthus sarmentosus P. DC.“sarmentoside B” and its relation to an original sarmentobioside. J. Chem. Soc. 1952. [Google Scholar] [CrossRef]
- Martin, J.; da S Sousa, T.; Crespo, G.; Palomo, S.; Gonzalez, I.; Tormo, J.R.; de la Cruz, M.; Anderson, M.; Hill, R.T.; Vicente, F.; et al. Kocurin, the true structure of PM181104, an anti-methicillin-resistant Staphylococcus aureus (MRSA) thiazolyl peptide from the marine-derived bacterium Kocuria palustris. Mar. Drugs 2013, 11, 387–398. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Yang, S.-Z.; Mu, B.-Z. Production and characterization of a C15-surfactin-O-methyl ester by a lipopeptide producing strain Bacillus subtilis HSO121. Process. Biochem. 2009, 44, 1144–1151. [Google Scholar] [CrossRef]
- Kalyon, B.; Helaly, S.E.; Scholz, R.; Nachtigall, J.; Vater, J.; Borriss, R.; Süssmuth, R.D. Plantazolicin A and B: Structure elucidation of ribosomally synthesized thiazole/oxazole peptides from Bacillus amyloliquefaciens FZB42. Org. Lett. 2011, 13, 2996–2999. [Google Scholar] [CrossRef]
- Wright, G.D. Opportunities for natural products in 21st century antibiotic discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H. Renaissance in antibacterial discovery from actinomycetes. Curr. Opin. Pharmacol. 2008, 8, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Schinke, C.; Martins, T.; Queiroz, S.C.N.; Melo, I.S.; Reyes, F.G.R. Antibacterial compounds from marine bacteria, 2010–2015. J. Nat. Prod. 2017, 80, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, Y. Pharmacological potential of sea cucumbers. Int. J. Mol. Sci. 2018, 19, 1342. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Conand, C.; Uthicke, S.; Byrne, M. Ecological roles of exploited sea cucumbers. In Oceanography and Marine Biology: An Annual Review V54; CRC Press: Boca Raton, FL, USA, 2016; pp. 367–386. [Google Scholar]
- Pereira, D.M.; Andrade, P.B.; Pires, R.A.; Reis, R.L. Chemical Ecology of Echinoderms: Impact of Environment and Diet in Metabolomic Profile; Nova Science Publishers: Hauppauge, NY, USA, 2014. [Google Scholar]
- Seipke, R.F.; Kaltenpoth, M.; Hutchings, M.I. Streptomyces as symbionts: An emerging and widespread theme? FEMS Microbiol. Rev. 2012, 36, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Kamarudin, K.R.; Ngah, N.; Hamid, T.H.T.A.; Susanti, D. Isolation of a PIgment-producing strain of Staphylococcus kloosii from the respiratory tree of Holothuria (Mertensiothuria) leucospilota (Brandt 1835) from Malaysian waters. Trop. Life Sci. Res. 2013, 24, 85–100. [Google Scholar] [PubMed]
- Brockmann, H.; Schmidt-Kastner, G. Valinomycin I, XXVII. Mitteil. über antibiotica aus actinomyceten. Chem. Ber. 1955, 88, 57–61. [Google Scholar] [CrossRef]
- Pimentel-Elardo, S.M.; Kozytska, S.; Bugni, T.S.; Ireland, C.M.; Moll, H.; Hentschel, U. Anti-parasitic compounds from Streptomyces sp. strains isolated from Mediterranean sponges. Mar. Drugs 2010, 8, 373–380. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Jan, J.-T.; Ma, S.-H.; Kuo, C.-J.; Juan, H.-F.; Cheng, Y.-S.E.; Hsu, H.-H.; Huang, H.-C.; Wu, D.; Brik, A. Small molecules targeting severe acute respiratory syndrome human coronavirus. Proc. Natl. Acad. Sci. USA 2004, 101, 10012–10017. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Q. Deciphering the biosynthetic codes for the potent anti-SARS-CoV cyclodepsipeptide valinomycin in Streptomyces tsusimaensis ATCC 15141. ChemBioChem 2006, 7, 471–477. [Google Scholar] [CrossRef]
- Sabdono, A.; Sawonua, P.H.; Kartika, A.G.D.; Amelia, J.M.; Radjasa, O.K. Coral diseases in Panjang Island, Java Sea: Diversity of anti–pathogenic bacterial coral symbionts. Procedia Chem. 2015, 14, 15–21. [Google Scholar] [CrossRef]
- Scheubert, K.; Hufsky, F.; Petras, D.; Wang, M.; Nothias, L.-F.; Duehrkop, K.; Bandeira, N.; Dorrestein, P.C.; Boecker, S. Significance estimation for large scale metabolomics annotations by spectral matching. Nat. Commun. 2017, 8, 1494. [Google Scholar] [CrossRef] [PubMed]
- Sottorff, I.; Wiese, J.; Lipfert, M.; Preußke, N.; Sönnichsen, F.D.; Imhoff, J.F. Different secondary metabolite profiles of phylogenetically almost identical Streptomyces griseus strains originating from geographically remote locations. Microorganisms 2019, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Gontang, E.A.; Fenical, W.; Jensen, P.R. Phylogenetic diversity of gram-positive bacteria cultured from marine sediments. Appl. Environ. Microbiol. 2007, 73, 3272–3282. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.B.; Garland, J.L.; Bolster, C.H.; Mills, A.L. Impact of dilution on microbial community structure and functional potential: Comparison of numerical simulations and batch culture experiments. Appl. Environ. Microbiol. 2001, 67, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Dong, H.; Zhang, G.; Yu, B.; Chapman, L.R.; Fields, M.W. Microbial diversity in water and sediment of Lake Chaka, an athalassohaline lake in northwestern China. Appl. Environ. Microbiol. 2006, 72, 3832–3845. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Tindall, B.J.; Rosselló-Móra, R.; Busse, H.-J.; Ludwig, W.; Kämpfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60, 249–266. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Okanya, P.W.; Mohr, K.I.; Gerth, K.; Jansen, R.; Müller, R. Marinoquinolines A−F, pyrroloquinolines from Ohtaekwangia kribbensis (Bacteroidetes). J. Nat. Prod. 2011, 74, 603–608. [Google Scholar] [CrossRef]
- Wang, M.; Jarmusch, A.K.; Vargas, F.; Aksenov, A.A.; Gauglitz, J.M.; Weldon, K.; Petras, D.; Silva, R.D.; Quinn, R.; Melnik, A.V.; et al. MASST: A web-based basic mass spectrometry search tool for molecules to search public data. BioRxiv 2019. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Holothuria leucospilota (HL) | Stichopus vastus (SV) | TOTAL | ||
|---|---|---|---|---|---|
| Internal Part | External Part | Internal Part | External Part | ||
| Actinobacteria | 23 | 19 | 3 | 18 | 63 |
| Firmicutes | 5 | 8 | 6 | 14 | 33 |
| Proteobacteria | 11 | 10 | 1 | 8 | 30 |
| Bacteroidetes | - | - | - | 1 | 1 |
| TOTAL | 39 | 37 | 10 | 41 | 127 |
| No. | Next Related Type Strain | Sample Accession Number | Type Strain Accession Number | Similarity to Type Strain (%) | Sequence Length (bp) | Sample Name | Antimicrobial Activity | |
|---|---|---|---|---|---|---|---|---|
| Antibacteria | Antifungal | |||||||
| Phylum Actinobacteria | ||||||||
| 1. | Brevibacterium luteolum | MK696423 | NR_114872.1 | 99.63 | 1076 | SV 4 (ext) | - | - |
| 2. | Cellulosimicrobium funkei | MK696437 | NR_042937.1 | 99.78 | 915 | HL 61 (ext) | - | - |
| 3. | Corynebacterium pilbarense | MK696498 | NR_116953.1 | 98.74 | 829 | HL 119 (ext) | - | - |
| 4. | Dermacoccus nishinomiyaensis | MK696488 | NR_044872.1 | 99.72 | 1063 | HL 57 (ext) | Bs (B), Sa (C) | Rg (A), Mh (A) |
| 5. | Dermacoccus profundi | MK696484 | NR_043262.1 | 99.89 | 1076 | HL 11 (int) | - | - |
| 6. | Dermacoccus profundi | MK696494 | NR_043262.1 | 99.72 | 916 | SV 127 (ext) | - | - |
| 7. | Dietzia maris | MK696467 | NR_118596.1 | 98.84 | 1126 | SV 164b (ext) | Sa (A) | - |
| 8. | Glutamicibacter sp.* (G. nicotianae) | MK696438 | NR_026190.1 | 96.38 | 1056 | HL 108 (ext) | Bs (B), Sa (A) | Rg (A) |
| 9. | Isoptericola chiayiensis | MK696432 | NR_116696.1 | 98.88 | 894 | HL 44 (ext) | Bs (A) | - |
| 10. | Janibacter alkaliphilus | MK696433 | NR_109453.1 | 98.92 | 1018 | SV 51 (ext) | Bs (A) | - |
| 11. | Janibacter anophelis | MK696442 | NR_043218.1 | 99.15 | 1062 | HL 24 (int) | - | - |
| 12. | Janibacter melonis | MK696486 | NR_025805.1 | 99.79 | 964 | HL 40 (int) | Bs (A) | - |
| 13. | Kocuria flava | MK696544 | NR_044308.1 | 99.21 | 892 | HL 55 (int) | Bs(E), Ec (A), Sa (D) | - |
| 14. | Kocuria palustris | MK696435 | NR_026451.1 | 99.90 | 1045 | HL 6 (ext) | Bs (C), Sa (C) | Rg (A) |
| 15. | Kocuria palustris | MK696424 | NR_026451.1 | 100.00 | 956 | HL 7 (ext) | Bs (B) | - |
| 16. | Kocuria palustris | MK696425 | NR_026451.1 | 100.00 | 922 | HL 8 (ext) | Bs (A) | - |
| 17. | Kocuria palustris | MK696524 | NR_026451.1 | 99.81 | 879 | HL 60 (ext) | - | - |
| 18. | Kocuria palustris | MK696426 | NR_026451.1 | 100.00 | 1021 | HL 12 (int) | Bs (B) | - |
| 19. | Kocuria palustris | MK696522 | NR_026451.1 | 99.71 | 917 | HL 42 (int) | - | - |
| 20. | Kocuria palustris | MK696441 | NR_026451.1 | 98.76 | 913 | SV 14 (ext) | - | - |
| 21. | Kytococcus sedentarius | MK696431 | NR_074714.2 | 99.88 | 1041 | HL 30 (int) | Bs (A) | - |
| 22. | Kytococcus sedentarius | MK696446 | NR_074714.2 | 99.72 | 838 | HL 43 (int) | - | - |
| 23. | Kytococcus sedentarius | MK696483 | NR_074714.2 | 99.72 | 980 | SV 2 (ext) | Bs (B) | - |
| 24. | Micrococcus aloeverae | MK696444 | NR_134088.1 | 99.78 | 1041 | HL 33 (ext) | - | - |
| 25. | Micrococcus aloeverae | MK696430 | NR_134088.1 | 99.79 | 937 | HL 29 (int) | Bs (A) | - |
| 26. | Micrococcus aloeverae | MK696436 | NR_134088.1 | 99.52 | 918 | SV 5 (ext) | Bs (A) | Rg (A) |
| 27. | Micrococcus aloeverae | MK696523 | NR_134088.1 | 99.36 | 908 | SV 52 (ext) | - | - |
| 28. | Micrococcus endophyticus | MK696473 | NR_044365.1 | 98.62 | 1018 | HL 261 (int) | - | - |
| 29. | Micrococcus flavus | MK696517 | NR_043881.1 | 99.20 | 1005 | HL 237 (int) | Bs (B), Sa (B), Rg (B) | - |
| 30. | Micrococcus terreus | MK696528 | NR_116649.1 | 99.44 | 1081 | SV 137 (ext) | - | - |
| 31. | Nocardioides sp.* (N. exalbidus) | MK696451 | NR_041526.1 | 97.96 | 1036 | HL 111 (ext) | Bs (A), Sa (H) | - |
| 32. | Ornithinimicrobium kibberense | MK696459 | NR_043056.1 | 99.59 | 988 | SV 135 (ext) | - | - |
| 33. | New Genus of family Propionibacteriaceae * (Pseudopropionibacterium profundi) | MK696480 | NR_159102.1 | 93.29 | 1047 | SV 17 (ext) | - | - |
| 34. | Rothia kristinae | MK696477 | NR_026199.1 | 99.29 | 989 | HL 37 (ext) | - | - |
| 35. | Serinicoccus sp.* (S. profundi) | MK696482 | NR_116387.1 | 97.91 | 719 | SV 16 (ext) | - | - |
| 36. | Streptomyces cavourensis | MK696479 | NR_043851.1 | 100.00 | 1034 | SV 21 (int) | Bs (H), Sa (E) | Mh (G) |
| Phylum Firmicutes | ||||||||
| 1. | Bacillus aryabhattai | MK696496 | NR_115953.1 | 99.91 | 1134 | HL 270 (int) | - | - |
| 2. | Bacillus cereus | MK696514 | NR_157734.1 | 99.91 | 1132 | HL 229 (int) | - | - |
| 3. | Bacillus idriensis | MK696468 | NR_043268.1 | 99.47 | 948 | HL 251 (int) | - | - |
| 4. | Bacillus safensis | MK696463 | NR_041794.1 | 100.00 | 927 | SV 147 (ext) | Bs (B), Sa (H) | - |
| 5. | Bacillus safensis | MK696525 | NR_113945.1 | 99.91 | 1126 | HL 63 (ext) | Sa (H) | - |
| 6. | New genus of family Bacillaceae (Bacillus sonorensis) * | MK696542 | NR_113993.1 | 92.88 | 1081 | HL 79 (ext) | Sa (A) | - |
| 7. | Staphylococcus arlettae | MK696500 | NR_024664.1 | 99.65 | 1149 | SV 133 (ext) | - | - |
| 8. | Staphylococcus cohnii | MK696452 | NR_036902.1 | 99.80 | 990 | HL 113 (ext) | - | - |
| 9. | Staphylococcus cohnii subsp. urealyticus | MK696447 | NR_037046.1 | 99.31 | 1121 | HL 67 (ext) | Sa (H) | - |
| 10. | Staphylococcus cohnii subsp. urealyticus | MK696422 | NR_037046.1 | 100.00 | 1116 | SV 1 (ext) | Bs (B) | - |
| 11. | Staphylococcus cohnii subsp. urealyticus | MK696458 | NR_037046.1 | 99.73 | 953 | SV 131 (ext) | - | - |
| 12. | Staphylococcus cohnii subsp. urealyticus | MK696440 | NR_037046.1 | 99.80 | 1098 | SV 144 (ext) | Sa (A) | - |
| 13. | Staphylococcus edaphicus | MK696526 | NR_156818.1 | 99.73 | 1108 | HL 75 (ext) | Bs (C), Sa (B) | Rg (B) |
| 14. | Staphylococcus haemolyticus | MK696532 | NR_113345.1 | 99.33 | 1044 | SV 183 (ext) | - | - |
| 15. | Staphylococcus pasteuri | MK696531 | NR_114435.1 | 99.91 | 1117 | SV 173 (ext) | - | - |
| 16. | Staphylococcus warneri | MK696543 | NR_025922.1 | 99.91 | 1084 | HL 100 (ext) | Sa (A) | - |
| Phylum Proteobacteria | ||||||||
| 1. | Acinetobacter schindleri | MK696475 | NR_025412.1 | 98.95 | 1047 | HL 265 (int) | - | - |
| 2. | Epibacterium mobile | MK696445 | NR_114024.1 | 99.64 | 1112 | HL 38 (ext) | Bs (B) | - |
| 3. | Erythrobacter vulgaris | MK696434 | NR_043136.1 | 99.18 | 980 | SV 54 (int) | Ec (A) | - |
| 4. | Erythrobacter vulgaris | MK696478 | NR_043136.1 | 99.36 | 937 | HL 45 (ext) | Bs (A) | - |
| 5. | Pantoea septica | MK696487 | NR_116752.1 | 99.14 | 1080 | SV 138 (ext) | Bs (A) | - |
| 6. | New genus of family Rhodobacteraceae (Paracoccus beibuensis) | - | NR_116400.1 | 93.08 | 1011 | SV 155 (ext) | Sa (E) | - |
| 7. | Paracoccus sp.* (P. koreensis) | MK696429 | NR_114060.1 | 97.33 | 940 | HL 28 (int) | Bs (A) | Mh (A) |
| 8. | Paracoccus marinus | MK696491 | NR_113921.1 | 99.03 | 928 | HL 256 (int) | Bs (A), Sa (A) | - |
| 9. | Paracoccus sulfuroxidans | MK696428 | NR_043887.1 | 98.25 | 861 | HL 27 (int) | - | - |
| 10. | Pseudomonas stutzeri | MK696497 | NR_041715.1 | 99.21 | 1079 | HL 26 (int) | Bs (B) | - |
| 11. | Psychrobacter celer | MK696489 | NR_043225.1 | 99.27 | 1100 | HL 58 (ext) | - | - |
| 12. | Psychrobacter marincola | MK696539 | NR_025458.1 | 99.40 | 1165 | HL 72 (ext) | Sa (A) | - |
| 13. | Vibrio alginolyticus | MK696427 | NR_118258.1 | 99.52 | 1039 | HL 22 (ext) | Bs (G), Sa (E), Ms (A) | Rg (B), Mh (B) |
| 14. | Vibrio sp.* (V. harveyi) | MK696456 | NR_043165.1 | 96.23 | 1074 | HL 125 (ext) | Bs (A), Sa (A) | - |
| 15. | Vibrio harveyi | MK696454 | NR_113784.1 | 99.80 | 996 | HL 121 (ext) | Sa (A) | Rg (A) |
| 16. | Vibrio owensii | MK696449 | NR_117424.1 | 99.34 | 907 | HL 107 (ext) | - | - |
| 17. | Vibrio owensii | MK696499 | NR_117424.1 | 99.44 | 1082 | HL 122 (ext) | Sa (A) | Rg (A) |
| Strain | Precursor Ions (m/z) | Finding Match Compounds Based on MS1 in Databases (M ± 0.005) | Finding Match Compounds Based on MS2 in Databases (incl. Analog) * | Annotation |
|---|---|---|---|---|
| Streptomyces cavourensis SV 21 | 458.181 [M + H]+ (M = 457.173) | F | LV | Partly identified with low match value |
| 490.207 [M + H]+ (M = 489.200) | F | LV | Partly identified with low match value | |
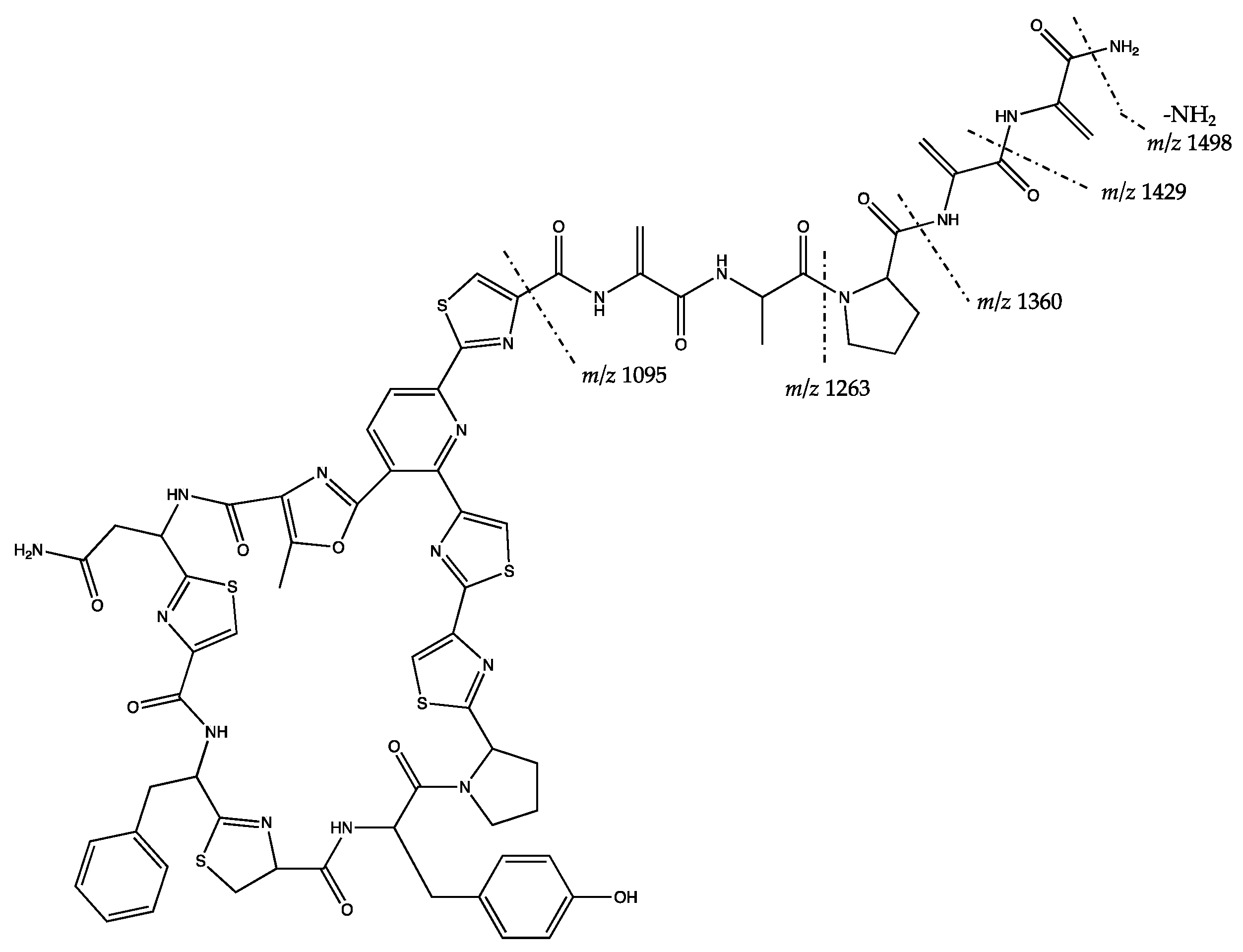
| 1128.665 [M + NH4]+ (M = 1110.630) | F | F | Putative Valinomycin * | |
| 1142.678 [M + NH4]+ (M = 1124.644) | NF | F | Partly identified as valinomycin derivate * | |
| 663.454 [M + H]+ (M = 662.447) | F | LV | Partly identified with low match value | |
| Kocuria flava HL 55 | 1140.219 [M + H]+ (M = 1139.211) | NF | NF | unidentified |
| 1515.373 [M + H]+ (M = 1514.366) | F | F | Putative kocurin * | |
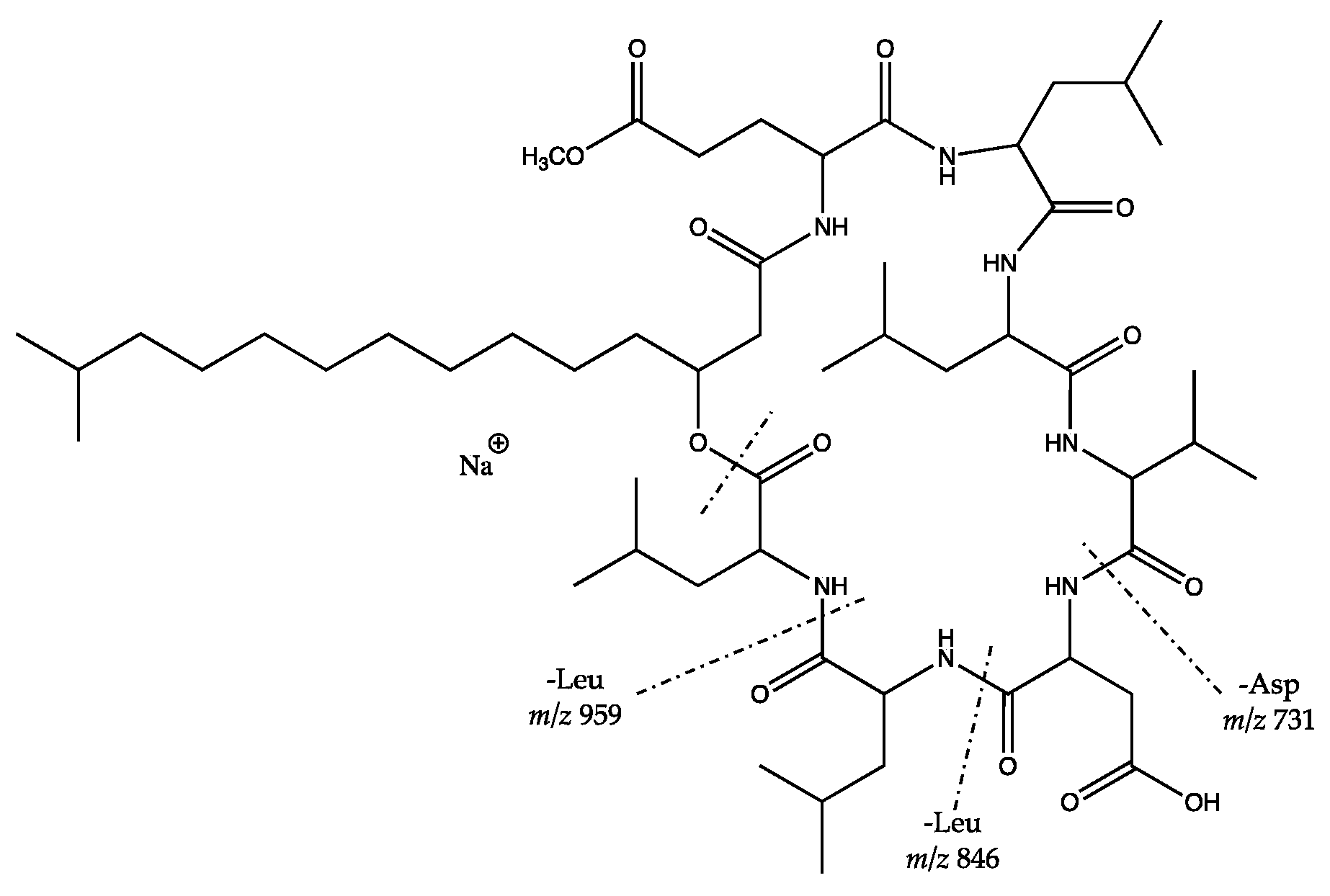
| Bacillus safensis HL 63 and Staphylococcus cohnii subsp. urealyticus HL 67 | 1070.643 [M + H]+ (M = 1069.636) | NF | F | Putative surfactins |
| 1102.616 [M + H]+ (M = 1101.609) | F | F | Putative surfactins | |
| 1076.629 [M+Na]+ (M = 1053.640) | NF | F | Putative surfactins | |
| 1068.661 [M + H]+ (M = 1067.654) | F | F | Putative surfactins | |
| 1022.674 [M + H]+ (M = 1021.667) | F | F | Putative surfactins | |
| 1058.671 [M + Na]+ (M = 1035.684) | F | F | Putative surfactins * | |
| 1072.686 [M + Na]+ (M = 1049.698) | F | F | Putative surfactins * | |
| 1096.692 [M + H]+ (M = 1095.685) | F | F | Putative surfactins | |
| 1086.702 [M + Na]+ (M = 1063.714) | F | F | Putative surfactins | |
| 875.534 [M + Na]+ (M = 852.545) | NF | NF | unidentified | |
| 1100.717 [M + Na]+ (M = 1077.723) | F | F | Putative surfactins | |
| Staphylococcus edaphicus HL 75 | 347.212 [M + H]+ (M = 346.205) | F | LV | Partly identified with low match value |
| 395.213 [M + H]+ (M = 394.206) | F | LV | Partly identified with low match value | |
| Bacillus safensis SV 147, SV 155 (putatively new genus of Rhodobacteraceae), and Nocardioides sp. HL 111 | 1336.478 [M + H]+ (M = 1335.471) | F | F | Putative plantazolicin A * |
| 1044.657 [M + Na]+ (M = 1021.668) | F | F | Putative surfactins | |
| 1058.671 [M + Na]+ (M = 1035.683) | F | F | Putative surfactins | |
| 1050.705 [M + H]+ (M = 1049.698) | F | F | Putative surfactins | |
| 1086.703 [M + Na]+ (M = 1063.713) | F | F | Putative surfactins |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wibowo, J.T.; Kellermann, M.Y.; Versluis, D.; Putra, M.Y.; Murniasih, T.; Mohr, K.I.; Wink, J.; Engelmann, M.; Praditya, D.F.; Steinmann, E.; et al. Biotechnological Potential of Bacteria Isolated from the Sea Cucumber Holothuria leucospilota and Stichopus vastus from Lampung, Indonesia. Mar. Drugs 2019, 17, 635. https://doi.org/10.3390/md17110635
Wibowo JT, Kellermann MY, Versluis D, Putra MY, Murniasih T, Mohr KI, Wink J, Engelmann M, Praditya DF, Steinmann E, et al. Biotechnological Potential of Bacteria Isolated from the Sea Cucumber Holothuria leucospilota and Stichopus vastus from Lampung, Indonesia. Marine Drugs. 2019; 17(11):635. https://doi.org/10.3390/md17110635
Chicago/Turabian StyleWibowo, Joko T., Matthias Y. Kellermann, Dennis Versluis, Masteria Y. Putra, Tutik Murniasih, Kathrin I. Mohr, Joachim Wink, Michael Engelmann, Dimas F. Praditya, Eike Steinmann, and et al. 2019. "Biotechnological Potential of Bacteria Isolated from the Sea Cucumber Holothuria leucospilota and Stichopus vastus from Lampung, Indonesia" Marine Drugs 17, no. 11: 635. https://doi.org/10.3390/md17110635
APA StyleWibowo, J. T., Kellermann, M. Y., Versluis, D., Putra, M. Y., Murniasih, T., Mohr, K. I., Wink, J., Engelmann, M., Praditya, D. F., Steinmann, E., & Schupp, P. J. (2019). Biotechnological Potential of Bacteria Isolated from the Sea Cucumber Holothuria leucospilota and Stichopus vastus from Lampung, Indonesia. Marine Drugs, 17(11), 635. https://doi.org/10.3390/md17110635