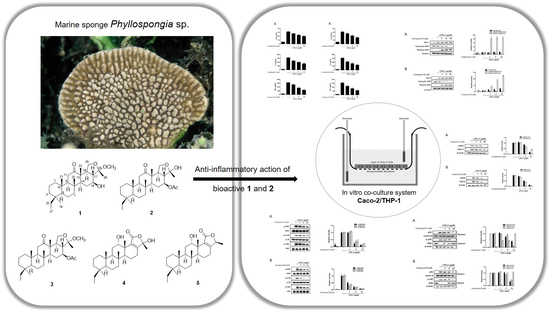
Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine

 , and
, and 
Abstract
:
1. Introduction
2. Results and Discussion
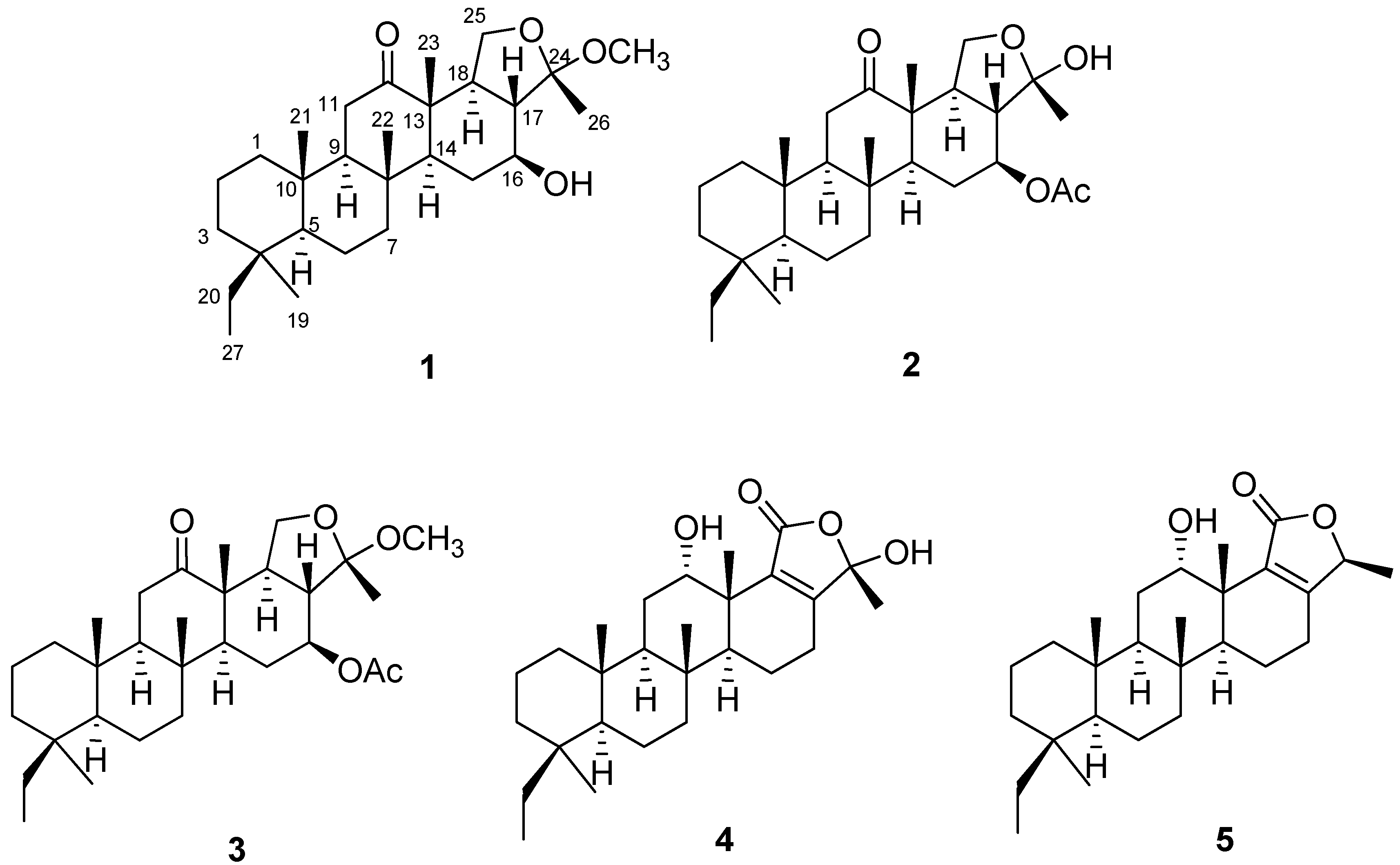
2.1. Isolation of Anti-Inflammatory Scalarane Sesterterpenes from Sponge Genus Phyllospongia Using Bioactivity-Guided Fractionation
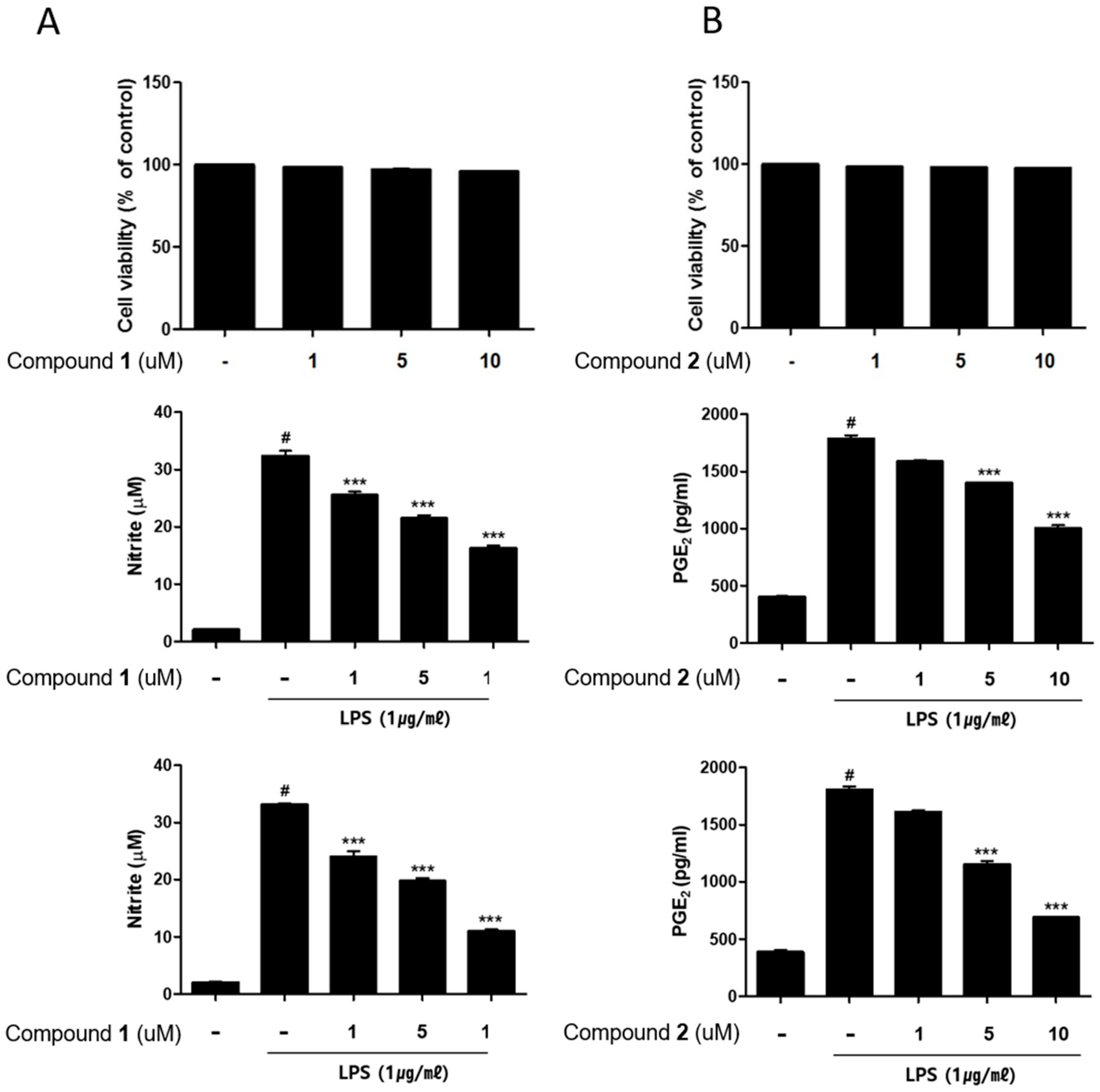
2.2. The Anti-Inflammatory Effects of 1–5 in RAW264.7 and THP-1 Macrophages
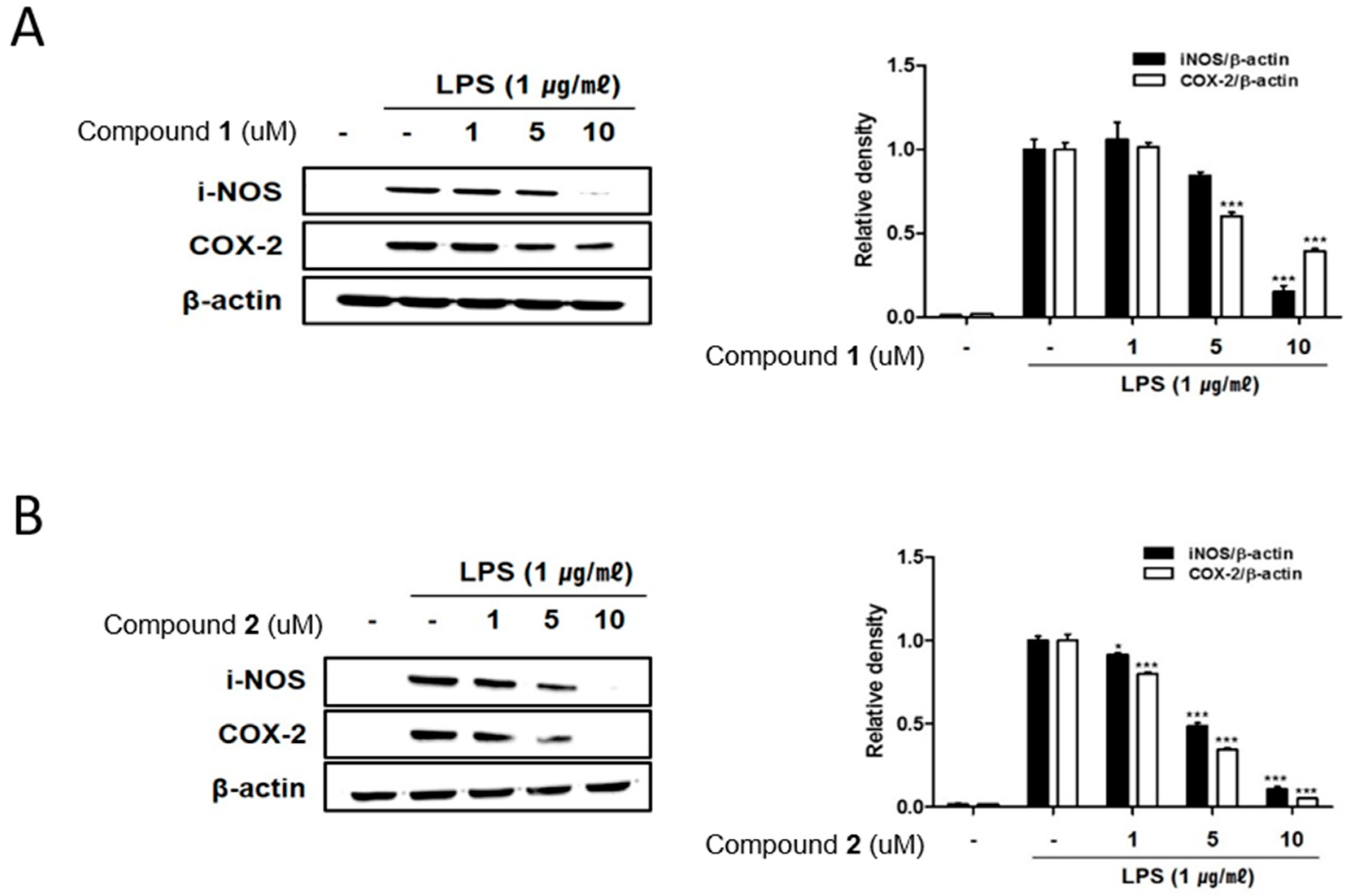
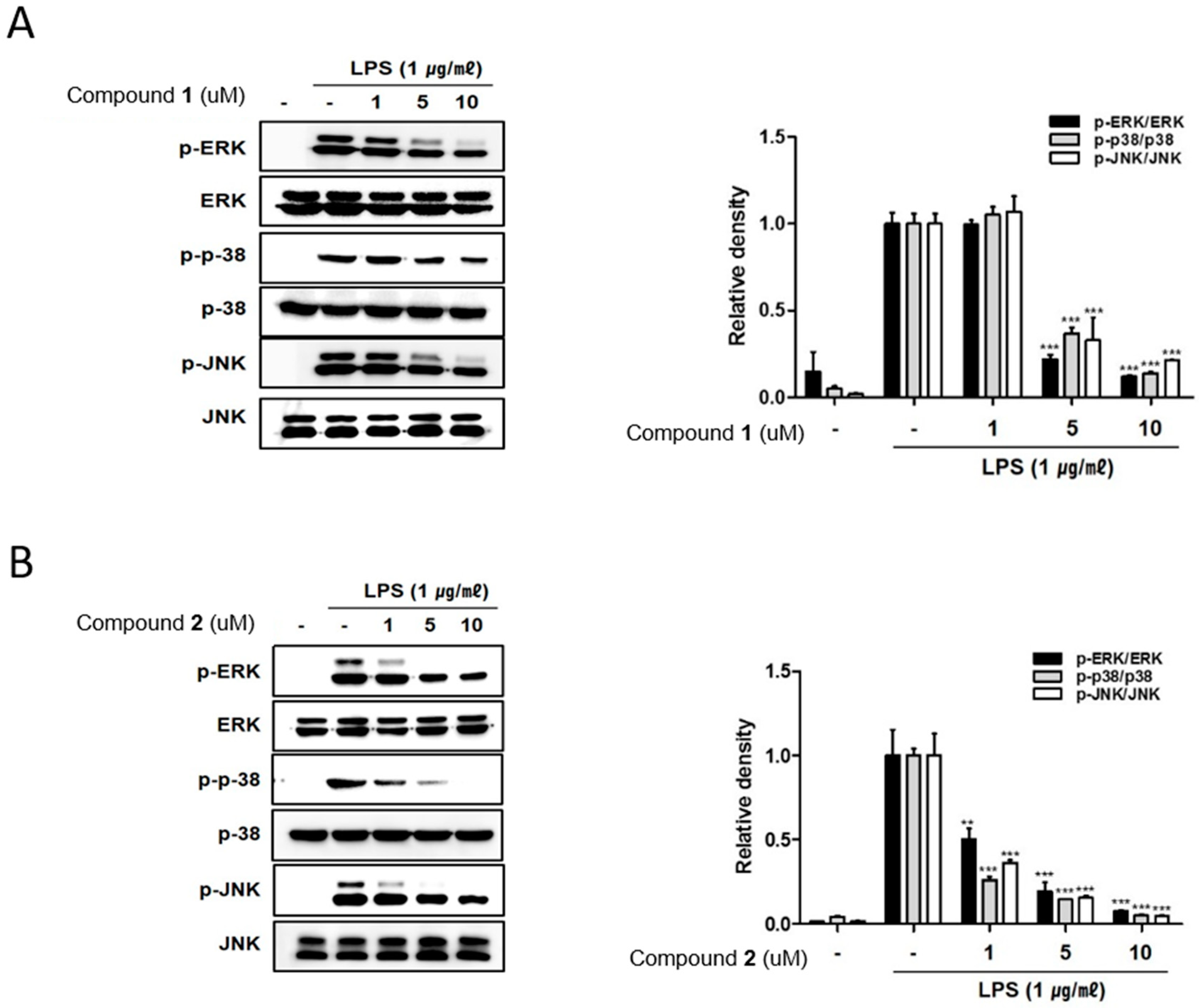
2.3. The Effects of Deacetylphylloketal and Phyllohemikal A on the Pro-Inflammatory Protein Expression and MAPK Phosphorylation in Caco-2 Co-Cultured THP-1 Macrophages
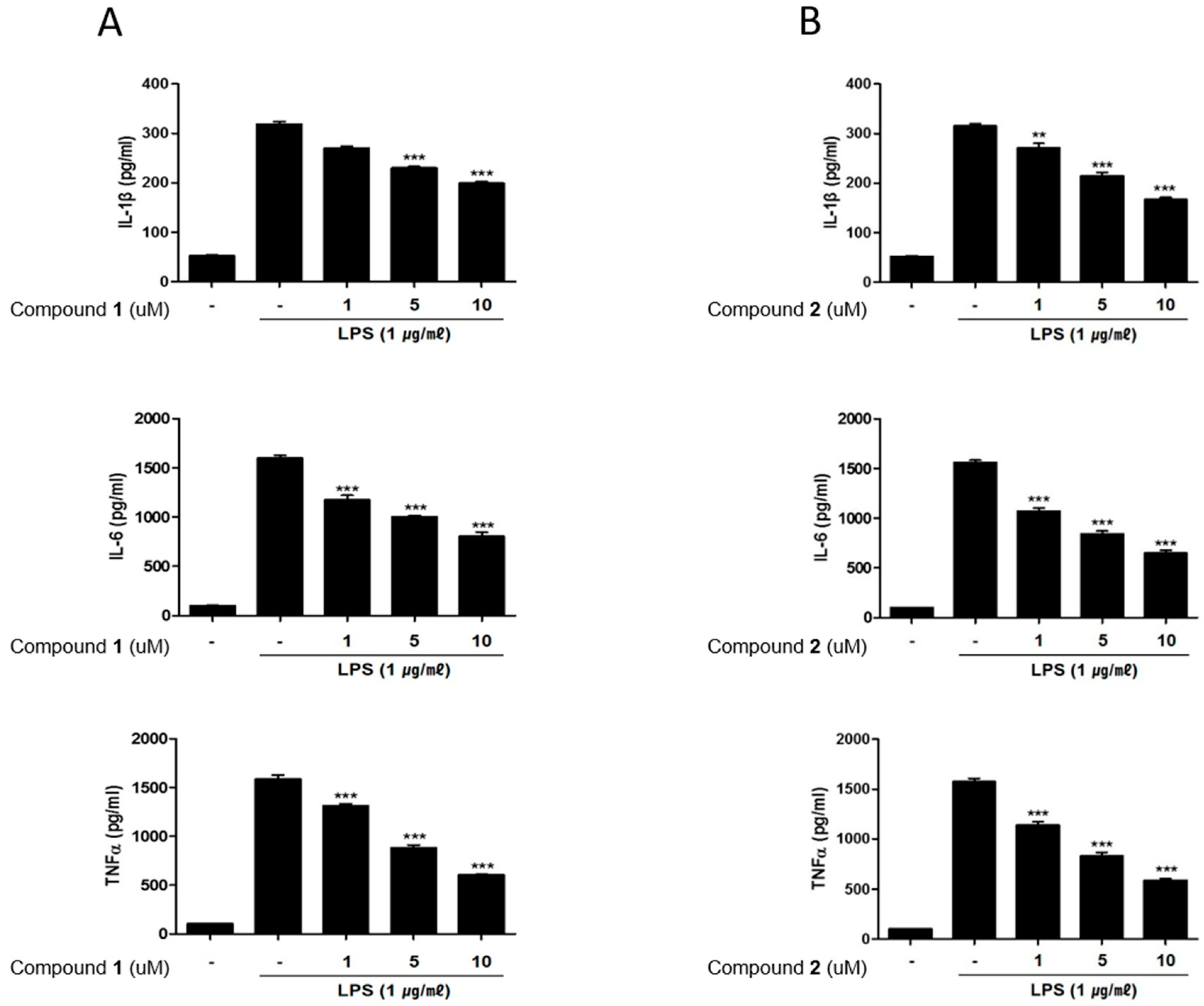
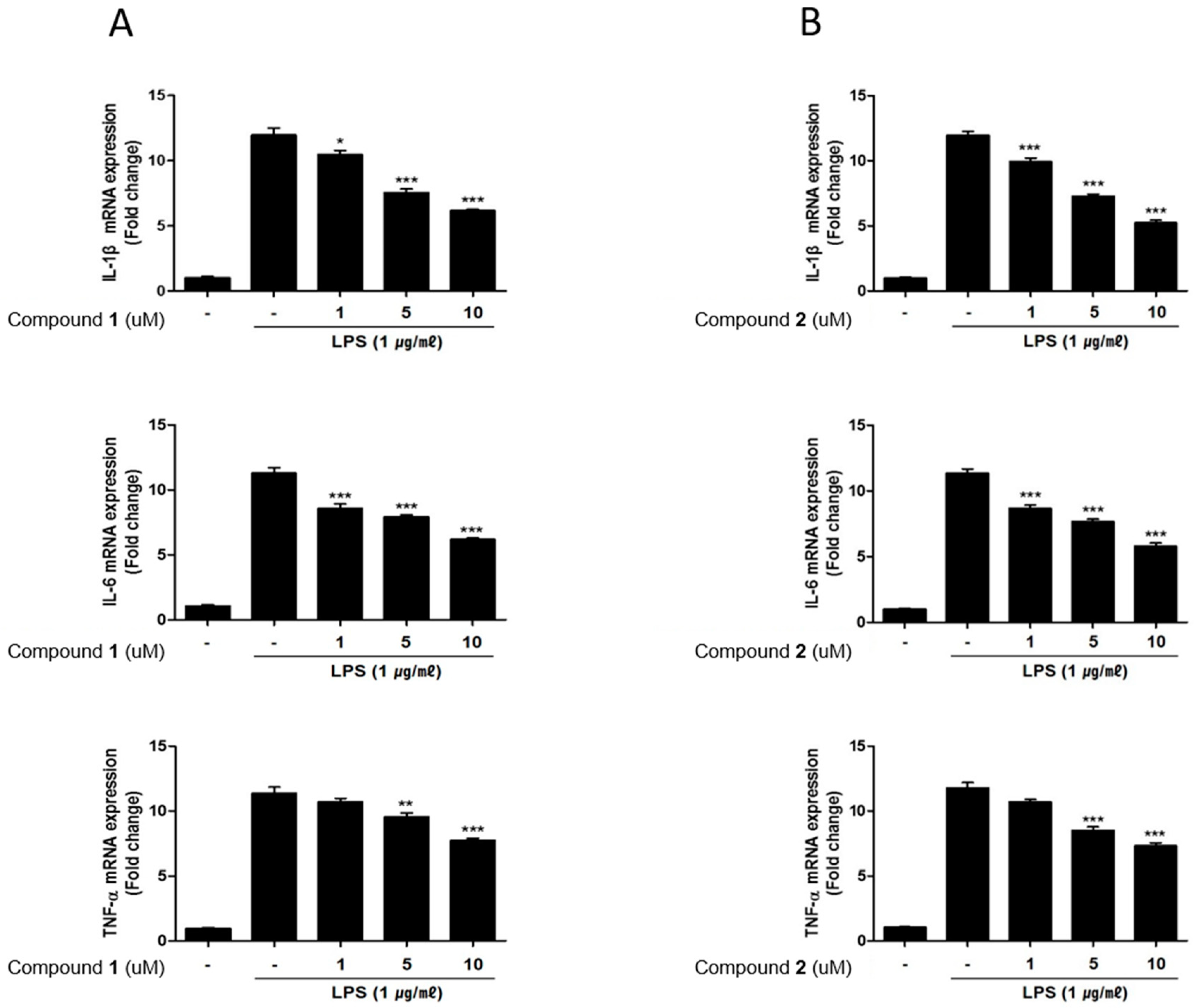
2.4. The Effects of Deacetylphylloketal and Phyllohemikal A on the Production and mRNA Expression of Proinflammatory Cytokines in Caco-2 Co-Cultured THP-1 Macrophages
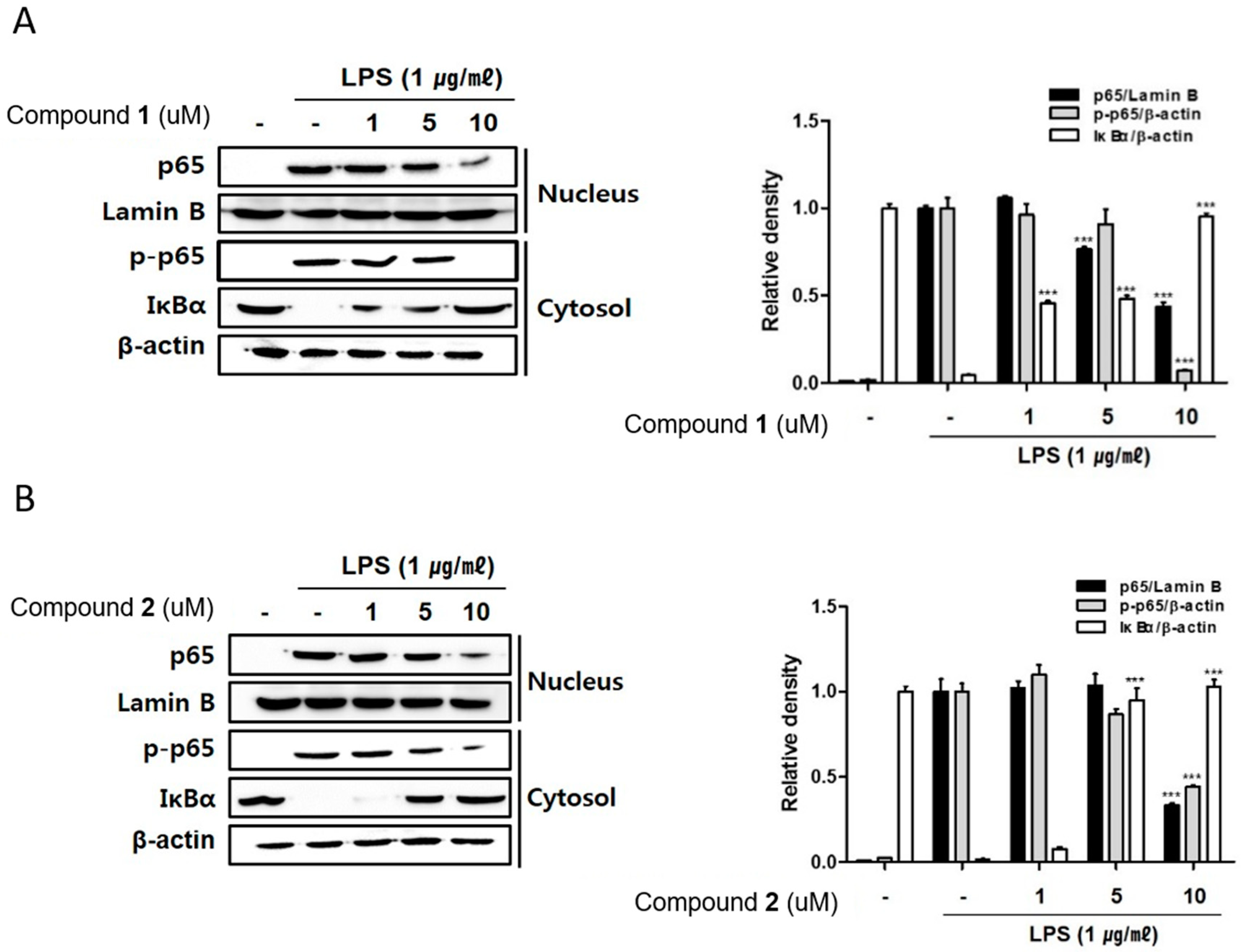
2.5. The Effects of Deacetylphylloketal and Phyllohemikal A on the Nucleus Translocation of NF-κB and Its Inhibitor, IκB-α
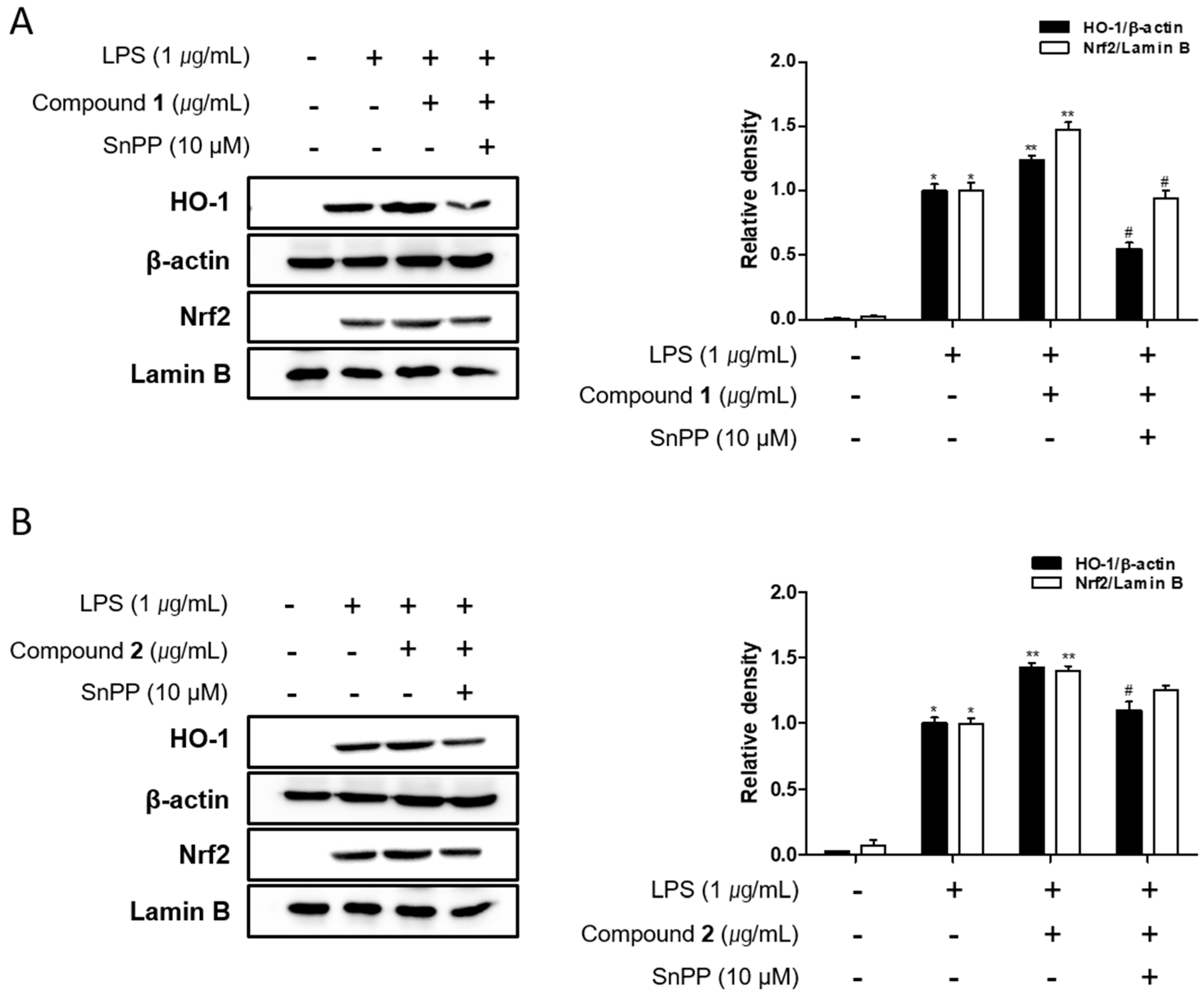
2.6. The Effects of Deacetylphylloketal and Phyllohemikal A on the Expression of HO-1 Mediated by Nuclear Translocation Nrf2 in Caco-2 Co-Cultured THP-1 Macrophages
3. Materials and Methods
3.1. Materials
3.2. Isolation of Scalarane Sesterterpenes from Sponge Genus Phyllopspongia
3.3. Cell Cultures
3.4. Differentiation of THP-1 to Macrophages
3.5. In Vitro Co-Culture Model of Intestine
3.6. TEER Measurement
3.7. Cell Viability Assay
3.8. Measurement of NO and PGE2 Production
3.9. Measurement of IL-1β, TNF-α and IL-6 Production
3.10. Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
3.11. Western Blot
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Chk2 | Checkpoint kinase 2 |
| LPS | lipopolysaccharide |
| PGE2 | Prostaglandin 2 |
| NO | Nitric oxide |
| iNOS | Inducible nitric oxide synthase |
| COX | Cyclooxygenase |
| HO-1 | Heme oxygenase-1 |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| Nrf2 | Nuclear factor erythroid 2 related factor 2 |
| MAPK | Mitogen-activated protein kinase |
| HRESIMS | High-resolution electrospray ionization Mass spectrometry |
| QTOF | Quadrupole time of flight mass spectrometer |
| HSQC | Heteronuclear single quantum coherence |
| COSY | Correlation spectroscopy |
| HMBC | Heteronuclear multiple bond correlation |
References
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, E.V.; Sandborn, W.J. Epidemiology of inflammatory bowel disease. Gastroenterol. Clin. N. Am. 2002, 31, 1–20. [Google Scholar] [CrossRef]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Kleiveland, C.R. Chapter 18: Co-culture Caco-2/Immune Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Kämpfer, A.A.; Urbán, P.; Gioria, S.; Kanase, N.; Stone, V.; Kinsner-Ovaskainen, A. Development of an in vitro co-culture model to mimic the human intestine in healthy and diseased state. Toxicol. In Vitro 2017, 45, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Rubas, W.; Jezyk, N.; Grass, G.M. Comparison of the Permeability Characteristics of a Human Colonic Epithelial (Caco-2) Cell Line to Colon of Rabbit, Monkey, and Dog Intestine and Human Drug Absorption. Pharm. Res. 1993, 10, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Lennernäs, H.; Palm, K.; Fagerholm, U.; Artursson, P. Comparison between active and passive drug transport in human intestinal epithelial (Caco-2) cells in vitro and human jejunum in vivo. Int. J. Pharm. 1996, 127, 103–107. [Google Scholar] [CrossRef]
- Shinde, P.; Banerjee, P.; Mandhare, A. Marine natural products as source of new drugs: A patent review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, M.; Wang, H.; Crews, P. Secondary metabolites from the marine sponge genus. Mar. Drugs 2017, 15, 12. [Google Scholar] [CrossRef]
- Wan, Y.Q.; Su, J.Y.; Zeng, L.M.; Liu, Q. The secondary metabolites of marine sponge Phyllospongia foliascens. Acta Sci. Nat. Univ. Sunyatseni 1998, 37, 81–84. [Google Scholar]
- Hassan, M.H.; Rateb, M.E.; Hetta, M.; Abdelaziz, T.A.; Sleim, M.A.; Jaspars, M.; Mohammed, R. Scalarane sesterterpenes from the Egyptian Red Sea sponge Phyllospongia lamellosa. Tetrahedron 2015, 71, 577–583. [Google Scholar] [CrossRef]
- Mohapatra, B.R.; Bapuji, M.; Sree, A. Antifungal efficacy of bacteria isolated from marine sedentary organisms. Folia Microbiol. 2002, 47, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.H.; Liu, Y.C.; Su, J.H.; El-Shazly, M.; Wu, C.F.; Du, Y.C.; Hsu, Y.M.; Yang, J.C.; Weng, M.K.; Chou, C.H.; et al. Antileukemic scalarane sesterterpenoids and meroditerpenoid from Carteriospongia (Phyllospongia) sp., induce apoptosis via dual inhibitory effects on topoisomerase II and Hsp90. Sci. Rep. 2016, 6, 36170. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Canning, C.; Wang, K.; Zhu, W.; Yang, F.; Zhang, Y.; Zhou, K. Antibacterial Activity of 2-(3’,5’-Dibromo-2’-methoxyphenoxy)-3,5- dibromophenol Isolated from Phyllospongia papyracea. Nat. Prod. Commun. 2017, 12, 567–569. [Google Scholar] [CrossRef]
- Chang, L.C.; Otero-Quintero, S.; Nicholas, G.M.; Bewley, C.A. Phyllolactones A-E: New bishomoscalarane sesterterpenes from the marine sponge Phyllospongia lamellosa. Tetrahedron 2001, 57, 5731–5738. [Google Scholar] [CrossRef]
- Fu, X.; Zeng, L.M.; Su, J.Y.; Francis, J.S. A new sesterterpene from the sponge Phyllospongia foliascens. Chin. Chem. Lett. 1991, 2, 543–544. [Google Scholar]
- Zeng, L.; Fu, X.; Su, J.Y.; Pordesimo, E.O.; Traeger, S.C.; Schmitz, J. Novel bishomoscarlarane sesterpenes from the sponge Phyllospongia foliascens. J. Nat. Prod. 1991, 54, 421–427. [Google Scholar] [CrossRef]
- Pettus, B.J.; Bielawski, J.; Porcelli, A.M.; Reames, D.L.; Johnson, K.R.; Morrow, J.; Chalfant, C.E.; Obeid, L.M.; Hannun, Y.A. The sphingosine kinase 1/sphingosine-1-phosphate pathway mediates COX-2 induction and PGE2 production in response to TNF-alpha. FASEB J. 2003, 17, 1411–1421. [Google Scholar] [CrossRef]
- Pan, M.-H.; Hsieh, M.-C.; Hsu, P.-C.; Ho, S.-Y.; Lai, C.-S.; Wu, H.; Sang, S.; Ho, C.-T. 6-Shogaol suppressed lipopolysaccharide-induced up-expression of iNOS and COX-2 in murine macrophages. Mol. Nutr. Food Res. 2008, 52, 1467–1477. [Google Scholar] [CrossRef]
- Wilson, K.; Ramanujam, K.; Mobley, H.; Musselman, R.; James, S.; Meltzer, S. Helicobacter pylori stimulates inducible nitric oxide synthase expression and activity in a murine macrophage cell line. Gastroenterology 1996, 111, 1524–1533. [Google Scholar] [CrossRef]
- Choi, E.-M.; Hwang, J.-K. Effects of Morus alba leaf extract on the production of nitric oxide, prostaglandin E2 and cytokines in RAW264.7 macrophages. Fitoterapia 2005, 76, 608–613. [Google Scholar] [CrossRef] [PubMed]
- MacDermott, R.P. Chemokines in the inflammatory bowel diseases. J. Clin. Immunol. 1999, 19, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Reinecker, H.C.; Steffen, M.; Witthoeft, T.; Pflueger, I.; Schreiber, S.; MacDermott, R.P.; Raedler, A. Enhanced secretion of tumour necrosis factor-alpha, IL-6, and IL-1 beta by isolated lamina propria mononuclear cells from patients with ulcerative colitis and Crohn’s disease. Clin. Exp. Immunol. 1993, 94, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto-Furusho, J.K. Inflammatory bowel disease therapy: Blockade of cytokines and cytokine signaling pathways. Curr. Opin. Gastroenterol. 2018, 34, 187–193. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Nuclear factor kappa B regulation of proinflammatory cytokines in human gestational tissues in vitro. Biol. Reprod. 2002, 67, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Takada, Y.; Shishodia, S.; Gutierrez, A.M.; Oommen, O.V.; Ichikawa, H.; Baba, Y.; Kumar, A. Nuclear transcription factor NF-kappa B: Role in biology and medicine. Indian J. Exp. Biol. 2004, 42, 341–353. [Google Scholar]
- Economou, J.S.; Rhoades, R.; Essner, W.H.; McBride, J.C.; Morton, D.L. Genetic analysis of the human tumor necrosis factor a/cachectin promoter region in a macrophage cell line. J. Exp. Med. 1989, 170, 321–326. [Google Scholar] [CrossRef]
- Appleby, S.B.; Ristimäki, A.; Neilson, K.; Narko, K.; Hla, T. Structure of the human cyclo-oxygenase-2 gene. Biochem. J. 1994, 302, 723–727. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, I.T.; Won, J.H. Anti-inflammatory activities of ent16alphaH,17-hydroxy-kauran-19-oic acid isolated from the roots of Siegesbeckia pubescens are due to the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via NF-kappaB inactivation. Eur. J. Pharmacol. 2007, 558, 185–193. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Saegusa, M.; Hashimura, M.; Kuwata, T. Pin1 acts as a modulator of cell proliferation through alteration in NF-kappaB but not beta-catenin/TCF4 signalling in a subset of endometrial carcinoma cells. J. Pathol. 2010, 222, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Bach, F.H.; Alam, J.; Soares, M.; Lu, H.T.; Wysk, M.; Davis, R.J.; Flavell, R.A.; Choi, A.M. Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat. Med. 2000, 6, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-S.; Li, B.; Im, N.-K.; Kim, Y.-C.; Jeong, G.-S. 4,2′,5′-Trihydroxy-4′-methoxychalcone from Dalbergia odorifera exhibits anti-inflammatory properties by inducing heme oxygenase-1 in murine macrophages. Int. Immunopharmacol. 2013, 16, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, C.; Liu, D.; Han, M.K.; Wang, L.; Merlin, D. Oral Delivery of Nanoparticles Loaded with Ginger Active Compound, 6-Shogaol, Attenuates Ulcerative Colitis and Promotes Wound Healing in a Murine Model of Ulcerative Colitis. J. Crohns Colitis 2018, 12, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; Talero, E.; Rodríguez-Luna, A.; García-Mauriño, S.; Motilva, V. Anti-inflammatory effects of an oxylipin-containing lyophilised biomass from a microalga in a murine recurrent colitis model. Br. J. Nutr. 2016, 116, 2044–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daigneault, M.; Perston, J.A.; Marriott, H.M.; Whyte, M.K.B.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef] [PubMed]
- Hetrick, E.M.; Schoenfisch, M.H. Analytical chemistry of nitric oxide. Annu. Rev. Anal. Chem. 2009, 2, 409–433. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1H, mult (J Hz) | 13C | No. | 1H, mult (J Hz) | 13C |
|---|---|---|---|---|---|
| 1 | 0.79, m; 1.58, m | 39.8, t | 15 | 1.48, m | 30.2, t |
| 2 | 1.39, m; 1.49, m | 18.1, t | 1.95, ddd (12.7, 4.9, 2.5) | ||
| 3 | 0.84, m; 1.67, m | 36.4, t | 16 | 3.77, td (10.5, 5.1) | 70.1, d |
| 4 | 36.1, s | 17 | 1.64, dd (12.7, 10.5) | 54.8, d | |
| 5 | 0.86, m | 58.6, d | 18 | 2.28, m | 46.8, d |
| 6 | 1.47, m; 1.57, m | 18.0, t | 19 | 0.80, s | 28.5, q |
| 7 | 0.95, td (12.5, 3.7) | 41.9, t | 20 | 1.18, m; 1.53, dd (14.2, 7.6) | 24.5, t |
| 1.83, dt (12.5, 3.2) | 21 | 0.88, s | 16.4, q | ||
| 8 | 37.6, s | 22 | 1.05, s | 16.8, q | |
| 9 | 1.17, dd (13.5, 2.5) | 61.3, d | 23 | 1.12, s | 13.7, q |
| 10 | 38.1, s | 24 | 104.4, s | ||
| 11 | 2.23, dd (13.5, 2.5); 2.60, t (13.5) | 34.8, t | 25 | 3.63, t (8.6); 4.26, t (8.6) | 67.5, t |
| 12 | 215.3, s | 263 | 1.49, s | 21.9, q | |
| 13 | 51.4, s | 27 | 0.75, t (7.6) | 8.8, q | |
| 14 | 1.28, m | 58.7, d | OCH | 3.25, s | 48.4, q |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.M.; Kim, N.-H.; Lee, S.; Kim, Y.N.; Heo, J.D.; Jeong, E.J.; Rho, J.-R. Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine. Mar. Drugs 2019, 17, 634. https://doi.org/10.3390/md17110634
Lee SM, Kim N-H, Lee S, Kim YN, Heo JD, Jeong EJ, Rho J-R. Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine. Marine Drugs. 2019; 17(11):634. https://doi.org/10.3390/md17110634
Chicago/Turabian StyleLee, Seon Min, Na-Hyun Kim, Sangbum Lee, Yun Na Kim, Jeong Doo Heo, Eun Ju Jeong, and Jung-Rae Rho. 2019. "Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine" Marine Drugs 17, no. 11: 634. https://doi.org/10.3390/md17110634
APA StyleLee, S. M., Kim, N. -H., Lee, S., Kim, Y. N., Heo, J. D., Jeong, E. J., & Rho, J. -R. (2019). Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine. Marine Drugs, 17(11), 634. https://doi.org/10.3390/md17110634