Sinulariolide Inhibits Gastric Cancer Cell Migration and Invasion through Downregulation of the EMT Process and Suppression of FAK/PI3K/AKT/mTOR and MAPKs Signaling Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Sinulariolide Treatment on Cell Survival of Gastric Cancer Cells
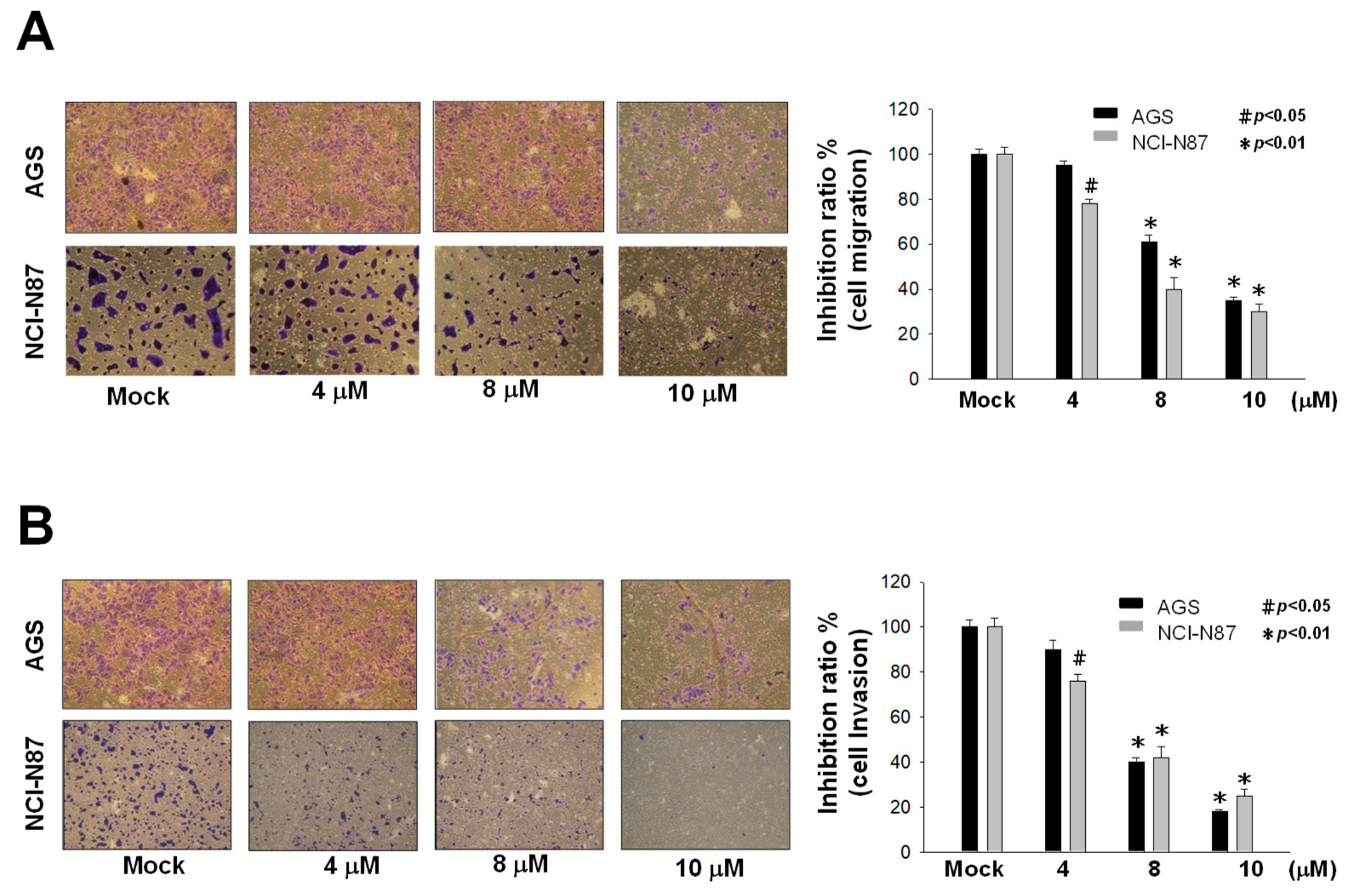
2.2. Inhibition of Metastasis and Invasion of AGS and NCI-N87 Cells by Sinulariolide
2.3. Effects of Sinulariolide on Expressions of MMP-2, MMP-9, uPA, TIMP-1, and TIMP-2
2.4. Effects of Sinulariolide Treatment on the Intracellular FAK/PI3K/AKT/mTOR Signaling Pathway
2.5. Sinulariolide Inhibits MAPKs Signaling Pathways, Affecting Cell Metastasis and Invasion
2.6. Sinulariolide Suppressed Epithelial–Mesenchymal Transition
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Drug Treatment, and Cell Viability Assay
4.2. Cell Migration and Invasion Analysis by Transwell Assay
4.3. Gelatin Zymography Assay
4.4. Protein Concentration Analysis and Western Blot Assay
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dai, W.; Shen, G.; Qiu, J.; Zhao, X.; Gao, Q. Aberrant expression of b7-h3 in gastric adenocarcinoma promotes cancer cell metastasis. Oncol. Rep. 2014, 32, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Ajani, J.A.; Faust, J.; Ikeda, K.; Yao, J.C.; Anbe, H.; Carr, K.L.; Houghton, M.; Urrea, P. Phase i pharmacokinetic study of s-1 plus cisplatin in patients with advanced gastric carcinoma. J. Clin. Oncol. 2005, 23, 6957–6965. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, A.; Yoshida, S.; Saijo, N. Disparities in gastric cancer chemotherapy between the east and west. J. Clin. Oncol. 2006, 24, 2188–2196. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, W.; Narahara, H.; Hara, T.; Takagane, A.; Akiya, T.; Takagi, M.; Miyashita, K.; Nishizaki, T.; Kobayashi, O.; Takiyama, W. S-1 plus cisplatin versus s-1 alone for first-line treatment of advanced gastric cancer (spirits trial): A phase iii trial. Lancet Oncol. 2008, 9, 215–221. [Google Scholar] [CrossRef]
- Ohtsu, A. Current status and future prospects of chemotherapy for metastatic gastric cancer: A review. Gastric Cancer 2005, 8, 95–102. [Google Scholar] [CrossRef]
- Yamashita, K.; Sakuramoto, S.; Kikuchi, S.; Katada, N.; Kobayashi, N.; Watanabe, M. Validation of staging systems for gastric cancer. Gastric Cancer 2008, 11, 111–118. [Google Scholar] [CrossRef]
- Park, J.-M.; Kim, Y.H. Current approaches to gastric cancer in korea. Gastrointest. Cancer Res. GCR 2008, 2, 137. [Google Scholar]
- Radhika, P. Chemical constituents and biological activities of the soft corals of genus cladiella: A review. Biochem. Syst. Ecol. 2006, 34, 781–789. [Google Scholar] [CrossRef]
- Li, H.-H.; Su, J.-H.; Chiu, C.-C.; Lin, J.-J.; Yang, Z.-Y.; Hwang, W.-I.; Chen, Y.-K.; Lo, Y.-H.; Wu, Y.-J. Proteomic investigation of the sinulariolide-treated melanoma cells a375: Effects on the cell apoptosis through mitochondrial-related pathway and activation of caspase cascade. Mar. Drugs 2013, 11, 2625–2642. [Google Scholar] [CrossRef]
- Neoh, C.-A.; Wang, R.Y.-L.; Din, Z.-H.; Su, J.-H.; Chen, Y.-K.; Tsai, F.-J.; Weng, S.-H.; Wu, Y.-J. Induction of apoptosis by sinulariolide from soft coral through mitochondrial-related and p38mapk pathways on human bladder carcinoma cells. Mar. Drugs 2012, 10, 2893–2911. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Su, J.-H.; Tsao, C.-Y.; Hung, C.-T.; Chao, H.-H.; Lin, J.-J.; Liao, M.-H.; Yang, Z.-Y.; Huang, H.H.; Tsai, F.-J. Sinulariolide induced hepatocellular carcinoma apoptosis through activation of mitochondrial-related apoptotic and perk/eif2α/atf4/chop pathway. Molecules 2013, 18, 10146–10161. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.; Wu, Y.-J.; Liu, Z.; Chuang, C.; Huang, H.; Kuo, S. Anticancer effects of sinulariolide-conjugated hyaluronan nanoparticles on lung adenocarcinoma cells. Molecules 2016, 21, 297. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Li, Y.R.; Huang, W.Y.; Su, J.H.; Chan, H.L.; Lin, S.H.; Liu, C.S.; Lin, S.C.; Lin, C.C.; Lin, C.H. Sinulariolide suppresses lps-induced phenotypic and functional maturation of dendritic cells. Mol. Med. Rep. 2017, 16, 6992–7000. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-J.; Neoh, C.-A.; Tsao, C.-Y.; Su, J.-H.; Li, H.-H. Sinulariolide suppresses human hepatocellular carcinoma cell migration and invasion by inhibiting matrix metalloproteinase-2/-9 through mapks and pi3k/akt signaling pathways. Int. J. Mol. Sci. 2015, 16, 16469–16482. [Google Scholar] [CrossRef]
- Cheng, T.-C.; Din, Z.-H.; Su, J.-H.; Wu, Y.-J.; Liu, C.-I. Sinulariolide suppresses cell migration and invasion by inhibiting matrix metalloproteinase-2/-9 and urokinase through the pi3k/akt/mtor signaling pathway in human bladder cancer cells. Mar. Drugs 2017, 15, 238. [Google Scholar] [CrossRef]
- Yeh, C.B.; Hsieh, M.J.; Hsieh, Y.H.; Chien, M.H.; Chiou, H.L.; Yang, S.F. Antimetastatic effects of norcantharidin on hepatocellular carcinoma by transcriptional inhibition of mmp-9 through modulation of nf-kb activity. PLoS ONE 2012, 7, e31055. [Google Scholar] [CrossRef]
- Yang, S.F.; Chen, M.K.; Hsieh, Y.S.; Yang, J.S.; Zavras, A.I.; Hsieh, Y.H.; Su, S.C.; Kao, T.Y.; Chen, P.N.; Chu, S.C. Antimetastatic effects of terminalia catappa l. On oral cancer via a down-regulation of metastasis-associated proteases. Food Chem. Toxicol. 2010, 48, 1052–1058. [Google Scholar] [CrossRef]
- Chang, L.; Karin, M. Mammalian map kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the mapk–ras–raf signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef]
- Reddy, K.B.; Nabha, S.M.; Atanaskova, N. Role of map kinase in tumor progression and invasion. Cancer Metastasis Rev. 2003, 22, 395–403. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.-J. Pathological roles of mapk signaling pathways in human diseases. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.O.; Park, S.J.; Yun, C.H.; Chung, A.S. Roles of matrix metalloproteinases in tumor metastasis and angiogenesis. J. Biochem. Mol. Biol. 2003, 36, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Huang, S. Role of mtor signaling in tumor cell motility, invasion and metastasis. Curr. Protein Pept. Sci. 2011, 12, 30–42. [Google Scholar] [PubMed]
- Kumar, A.; El-Osta, A.; Hussain, A.A.; Marshall, J. Increased sequestration of matrix metalloproteinases in ageing human bruch’s membrane: Implications for ecm turnover. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2664–2670. [Google Scholar] [CrossRef] [PubMed]
- Bauvois, B. New facets of matrix metalloproteinases mmp-2 and mmp-9 as cell surface transducers: Outside-in signaling and relationship to tumor progression. Biochim. Biophys. Acta 2012, 1825, 29–36. [Google Scholar] [CrossRef]
- Lustosa, S.A.; Viana Lde, S.; Affonso, R.J., Jr.; Silva, S.R.; Denadai, M.V.; de Toledo, S.R.; Oliveira, I.D.; Matos, D. Expression profiling using a cdna array and immunohistochemistry for the extracellular matrix genes fn-1, itga-3, itgb-5, mmp-2, and mmp-9 in colorectal carcinoma progression and dissemination. Sci. World J. 2014, 2014, 102541. [Google Scholar] [CrossRef]
- Mason, S.D.; Joyce, J.A. Proteolytic networks in cancer. Trends Cell Biol. 2011, 21, 228–237. [Google Scholar] [CrossRef]
- Hildenbrand, R.; Allgayer, H.; Marx, A.; Stroebel, P. Modulators of the urokinase-type plasminogen activation system for cancer. Expert Opin. Investig. Drugs 2010, 19, 641–652. [Google Scholar] [CrossRef]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Basu, B.; Correa de Sampaio, P.; Mohammed, H.; Fogarasi, M.; Corrie, P.; Watkins, N.A.; Smethurst, P.A.; English, W.R.; Ouwehand, W.H.; Murphy, G. Inhibition of mt1-mmp activity using functional antibody fragments selected against its hemopexin domain. Int. J. Biochem. Cell Biol. 2012, 44, 393–403. [Google Scholar] [CrossRef]
- Parsons, J.T. Focal adhesion kinase: The first ten years. J. Cell Sci. 2003, 116, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.W.; Ruefli, A.A.; Lowe, S.W. Apoptosis: A link between cancer genetics and chemotherapy. Cell 2002, 108, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-kinase–akt pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Oshimura, M.A.; Ito, H. Pi3k-akt pathway: Its functions and alterations in human cancer. Apoptosis 2004, 9, 667–676. [Google Scholar] [CrossRef]
- Gupta, A.K.; Soto, D.E.; Feldman, M.D.; Goldsmith, J.D.; Mick, R.; Hahn, S.M.; Machtay, M.; Muschel, R.J.; McKenna, W.G. Signaling pathways in nsclc as a predictor of outcome and response to therapy. Lung 2004, 182, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.-W.; Chen, P.-S.; Wu, C.-H.; Jeng, Y.-F.; Wang, C.-J. A-chaconine-reduced metastasis involves a pi3k/akt signaling pathway with downregulation of nf-κb in human lung adenocarcinoma a549 cells. J. Agric. Food Chem. 2007, 55, 11035–11043. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, W.; Zhang, X.; Lin, S.; Chen, Z. Overexpression of kiss-1 reduces colorectal cancer cell invasion by downregulating mmp-9 via blocking pi3k/akt/nf-κb signal pathway. Int. J. Oncol. 2016, 48, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Lin, X.; Ding, L.; Zeng, Y.; Pang, D.; Ouyang, N.; Xiang, Y.; Yao, H. Arhgap 42 promotes cell migration and invasion involving pi 3k/akt signaling pathway in nasopharyngeal carcinoma. Cancer Med. 2018, 7, 3862–3874. [Google Scholar] [CrossRef]
- Fujita, M.; Imadome, K.; Endo, S.; Shoji, Y.; Yamada, S.; Imai, T. Nitric oxide increases the invasion of pancreatic cancer cells via activation of the pi3k–akt and rhoa pathways after carbon ion irradiation. FEBS Lett. 2014, 588, 3240–3250. [Google Scholar] [CrossRef] [Green Version]
- Badgwell, D.B.; Lu, Z.; Le, K.; Gao, F.; Yang, M.; Suh, G.K.; Bao, J.-J.; Das, P.; Andreeff, M.; Chen, W. The tumor-suppressor gene arhi (diras3) suppresses ovarian cancer cell migration through inhibition of the stat3 and fak/rho signaling pathways. Oncogene 2012, 31, 68–79. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yan, J.; Xu, H.; Yang, Y.; Li, W.; Wu, H.; Liu, C. Extremely low frequency electromagnetic fields promote mesenchymal stem cell migration by increasing intracellular ca 2+ and activating the fak/rho gtpases signaling pathways in vitro. Stem Cell. Res. Ther. 2018, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Bandaru, P.; Kondo, Y.; Kuriyan, J. The interdependent activation of son-of-sevenless and ras. Cold Spring Harb. Perspect. Med. 2019, 9, a031534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giubellino, A.; Burke, T.R.; Bottaro, D.P. Grb2 signaling in cell motility and cancer. Expert Opin. Ther. Targets 2008, 12, 1021–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, P.P.; Blenis, J. Erk and p38 mapk-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebolt-Leopold, J.S. Development of anticancer drugs targeting the map kinase pathway. Oncogene 2000, 19, 6594–6599. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Shi, S.; Ma, Y.G.; Fan, F.; Yao, Z.Z. Lysophosphatidic acid enhances human hepatocellular carcinoma cell migration, invasion and adhesion through p38 mapk pathway. Hepato-Gastroenterol. 2012, 59, 785–789. [Google Scholar]
- Tan, F.L.; Ooi, A.; Huang, D.; Wong, J.C.; Qian, C.N.; Chao, C.; Ooi, L.; Tan, Y.M.; Chung, A.; Cheow, P.C.; et al. P38delta/mapk13 as a diagnostic marker for cholangiocarcinoma and its involvement in cell motility and invasion. Int. J. Cancer 2010, 126, 2353–2361. [Google Scholar] [CrossRef]
- Tsai, J.P.; Hsiao, P.C.; Yang, S.F.; Hsieh, S.C.; Bau, D.T.; Ling, C.L.; Pai, C.L.; Hsieh, Y.H. Licochalcone a suppresses migration and invasion of human hepatocellular carcinoma cells through downregulation of mkk4/jnk via nf-kappab mediated urokinase plasminogen activator expression. PLoS ONE 2014, 9, e86537. [Google Scholar]
- Zucchini-Pascal, N.; Peyre, L.; Rahmani, R. Crosstalk between beta-catenin and snail in the induction of epithelial to mesenchymal transition in hepatocarcinoma: Role of the erk1/2 pathway. Int. J. Mol. Sci. 2013, 14, 20768–20792. [Google Scholar] [CrossRef] [Green Version]
- Onder, T.T.; Gupta, P.B.; Mani, S.A.; Yang, J.; Lander, E.S.; Weinberg, R.A. Loss of e-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008, 68, 3645–3654. [Google Scholar] [CrossRef]
- Canel, M.; Serrels, A.; Frame, M.C.; Brunton, V.G. E-cadherin–integrin crosstalk in cancer invasion and metastasis. J. Cell Sci. 2013, 126, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalle Vedove, A.; Falchi, F.; Donini, S.; Dobric, A.; Germain, S.; Di Martino, G.P.; Prosdocimi, T.; Vettraino, C.; Torretta, A.; Cavalli, A. Structure-based virtual screening allows the identification of efficient modulators of e-cadherin-mediated cell–cell adhesion. Int. J. Mol. Sci. 2019, 20, 3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.-H.; Lee, H.; Chiang, T.-A.; Chen, M.-J.; Lin, S.-Y. Preventive effect of methanol extract from agrocybe cylindracea (meac) mushroom on the cooking oil fumes-induced expressions of cyclooxygenase-2 via suppression of nuclear factor-kappab (nf-κb) in cl-3 cells. J. Food Drug Anal. 2011, 19, 340–348. [Google Scholar]
- Chen, P.H.; Tu, H.P.; Wang, S.J.; Ko, A.M.S.; Lee, C.P.; Chiang, T.A.; Tsai, Y.S.; Lee, C.H.; Shieh, T.Y.; Ko, C.H. Monoamine oxidase a variants are associated with heavy betel quid use. Addict. Biol. 2012, 17, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-C.; Lai, K.-H.; Su, J.-H.; Wu, Y.-J.; Sheu, J.-H. 7-acetylsinumaximol b induces apoptosis and autophagy in human gastric carcinoma cells through mitochondria dysfunction and activation of the perk/eif2α/atf4/chop signaling pathway. Mar. Drugs 2018, 16, 104. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.-Y.; Wu, M.-L.; Chang, C.-I.; Liu, C.-I.; Cheng, T.-C.; Wu, Y.-J. Bornyl cis-4-hydroxycinnamate suppresses cell metastasis of melanoma through fak/pi3k/akt/mtor and mapk signaling pathways and inhibition of the epithelial-to-mesenchymal transition. Int. J. Mol. Sci. 2018, 19, 2152. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-J.; Lin, S.-H.; Din, Z.-H.; Su, J.-H.; Liu, C.-I. Sinulariolide Inhibits Gastric Cancer Cell Migration and Invasion through Downregulation of the EMT Process and Suppression of FAK/PI3K/AKT/mTOR and MAPKs Signaling Pathways. Mar. Drugs 2019, 17, 668. https://doi.org/10.3390/md17120668
Wu Y-J, Lin S-H, Din Z-H, Su J-H, Liu C-I. Sinulariolide Inhibits Gastric Cancer Cell Migration and Invasion through Downregulation of the EMT Process and Suppression of FAK/PI3K/AKT/mTOR and MAPKs Signaling Pathways. Marine Drugs. 2019; 17(12):668. https://doi.org/10.3390/md17120668
Chicago/Turabian StyleWu, Yu-Jen, Shih-Hsiung Lin, Zhong-Hao Din, Jui-Hsin Su, and Chih-I Liu. 2019. "Sinulariolide Inhibits Gastric Cancer Cell Migration and Invasion through Downregulation of the EMT Process and Suppression of FAK/PI3K/AKT/mTOR and MAPKs Signaling Pathways" Marine Drugs 17, no. 12: 668. https://doi.org/10.3390/md17120668
APA StyleWu, Y. -J., Lin, S. -H., Din, Z. -H., Su, J. -H., & Liu, C. -I. (2019). Sinulariolide Inhibits Gastric Cancer Cell Migration and Invasion through Downregulation of the EMT Process and Suppression of FAK/PI3K/AKT/mTOR and MAPKs Signaling Pathways. Marine Drugs, 17(12), 668. https://doi.org/10.3390/md17120668