Quantitative Structure-Activity Relationship Model to Predict Antioxidant Effects of the Peptide Fraction Extracted from a Co-Culture System of Chlorella pyrenoidosa and Yarrowia lipolytica


Abstract
:1. Introduction
2. Results and Discussion
2.1. Amino Acid Composition
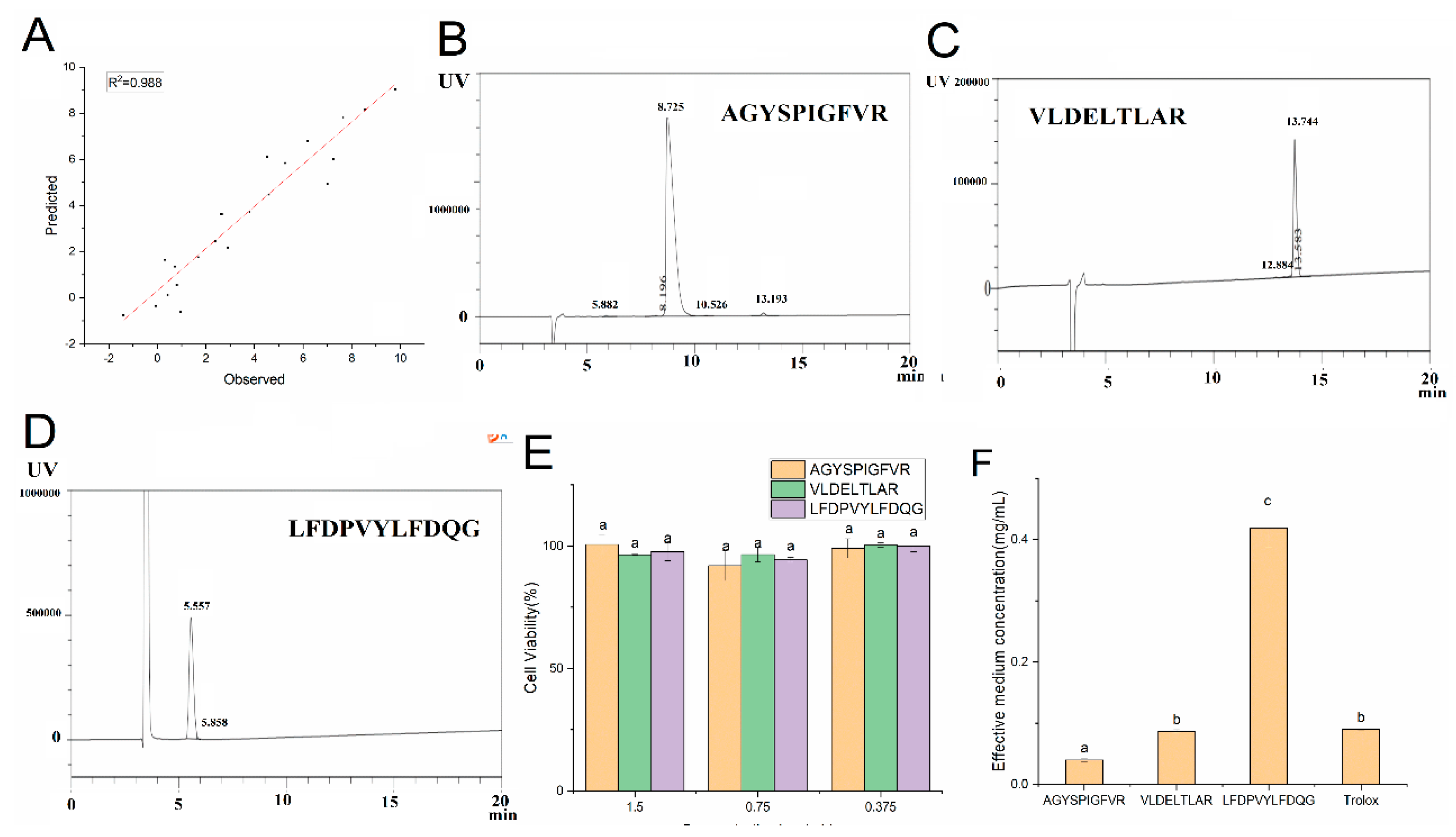
2.2. EHP Sequence
2.3. In Vitro Antioxidant Assay
2.4. Effect of EHP on Cell Viability
2.5. Effects of EHP on Reactive Oxygen Species (ROS) Generation
2.6. Effect of EHP on Malondialdehyde (MDA) Levels
2.7. Effect of EHP on Reduced Glutathione (GSH) and Oxidized Glutathione (GSSG) Levels
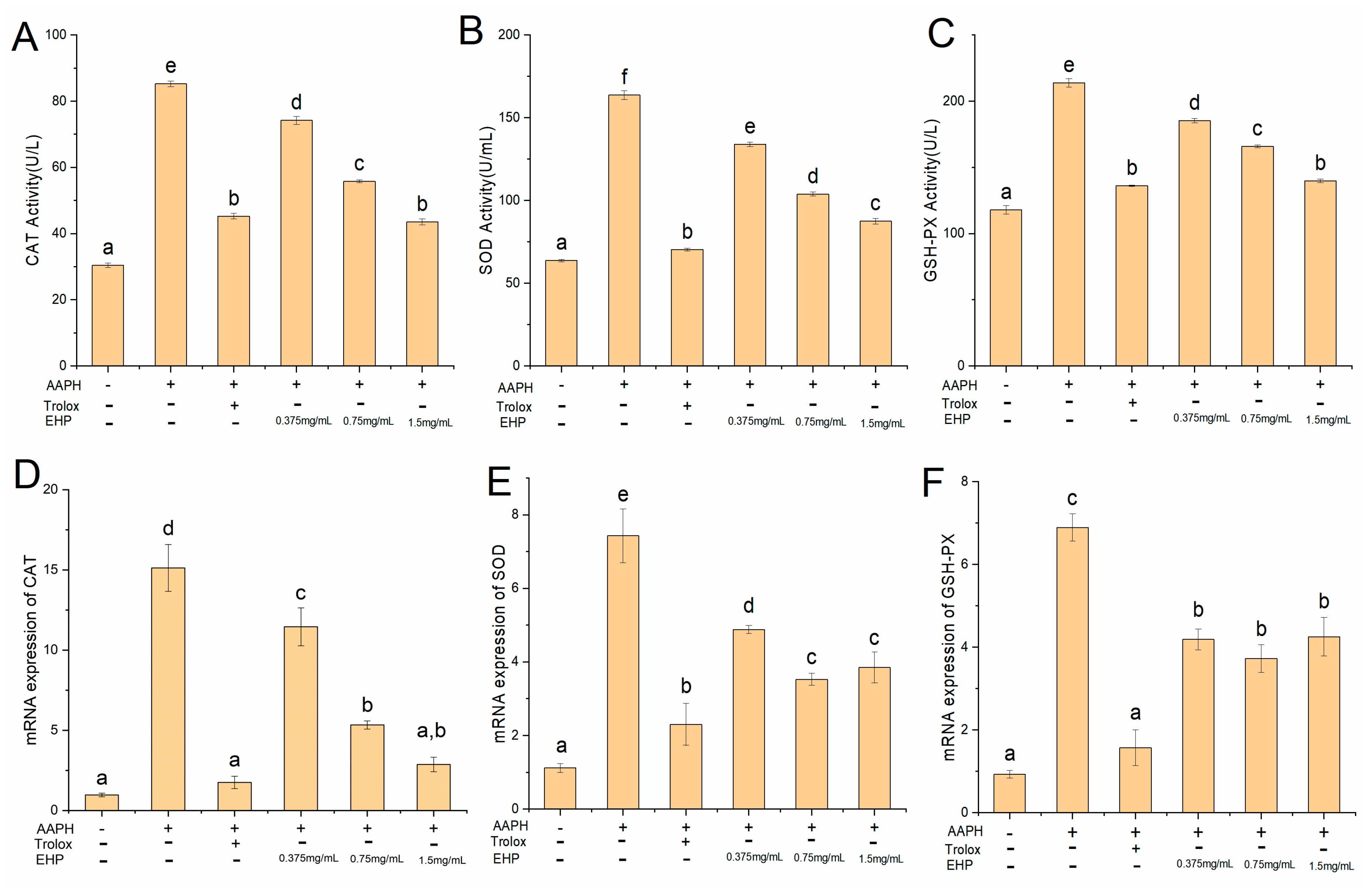
2.8. Effect of EHP on Superoxide Dismutase (SOD), Catalase (CAT), and Glutathione Peroxidase (GSH-PX) Activities
2.9. mRNA and Protein Expression of Nrf2, Kelch-Like ECH-Associated Protein-1 (Keap1), CAT, SOD, and GSH-PX
2.10. QSAR Model Establishment and Verification
3. Materials and Methods
3.1. Materials
3.2. EHP Preparation from Symbiotic Culture
3.3. Qualitative Proteomics Assay Using Liquid Chromatography (LC)-MS/MS
3.4. Amino Acid Analysis
3.5. In Vitro Evaluation of Free Radical Scavenging Capabilities
3.6. Determination of Antioxidative Abilities of EHP in HepG2
3.6.1. Cell Culture
3.6.2. Cell Viability Assay
3.6.3. Measurement of ROS Production
3.6.4. Determination of MDA, GSH, and GSSG Levels
3.6.5. Determination of SOD, CAT, and GSH-PX Levels
3.6.6. Analysis of mRNA Expression of SOD, CAT, GSH-PX, Keap1, and Nrf2
3.6.7. Analysis of Protein Levels of SOD, CAT, GSH-PX, Keap1, and Nrf2
3.7. Quantitative Structure–Activity Relationship Modeling
3.8. Validation Experiments
3.9. Median Effective Concentration (EC50) Assay
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Aboushanab, R.A.I.; Ji, M.K.; Kim, H.C.; Paeng, K.J.; Jeon, B.H. Microalgal species growing on piggery wastewater as a valuable candidate for nutrient removal and biodiesel production. J. Environ. Manag. 2013, 115, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, J.A. The protein quality, digestibility and composition of Chlorella 71105. J. Food Sci. 2010, 28, 229–232. [Google Scholar] [CrossRef]
- Cai, X.; Yang, Q.; Wang, S. Antioxidant and hepatoprotective effects of pigment-protein complex from Chlorella vulgaris on carbon tetrachloride-induced liver damage in vivo. RSC Adv. 2015, 116, 96097–96104. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and Antioxidant Activities of the Peptide Fraction from Algae Protein Waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Guo, Z.; Tong, Y.W. The interactions between Chlorella vulgaris and algal symbiotic bacteria under photoautotrophic and photoheterotrophic conditions. J. Appl. Phycol. 2014, 26, 1483–1492. [Google Scholar] [CrossRef]
- González, C.; Marciniak, J.; Villaverde, S.; García-Encina, P.A.; Murioz, R. Microalgae-based processes for the biodegradation of pretreated piggery wastewaters. Appl. Microbiol. Biotechnol. 2008, 80, 891–898. [Google Scholar] [CrossRef]
- Liu, J.; Willför, S.; Xu, C. A review of bioactive plant polysaccharides: Biological activities, functionalization, and biomedical applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 31–61. [Google Scholar] [CrossRef]
- Noguchi, N.; Yanagita, T.; Rahman, S.M.; Ando, Y. Chlorella Protein Hydrolysate Attenuates Glucose Metabolic Disorder and Fatty Liver in High-fat Diet-induced Obese Mice. J. Oleo Sci. 2016, 65, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.C.; Kang, N.; Kim, E.; Min, C.K.; Lee, S.H.; Kang, S.M.; Lee, J.B.; Jeon, B.T.; Kim, S.K.; Park, S.J. A novel angiotensin I-converting enzyme (ACE) inhibitory peptide from a marine Chlorella ellipsoidea and its antihypertensive effect in spontaneously hypertensive rats. Process Biochem. 2012, 47, 2005–2011. [Google Scholar] [CrossRef]
- Tomás, H.; Juan, G. Hydroxyl radical reactions and the radical scavenging activity of β-carboline alkaloids. Food Chem. 2015, 172, 640–649. [Google Scholar]
- Nikoo, M.; Benjakul, S.; Ehsani, A.; Jing, L.; Wu, F.; Na, Y.; Xu, B.; Jin, Z.; Xu, X. Antioxidant and cryoprotective effects of a tetrapeptide isolated from Amur sturgeon skin gelatin. J. Funct. Foods 2014, 7, 609–620. [Google Scholar] [CrossRef]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Muller, A.T.; Kaymaz, A.C.; Gabernet, G.; Posselt, G.; Wessler, S.; Hiss, J.A.; Schneider, G. Sparse Neural Network Models of Antimicrobial Peptide-Activity Relationships. Qsar Comb. Sci. 2016, 35, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Omar, D.; Mohammad, G. Exploring QSARs for inhibitory activity of non-peptide HIV-1 protease inhibitors by GA-PLS and GA-SVM. Chem. Biol. Drug Des. 2010, 75, 506–514. [Google Scholar]
- Chuan, S.; Fang, T.J.; Tung-Kung, W.; Peng-Hsiang, L. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar]
- Rajabzadeh, M.; Pourashouri, P.; Shabanpour, B.; Alishahi, A. Amino acid composition, antioxidant and functional properties of protein hydrolysates from the roe of rainbow trout (Oncorhynchus mykiss). Int. J. Food Sci. Technol. 2018, 53, 313–319. [Google Scholar] [CrossRef]
- Olsen, J.V.; Ong, S.E.; Mann, M. Trypsin Cleaves Exclusively C-terminal to Arginine and Lysine Residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Adebiyi, A.P.; Adebiyi, A.O.; Yamashita, J.; Ogawa, T.; Muramoto, K. Purification and characterization of antioxidative peptides derived from rice bran protein hydrolysates. Eur. Food Res. Technol. 2009, 228, 553. [Google Scholar] [CrossRef]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Jung, W.K.; Je, J.Y.; Kim, S.K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Wu, J.; Sun, B.; Luo, X.; Zhao, M.; Zheng, F.; Sun, J.; Li, H.; Sun, X.; Huang, M. Cytoprotective effects of a tripeptide from Chinese Baijiu against AAPH-induced oxidative stress in HepG2 cells via Nrf2 signaling. Rsc Adv. 2018, 8, 10898–10906. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Paluszczak, J.; Szaefer, H.; Baer-Dubowska, W. The activation of the Nrf2/ARE pathway in HepG2 hepatoma cells by phytochemicals and subsequent modulation of phase II and antioxidant enzyme expression. J. Physiol. Biochem. 2015, 71, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Samatiwat, P.; Prawan, A.; Senggunprai, L.; Kukongviriyapan, V. Taxifolin Exerts Cytoprotective Effect by Activation of Nrf2-ARE Signaling Pathway in HepG2 cells. Srinagarind Med. J. (SMJ) 2014, 29, 122–125. [Google Scholar]
- Hempel, N.; Melendez, J.A. Intracellular redox status controls membrane localization of pro- and anti-migratory signaling molecules. Redox Biol. 2014, 2, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41. [Google Scholar] [CrossRef]
- Wijeratne, S.S.K.; Cuppett, S.L.; Schlegel, V. Hydrogen Peroxide Induced Oxidative Stress Damage and Antioxidant Enzyme Response in Caco-2 Human Colon Cells. J. Agric. Food Chem. 2005, 53, 8768–8774. [Google Scholar] [CrossRef]
- Ham, H.; Yoon, S.W.; Kim, I.H.; Kwak, J.; Lee, J.S.; Jeong, H.S.; Lee, J. Protective effects of unsaponifiable matter from rice bran on oxidative damage by modulating antioxidant enzyme activities in HepG2 cells. LWT Food Sci. Technol. 2015, 61, 602–608. [Google Scholar] [CrossRef]
- Korotkova, E.I.; Avramchik, O.A.; Kagiya, T.V.; Karbainov, Y.A.; Tcherdyntseva, N.V. Study of antioxidant properties of a water-soluble Vitamin E derivative-tocopherol monoglucoside (TMG) by differential pulse voltammetry. Talanta 2004, 63, 729–734. [Google Scholar] [CrossRef]
- Takafumi, S.; Masayuki, Y. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar]
- Uruno, A.; Yagishita, Y.; Yamamoto, M. The Keap1–Nrf2 system and diabetes mellitus. Arch. Biochem. Biophys. 2015, 566, 76–84. [Google Scholar] [CrossRef]
- Scolastici, C.; Lima, R.O.A.; Barbisan, L.F.; Ferreira, A.L.A.; Ribeiro, D.A.; Salvadori, D.M.F. Antigenotoxicity and antimutagenicity of lycopene in HepG2 cell line evaluated by the comet assay and micronucleus test. Toxicol. Vitro 2008, 22, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jing, W.; Liu, Y.; Gong, L.; Sun, B. Wheat bran feruloyl oligosaccharides ameliorate AAPH-induced oxidative stress in HepG2 cells via Nrf2 signalling. J. Funct. Foods 2016, 25, 333–340. [Google Scholar] [CrossRef]
- Ken, I.; Tong, K.I.; Masayuki, Y. Molecular mechanism activating Nrf2-Keap1 pathway in regulation of adaptive response to electrophiles. Free Radic. Biol. Med. 2004, 36, 1208–1213. [Google Scholar]
- Matsui, R.; Honda, R.; Kanome, M.; Hagiwara, A.; Matsuda, Y.; Togitani, T.; Ikemoto, N.; Terashima, M. Designing antioxidant peptides based on the antioxidant properties of the amino acid side-chains. Food Chem. 2018, 245, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.J.; Jung, W.K.; Kim, S.K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour. Technol. 2008, 99, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.; Ghosh, G. Amino acid composition, antioxidant and functional properties of protein hydrolysates from Cucurbitaceae seeds. J. Food Sci. Technol. 2017, 54, 4162–4172. [Google Scholar] [CrossRef]
- Doktorovová, S.; Santos, D.L.; Costa, I.; Andreani, T.; Souto, E.B.; Silva, A.M. Cationic solid lipid nanoparticles interfere with the activity of antioxidant enzymes in hepatocellular carcinoma cells. Int. J. Pharm. 2014, 471, 18–27. [Google Scholar] [CrossRef]
- Wenzhen, L.; Zhengxiang, N.; Luying, C.; Qingyi, W.; Erdong, Y.; Jiguo, Y.; Jiaoyan, R. Intracellular antioxidant detoxifying effects of diosmetin on 2,2-azobis(2-amidinopropane) dihydrochloride (AAPH)-induced oxidative stress through inhibition of reactive oxygen species generation. J. Agric. Food Chem. 2014, 62, 8648–8654. [Google Scholar]
- Wang, J.; Sun, B.; Cao, Y.; Tian, Y. Protection of wheat bran feruloyl oligosaccharides against free radical-induced oxidative damage in normal human erythrocytes. Food Chem. Toxicol. 2009, 47, 1591–1599. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid Redox Signal 2015, 7, 385–394. [Google Scholar] [CrossRef]
- Mei, H.; Liao, Z.; Zhou, Y.; Li, S. A new set of amino acid descriptors and its application in peptide QSARs. Pept. Sci. 2010, 80, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, J.; Zhang, X. Extraction of intracellular protein from Chlorella pyrenoidosa using a combination of ethanol soaking, enzyme digest, ultrasonication and homogenization techniques. Bioresour. Technol. 2017, 247, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. MCP 2012, 11, M111–010587. [Google Scholar] [CrossRef]
- Liu, H.; Wu, H.; Wang, Q. The Variations, Including Structures and Attenuation to Hemolysis, of Peptide Purified from Dendrobium aphyllum During In Vitro Gastro-Intestinal Digestion and Caco-2 Uptake and Transportation. Int. J. Pept. Res. Ther. 2018, 25, 1–13. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Xie, Z.; Huang, J.; Xu, X.; Jin, Z. Antioxidant activity of peptides isolated from alfalfa leaf protein hydrolysate. Food Chem. 2008, 111, 370–376. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Ramos, S.; Cordero-Herrera, I.; Recio, I.; Goya, L.; Hernández-Ledesma, B. In vitro chemo-protective effect of bioactive peptide lunasin against oxidative stress in human HepG2 cells. Food Res. Int. 2014, 62, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.L.; Hai, L.R. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8970. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, Y.-W.; Li, B. Characterization of structure–antioxidant activity relationship of peptides in free radical systems using QSAR models: Key sequence positions and their amino acid properties. J. Theor. Biol. 2013, 318, 29–43. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Total (nmol/mg) | Free (nmol/mg) |
|---|---|---|
| Asp | 17.02 ± 1.38 ab | ND c |
| Thr | 8.10 ± 0.02 a | ND c |
| Ser | 6.40 ± 0.1 a | ND c |
| Glu | 18.71 ± 1.84 b | 0.038 ± 0.02 b |
| Gly | 13.42 ± 1.27 a | 0.29 ± 0.09 a |
| Ala | 22.20 ± 1.31 a | ND c |
| Cys | ND c | ND c |
| Val | 14.55 ± 0.41 a | ND c |
| Met | ND c | ND c |
| Ile | 11.03 ± 1.12 b | 0.21 ± 0.01 a |
| Leu | 15.00 ± 1.13 a | 0.69 ± 0.02 a |
| Tyr | 6.21 ± 0.22 a | 0.29 ± 0.01 a |
| Phe | 2.97 ± 0.59 b | 0.40 ± 0.01 a |
| His | 0.26 ± 0.01 a | ND c |
| Lys | 10.06 ± 0.41 b | 0.45 ± 0.04 b |
| Arg | 8.80 ± 1.64 b | 0.60 ± 0.02 a |
| Pro | 4.91 ± 0.3 a | ND c |
| HAAsd | 76.87 ± 5.08 a | 1.59 ± 0.05 a |
| NCAAse | 35.73 ± 3.22 b | 0.038 ± 0.02 a |
| AAAsf | 9.18 ± 0.81 a | 0.69 ± 0.02 a |
| Sum | 159.64 ± 11.75 b | 2.96 ± 0.24 a |
| No | Sequence | Activity |
|---|---|---|
| 1 | WY | 7.67 |
| 2 | WYS | 4.45 |
| 3 | WYSL | 4.52 |
| 4 | WNIP | 15.47 |
| 5 | GWNI | 13.9 |
| 6 | YVEEL | 0.799 |
| 7 | MHIRL | 0.306 |
| 8 | SALAM | 2.66 |
| 9 | WYSLA | 4.59 |
| 10 | AGWNI | 8.55 |
| 11 | LGFEY | 9.79 |
| 12 | GWNIP | 6.19 |
| 13 | WYSLAM | 7 |
| 14 | AGWNIP | 7.64 |
| 15 | LGFEYY | 7.25 |
| 16 | VIPMGL | 2.89 |
| 17 | AGWNIPIGT | 5.25 |
| 18 | LSKAQSDFG | -1.4 |
| 19 | YAEERYPIL | 3.8 |
| 20 | LVEKGDVAFI | -0.07 |
| 21 | WYSLAMAASDI | 2.621 |
| 22 | IEWEGIESGSVEQA | 0.73 |
| 23 | AIEWEGIESGSVEQA | 0.95 |
| 24 | IAAEVYEHTEGSTTSY | 2.39 |
| 25 | PIAAEVYEHTEGSTTSY | 1.68 |
| 26 | IANNEADAISLDGGQVFEAG | 0.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Li, S.; Zhong, Y.; Liu, J.; Liu, H.; Cheng, J.; Ma, L.; Huang, Y.; Cai, X.; Liu, H.; et al. Quantitative Structure-Activity Relationship Model to Predict Antioxidant Effects of the Peptide Fraction Extracted from a Co-Culture System of Chlorella pyrenoidosa and Yarrowia lipolytica. Mar. Drugs 2019, 17, 633. https://doi.org/10.3390/md17110633
Liu H, Li S, Zhong Y, Liu J, Liu H, Cheng J, Ma L, Huang Y, Cai X, Liu H, et al. Quantitative Structure-Activity Relationship Model to Predict Antioxidant Effects of the Peptide Fraction Extracted from a Co-Culture System of Chlorella pyrenoidosa and Yarrowia lipolytica. Marine Drugs. 2019; 17(11):633. https://doi.org/10.3390/md17110633
Chicago/Turabian StyleLiu, Huifan, Sufen Li, Yuming Zhong, Jianliang Liu, Hui Liu, Jian Cheng, Lukai Ma, Yuqing Huang, Xuanyi Cai, Haijun Liu, and et al. 2019. "Quantitative Structure-Activity Relationship Model to Predict Antioxidant Effects of the Peptide Fraction Extracted from a Co-Culture System of Chlorella pyrenoidosa and Yarrowia lipolytica" Marine Drugs 17, no. 11: 633. https://doi.org/10.3390/md17110633
APA StyleLiu, H., Li, S., Zhong, Y., Liu, J., Liu, H., Cheng, J., Ma, L., Huang, Y., Cai, X., Liu, H., Zheng, J., Su, Z., & Wang, Q. (2019). Quantitative Structure-Activity Relationship Model to Predict Antioxidant Effects of the Peptide Fraction Extracted from a Co-Culture System of Chlorella pyrenoidosa and Yarrowia lipolytica. Marine Drugs, 17(11), 633. https://doi.org/10.3390/md17110633