Nutrient Deprivation-Associated Changes in Green Microalga Coelastrum sp. TISTR 9501RE Enhanced Potent Antioxidant Carotenoids

 and
and
Abstract
:1. Introduction
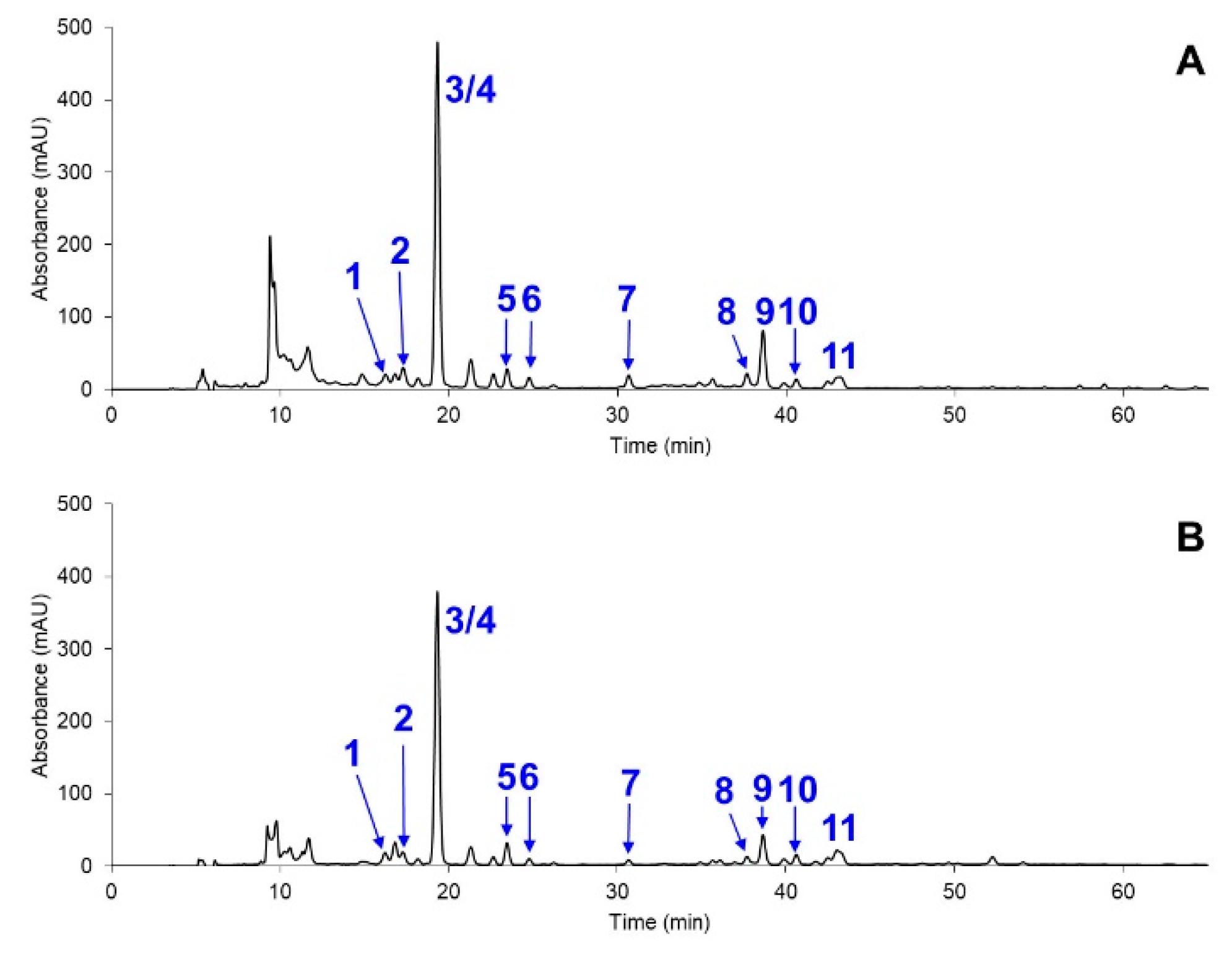
2. Results and Discussion
2.1. Morphology and Genetic Identification of the Microalga Coelastrum sp. TISTR 9501RE
2.2. Carotenoid Profiles of Coelastrum sp. TISTR 9501RE
3. Materials and Methods
3.1. Microalgal Strain and Culture Conditions
3.2. Molecular Identification and Polyphasic Taxonomy Approaches
3.3. Carotenoids Extraction
- The suspension was vortexed for 5 min, followed by 2 min centrifugation at 10,000 rpm. The supernatant was collected, and 1 mL of fresh acetone was added to the crude precipitate. This process was repeated one more time.
- The suspension from step 1 was sonicated for 30 min, followed by 2 min centrifugation at 10,000 rpm. Then, the supernatant was collected, and 1 mL of fresh acetone was added to the crude precipitate.
- Step 1 was repeated exactly as shown above.
- Step 2 was repeated exactly as shown above.
- Step 1 was repeated exactly as shown above.
3.4. Determination of the Overall Chlorophyll Content
3.5. HPLC and LC-MS Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Maresca, J.A.; Graham, J.E.; Bryant, D.A. The biochemical basis for structural diversity in the carotenoids of chlorophototrophic bacteria. Photosynth. Res. 2008, 97, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Avalos, J.; Carmen Limón, M. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef] [PubMed]
- Adadi, P.; Barakova, N.V.; Krivoshapkina, E.F. Selected methods of extracting carotenoids, characterization, and health concerns: A review. J. Agric. Food Chem. 2018, 66, 5925–5947. [Google Scholar] [CrossRef]
- Polívka, T.; Frank, H.A. Molecular factors controlling photosynthetic light harvesting by carotenoids. Acc. Chem. Res. 2010, 43, 1125–1134. [Google Scholar] [CrossRef]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Functional properties of carotenoids originating from algae. J. Sci. Food Agric. 2013, 93, 5–11. [Google Scholar] [CrossRef]
- Wang, B.; Zarka, A.; Trebst, A.; Boussiba, S. Astaxanthin accumulation in Haematococcus pluvialis (chlorophyceae) as an active photoprotective process under high irradiance. J. Phycol. 2003, 39, 1116–1124. [Google Scholar] [CrossRef]
- Cazzonelli, C.I. Carotenoids in nature: Insights from plants and beyond. Funct. Plant Biol. 2011, 38, 833–847. [Google Scholar] [CrossRef]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Breithaupt, D.E. Modern application of xanthophylls in animal feeding—A review. Trends Food Sci. Technol. 2007, 18, 501–506. [Google Scholar] [CrossRef]
- Vílchez, C.; Forján, E.; Cuaresma, M.; Bédmar, F.; Garbayo, I.; Vega, J.M. Marine carotenoids: Biological functions and commercial applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.-S. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef]
- Naguib, Y.M.A. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Higuera-Ciapara, I.; Félix-Valenzuela, L.; Goycoolea, F.M. Astaxanthin: A review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 2006, 46, 185–196. [Google Scholar] [CrossRef]
- Yuan, J.-P.; Peng, J.; Yin, K.; Wang, J.-H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.; Phang, S.-M.; Ravi, S.; Aswathanarayana, R. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Artaria, C. Astaxanthin in cardiovascular health and disease: Mechanisms of action, therapeutic merits, and knowledge gaps. Food Funct. 2017, 8, 39–63. [Google Scholar] [CrossRef]
- Davinelli, S.; Nielsen, M.; Scapagnini, G. Astaxanthin in skin health, repair, and disease: A comprehensive review. Nutrients 2018, 10, 522. [Google Scholar] [CrossRef]
- Viera, I.; Pérez-Gálvez, A.; Roca, M. Bioaccessibility of marine carotenoids. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Torrissen, O.J. Pigmentation of salmonids—Effect of carotenoids in eggs and start-feeding diet on survival and growth rate. Aquaculture 1984, 43, 185–193. [Google Scholar] [CrossRef]
- Surai, A.P.; Surai, P.F.; Steinberg, W.; Wakeman, W.G.; Speake, B.K.; Sparks, N.H.C. Effect of canthaxanthin content of the maternal diet on the antioxidant system of the developing chick. Br. Poult. Sci. 2003, 44, 612–619. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Rimbach, G. Canthaxanthin: From molecule to function. Mol. Nutr. Food Res. 2017, 61, 1600469. [Google Scholar] [CrossRef]
- Ma, L.; Lin, X.-M. Effects of lutein and zeaxanthin on aspects of eye health. J. Sci. Food Agric. 2010, 90, 2–12. [Google Scholar] [CrossRef]
- Koushan, K.; Rusovici, R.; Li, W.; Ferguson, L.; Chalam, K. The role of lutein in eye-related disease. Nutrients 2013, 5, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Manayi, A.; Abdollahi, M.; Raman, T.; Nabavi, S.F.; Habtemariam, S.; Daglia, M.; Nabavi, S.M. Lutein and cataract: From bench to bedside. Crit. Rev. Biotechnol. 2016, 36, 829–839. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Varela, J.C.; Pereira, H.; Vila, M.; León, R. Production of carotenoids by microalgae: Achievements and challenges. Photosynth. Res. 2015, 125, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-M.; Ren, L.-J.; Zhao, Q.-Y.; Ji, X.-J.; Huang, H. Microalgae for the production of lipid and carotenoids: A review with focus on stress regulation and adaptation. Biotechnol. Biofuels 2018, 11, 272. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced high-yield production of zeaxanthin, lutein, and β-carotene by a mutant of Chlorella zofingiensis. J. Agric. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Patrick, L. Beta carotene: The controversy continues. Altern. Med. Rev. 2000, 5, 530–545. [Google Scholar] [PubMed]
- Lin, J.-H.; Lee, D.-J.; Chang, J.-S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Sowbhagya, H.B.; Sampathu, S.R.; Krishnamurthy, N. Natural colorant from marigold-chemistry and technology. Food Rev. Int. 2004, 20, 33–50. [Google Scholar] [CrossRef]
- Rodríguez-Sáiz, M.; de la Fuente, J.L.; Barredo, J.L. Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Appl. Microbiol. Biotechnol. 2010, 88, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, D.; Niu, J.; Shen, S.; Wang, G. An economic assessment of astaxanthin production by large scale cultivation of Haematococcus pluvialis. Biotechnol. Adv. 2011, 29, 568–574. [Google Scholar] [CrossRef]
- Hejazi, M.A.; Holwerda, E.; Wijffels, R.H. Milking microalga Dunaliella salina for β-carotene production in two-phase bioreactors. Biotechnol. Bioeng. 2004, 85, 475–481. [Google Scholar] [CrossRef]
- Kim, S.-H.; Liu, K.-H.; Lee, S.-Y.; Hong, S.-J.; Cho, B.-K.; Lee, H.; Lee, C.-G.; Choi, H.-K. Effects of light intensity and nitrogen starvation on glycerolipid, glycerophospholipid, and carotenoid composition in Dunaliella tertiolecta culture. PLoS ONE 2013, 8, e72415. [Google Scholar] [CrossRef] [PubMed]
- Minyuk, G.; Chelebieva, E.; Chubchikova, I.; Dantsyuk, N.; Drobetskaya, I.; Sakhon, E.; Chekanov, K.; Solovchenko, A. Stress-induced secondary carotenogenesis in Coelastrella rubescens (Scenedesmaceae, Chlorophyta), a producer of value-added keto-carotenoids. Algae 2017, 32, 245–259. [Google Scholar] [CrossRef]
- Zhang, P.; Li, Z.; Lu, L.; Xiao, Y.; Liu, J.; Guo, J.; Fang, F. Effects of stepwise nitrogen depletion on carotenoid content, fluorescence parameters and the cellular stoichiometry of Chlorella vulgaris. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 181, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.J.; Lin, S.; Xu, W.; Cheung, P.C.K. Enhancement of the production of bioactive microalgal metabolites by ultraviolet radiation (UVA 365 nm). J. Agric. Food Chem. 2018, 66, 10215–10224. [Google Scholar] [CrossRef]
- Janchot, K.; Rauytanapanit, M.; Honda, M.; Hibino, T.; Sirisattha, S.; Praneenararat, T.; Kageyama, H.; Waditee-Sirisattha, R. Effects of potassium chloride-induced stress on the carotenoids canthaxanthin, astaxanthin, and lipid accumulations in the green Chlorococcal microalga strain TISTR 9500. J. Eukaryot. Microbiol. 2019. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, C.; Hou, Y.; Chen, S.; Xiao, D.; Zhang, J.; Chen, F. Isolation and characterization of a marine microalga for biofuel production with astaxanthin as a co-product. Energies 2013, 6, 2759–2772. [Google Scholar] [CrossRef]
- Úbeda, B.; Gálvez, J.Á.; Michel, M.; Bartual, A. Microalgae cultivation in urban wastewater: Coelastrum cf. pseudomicroporum as a novel carotenoid source and a potential microalgae harvesting tool. Bioresour. Technol. 2017, 228, 210–217. [Google Scholar]
- Soares, A.T.; da Costa, D.C.; Vieira, A.A.H.; Antoniosi Filho, N.R. Analysis of major carotenoids and fatty acid composition of freshwater microalgae. Heliyon 2019, 5, e01529. [Google Scholar] [CrossRef] [Green Version]
- Štenclová, L.; Fučíková, K.; Kaštovský, J.; Pažoutová, M. Molecular and morphological delimitation and generic classification of the family Oocystaceae (Trebouxiophyceae, Chlorophyta). J. Phycol. 2017, 53, 1263–1282. [Google Scholar] [CrossRef]
- Deli, J.; Gonda, S.; Nagy, L.Z.; Szabó, I.; Gulyás-Fekete, G.; Agócs, A.; Marton, K.; Vasas, G. Carotenoid composition of three bloom-forming algae species. Food Res. Int. 2014, 65, 215–223. [Google Scholar] [CrossRef]
- Han, D.; Li, Y.; Hu, Q. Astaxanthin in microalgae: Pathways, functions and biotechnological implications. Algae 2013, 28, 131–147. [Google Scholar] [CrossRef]
- Jin, H.; Lao, Y.M.; Zhou, J.; Zhang, H.J.; Cai, Z.H. Simultaneous determination of 13 carotenoids by a simple C18 column-based ultra-high-pressure liquid chromatography method for carotenoid profiling in the astaxanthin-accumulating Haematococcus pluvialis. J. Chromatogr. A 2017, 1488, 93–103. [Google Scholar] [CrossRef]
- Masojídek, J.; Torzillo, G. Mass cultivation of freshwater microalgae. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 2226–2235. [Google Scholar]
- Liu, J.; Sun, Z.; Gerken, H.; Liu, Z.; Jiang, Y.; Chen, F. Chlorella zofingiensis as an alternative microalgal producer of astaxanthin: Biology and industrial potential. Mar. Drugs 2014, 12, 3487–3515. [Google Scholar] [CrossRef] [PubMed]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Sashidhar, B.; Adholeya, A. The isolation and identification of new microalgal strains producing oil and carotenoid simultaneously with biofuel potential. Bioresour. Technol. 2016, 211, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-W.; Chuang, L.-T.; Yu, P.-C.; Chen, C.-N.N. Pigment production by a new thermotolerant microalga Coelastrella sp. F50. Food Chem. 2013, 138, 2071–2078. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Ritchie, R.J. Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents. Photosynthetica 2008, 46, 115–126. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Identity * | Retention Time ** | Proposed Formula | Precursor Mass | Found at Mass | Mass Error (ppm) |
|---|---|---|---|---|---|---|
| 1 | All-trans-astaxanthin | 16.24 | C40H52O4 | 597.3938 | 597.3936 | −0.48 |
| 2 | Violaxanthin isomer | 17.28 | C40H56O4 | 601.4251 | 601.4234 | −2.91 |
| 3 | All-trans-lutein | 19.32 | C40H56O2 | 569.4353 | 569.4317 | −6.34 |
| 4 | Chlorophyll b | 19.32 | C55H70N4O6Mg | 907.5219 | 907.5214 | −0.45 |
| 5 | All-trans-Canthaxanthin | 23.45 | C40H52O2 | 565.4040 | 565.4030 | −1.75 |
| 6 | Chlorophyll a | 25.65 | C55H72N4O5Mg | 893.5426 | 893.5393 | −3.66 |
| 7 | AME C18:4 isomer | 30.66 | C58H78O5 | 855.5922 | 855.5877 | −5.27 |
| 8 | AME C18:1 isomer | 37.67 | C58H84O5 | 861.6392 | 861.6346 | −5.29 |
| 9 | Chlorophyll b epimer | 38.62 | C55H70N4O6Mg | 907.5219 | 907.5294 | 8.32 |
| 10 | AME C18:2 isomer | 41.76 | C58H82O5 | 859.6235 | 859.6211 | −2.85 |
| 11 | Chlorophyll a epimer | 42.96 | C55H72N4O5Mg | 893.5426 | 893.5474 | 5.43 |
| Entry | Identity * | Retention Time ** | Proposed Formula | Precursor Mass | Found at Mass | Mass Error (ppm) |
|---|---|---|---|---|---|---|
| 1 | All-trans-astaxanthin | 16.22 | C40H52O4 | 597.3938 | 597.3935 | −0.53 |
| 2 | Violaxanthin isomer | 17.28 | C40H56O4 | 601.4251 | 601.4251 | −0.04 |
| 3 | All-trans-lutein | 19.33 | C40H56O2 | 569.4353 | 569.4327 | −4.54 |
| 4 | Chlorophyll b | 19.33 | C55H70N4O6Mg | 907.5219 | 907.5218 | −0.03 |
| 5 | All-trans-Canthaxanthin | 23.46 | C40H52O2 | 565.4040 | 565.4035 | −0.87 |
| 6 | Chlorophyll a | 25.65 | C55H72N4O5Mg | 893.5426 | 893.5425 | −0.12 |
| 7 | AME C18:4 isomer | 30.67 | C58H78O5 | 855.5922 | 855.5918 | −0.51 |
| 8 | AME C18:1 isomer | 37.69 | C58H84O5 | 861.6392 | 861.6392 | 0.02 |
| 9 | Chlorophyll b epimer | 38.64 | C55H70N4O6Mg | 907.5220 | 907.5329 | 12.2 |
| 10 | AME C18:2 isomer | 41.77 | C58H82O5 | 859.6235 | 859.6231 | −0.49 |
| 11 | Chlorophyll a epimer | 43.00 | C55H72N4O5Mg | 893.5426 | 893.5507 | 9.04 |
| Compound | Amount (mg/g DW) | ||
|---|---|---|---|
| Control Condition | Nutrient-Depleted Condition | Nutrient-Depleted Condition (20,000-L Pond) | |
| All-trans-astaxanthin | 0.03 ± 0.001 | 0.11 ± 0.01 | 0.18 ± 0.004 |
| All-trans-lutein | 2.35 ± 0.05 | 4.18 ± 0.46 | 3.13 ± 0.07 |
| All-trans-Canthaxanthin | 0.27 ± 0.03 | 1.15 ± 0.10 | 1.37 ± 0.03 |
| Entry | Identity | Retention Time * | Proposed Formula | Precursor Mass | Found at Mass | Mass Error (ppm) |
|---|---|---|---|---|---|---|
| 1 | All-trans-astaxanthin | 16.21 | C40H52O4 | 597.3938 | 597.3935 | −0.60 |
| 2 | All-trans-lutein | 19.32 | C40H56O2 | 569.4353 | 569.4351 | −0.39 |
| 3 | Chlorophyll b | 19.32 | C55H70N4O6Mg | 907.5219 | 907.5227 | 0.88 |
| 4 | All-trans-Canthaxanthin | 23.44 | C40H52O2 | 565.4040 | 565.4044 | 0.65 |
| 5 | Chlorophyll a | 25.64 | C55H72N4O5Mg | 893.5426 | 893.5434 | 0.95 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rauytanapanit, M.; Janchot, K.; Kusolkumbot, P.; Sirisattha, S.; Waditee-Sirisattha, R.; Praneenararat, T. Nutrient Deprivation-Associated Changes in Green Microalga Coelastrum sp. TISTR 9501RE Enhanced Potent Antioxidant Carotenoids. Mar. Drugs 2019, 17, 328. https://doi.org/10.3390/md17060328
Rauytanapanit M, Janchot K, Kusolkumbot P, Sirisattha S, Waditee-Sirisattha R, Praneenararat T. Nutrient Deprivation-Associated Changes in Green Microalga Coelastrum sp. TISTR 9501RE Enhanced Potent Antioxidant Carotenoids. Marine Drugs. 2019; 17(6):328. https://doi.org/10.3390/md17060328
Chicago/Turabian StyleRauytanapanit, Monrawat, Kantima Janchot, Pokchut Kusolkumbot, Sophon Sirisattha, Rungaroon Waditee-Sirisattha, and Thanit Praneenararat. 2019. "Nutrient Deprivation-Associated Changes in Green Microalga Coelastrum sp. TISTR 9501RE Enhanced Potent Antioxidant Carotenoids" Marine Drugs 17, no. 6: 328. https://doi.org/10.3390/md17060328
APA StyleRauytanapanit, M., Janchot, K., Kusolkumbot, P., Sirisattha, S., Waditee-Sirisattha, R., & Praneenararat, T. (2019). Nutrient Deprivation-Associated Changes in Green Microalga Coelastrum sp. TISTR 9501RE Enhanced Potent Antioxidant Carotenoids. Marine Drugs, 17(6), 328. https://doi.org/10.3390/md17060328