Astaxanthin Complexes to Attenuate Muscle Damage after In Vivo Femoral Ischemia-Reperfusion

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. P/D/CD-A Scaffold Loading and Antioxidant Activity Evaluation
2.2. Evaluation of In Vitro and In Vivo Toxicity of P/D/CD-A Scaffold
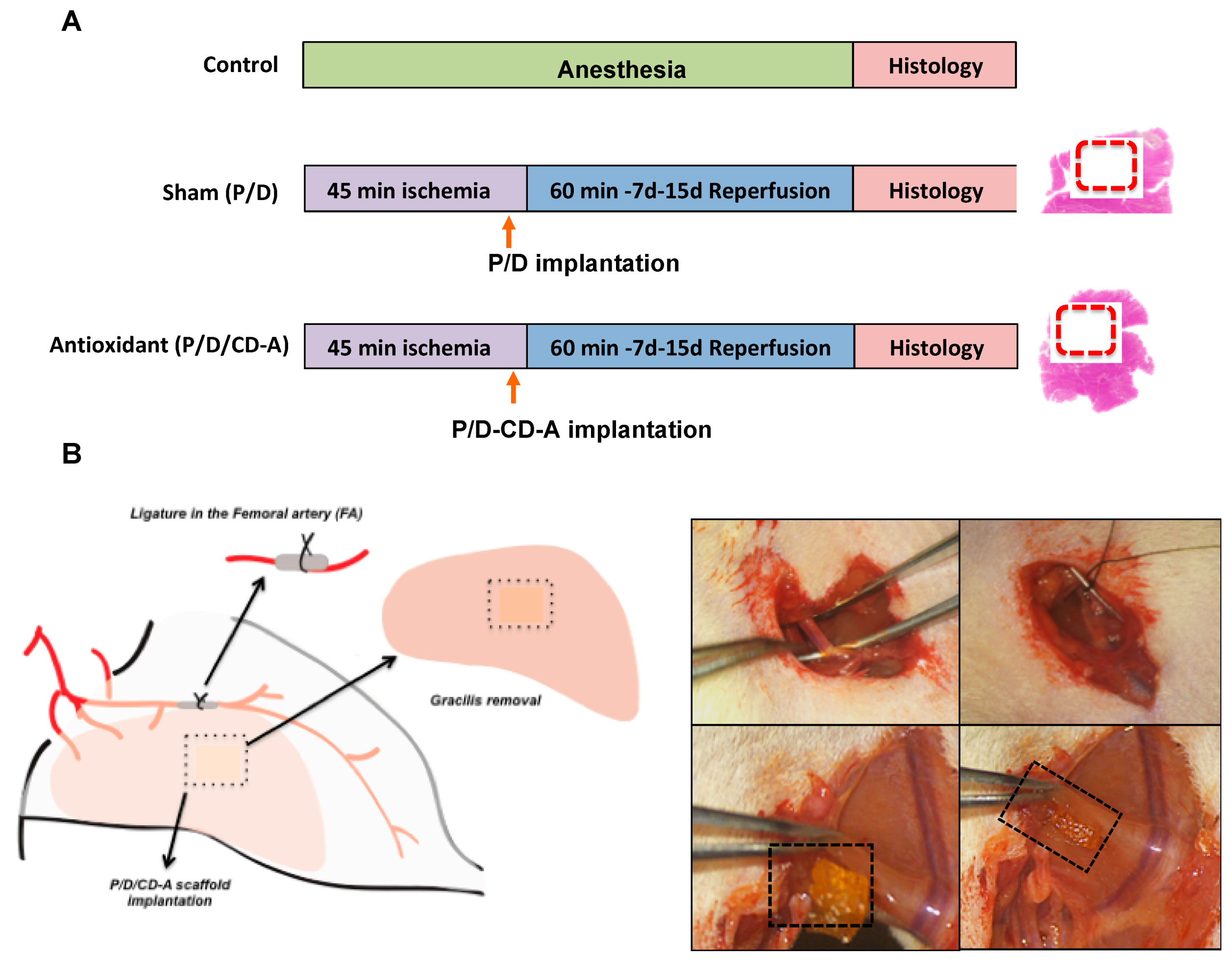
2.3. In Vivo Evaluation of P/D/CD-A Scaffolds in Femoral I/R Model
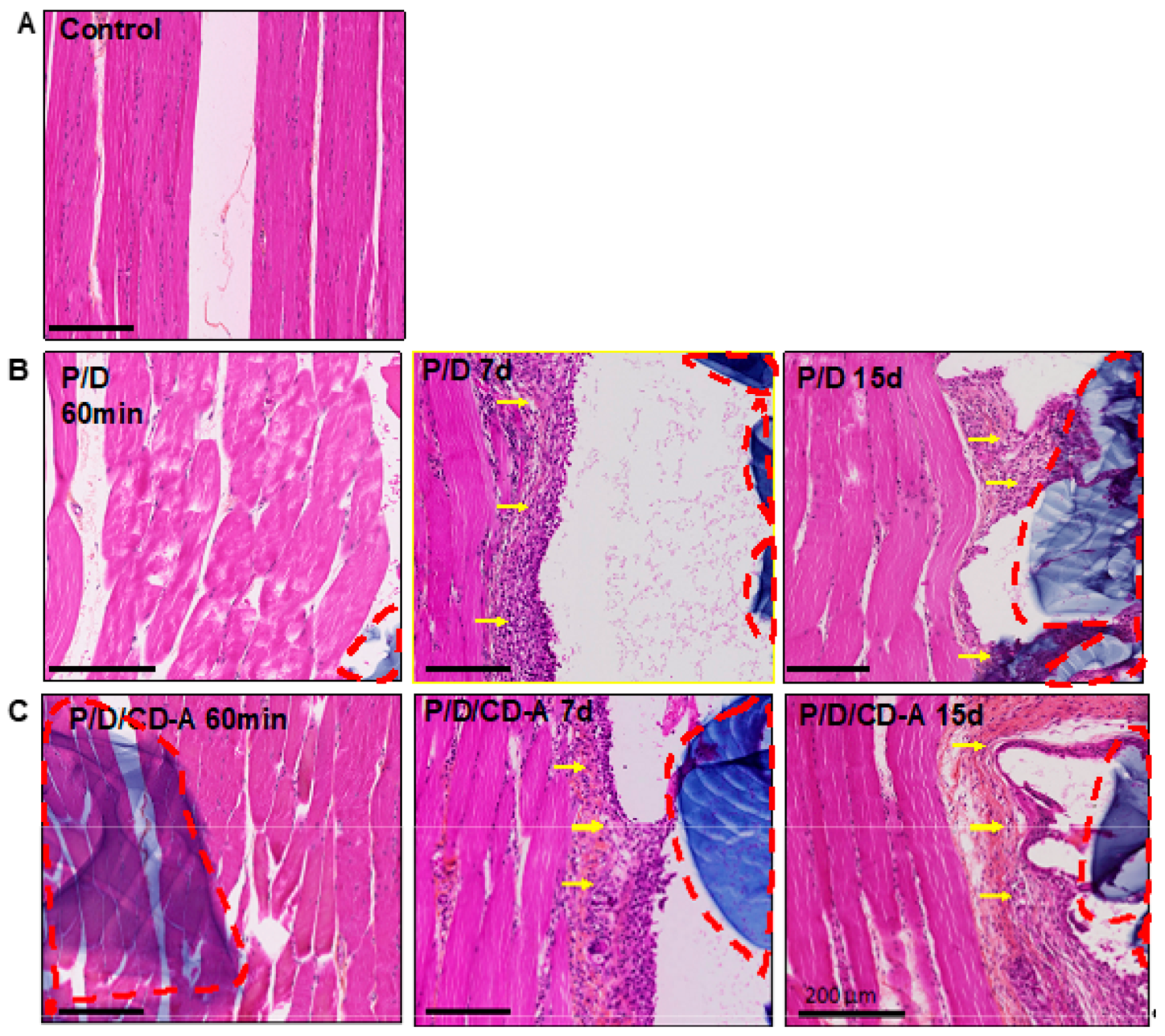
2.3.1. Histological and Immunohistological Analysis
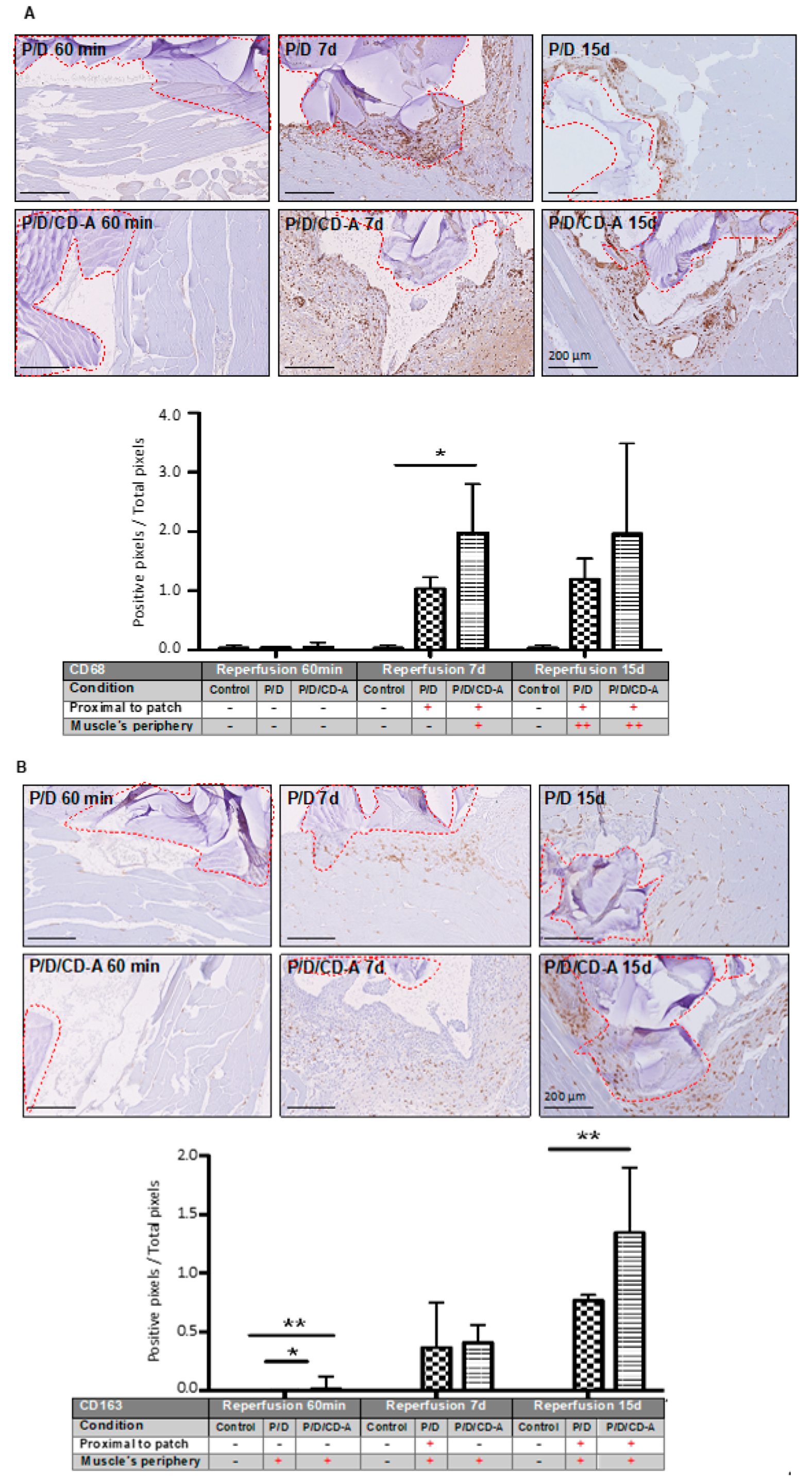
2.3.2. Phagocyte Responses to Scaffold
2.3.3. Nrf2/HO1/NQO1 Endogenous Antioxidant Systems
2.3.4. Detection of Oxidative Stress induced by Iron Overload in the Tissues
3. Discussion
4. Materials and Methods
4.1. Chemical and Biological Reagents
4.2. Pullulan/Dextran/CycloDextrin-Astaxanthin Scaffold Preparation and Loading
4.2.1. P/D Scaffold Preparation
4.2.2. CD-A Preparation
4.2.3. Scaffold Loading and Characterization
4.2.4. Release Kinetics Evaluation
4.2.5. CD-A Antioxidant Activity Evaluation
4.2.6. Cell Viability Assay
4.2.7. Surgical Procedure and Experimental Design
4.2.8. Histological and Immunological Evaluation
4.2.9. Statistical Analysis
4.2.10. List of abbreviations
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Madamanchi, N.R.; Vendrov, A.; Runge, M.S. Oxidative stress and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Collard, C.D.; Gelman, S. Prevention of Ischemia–Reperfusion Injury. Anesthesiology 2001, 94, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Yellon, D.M.; Hausenloy, D.J. Myocardial Reperfusion Injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M. Oxygen-derived free radicals in postischemic tissue injury. N. Engl. J. Med. 1985, 312, 159–163. [Google Scholar] [PubMed]
- Guillot, M.; Charles, A.L.; Chamaraux-Tran, T.N.; Bouitbir, J.; Meyer, A.; Zoll, J.; Schneider, F.; Geny, B. Oxidative stress precedes skeletal muscle mitochondrial dysfunction during experimental aortic cross-clamping but is not associated with early lung, heart, brain, liver, or kidney mitochondrial impairment. J. Vasc. Surg. 2014, 60, 1043–1051. [Google Scholar] [CrossRef]
- Blaisdell, W.F. The pathophysiology of skeletal muscle ischemia and the reperfusion syndrome: A review. Cardiovasc. Surg. 2002, 10, 620–630. [Google Scholar] [CrossRef]
- Gardner, A.W.; Parker, D.E.; Montgomery, P.S.; Sosnowska, D.; Casanegra, A.I.; Ungvari, Z.; Csiszar, A.; Sonntag, W.E. Endothelial Cell Inflammation and Antioxidant Capacity are Associated with Exercise Performance and Microcirculation in Patients with Symptomatic Peripheral Artery Disease. Angiology 2015, 66, 867–874. [Google Scholar] [CrossRef]
- Charles, A.L.; Guilbert, A.S.; Guillot, M.; Talha, S.; Lejay, A.; Meyer, A.; Kindo, M.; Wolff, V.; Bouitbir, J.; Zoll, J.; et al. Muscles susceptibility to ischemia-reperfusion injuries depends on fiber type specific antioxidant level. Front. Physiol. 2017, 8, 52. [Google Scholar] [CrossRef]
- Ranga Rao, A.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. In vivo bioavailability and antioxidant activity of carotenoids from microalgal biomass—A repeated dose study. Food Res. Int. 2013, 54, 711–717. [Google Scholar] [CrossRef]
- Pashkow, F.J.; Watumull, D.G.; Campbell, C.L. Astaxanthin: A novel potential treatment for oxidative stress and inflammation in cardiovascular disease. Am. J. Cardiol. 2008, 101, 58D–68D. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Fu, K.; Zhao, X.; Zhang, Y.; Yuan, Y.; Zhang, S.; Gu, X.; Guo, H. Protective effects of astaxanthin against ischemia/reperfusion induced renal injury in mice. J. Transl. Med. 2015, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, F.; Xia, Y.; Dai, W.; Chen, K.; Li, S.; Liu, T.; Zheng, Y.; Wang, J.; Lu, W.; et al. Astaxanthin pretreatment attenuates hepatic ischemia reperfusion-induced apoptosis and autophagy via the ROS/MAPK pathway in mice. Mar. Drugs 2015, 13, 3368–3387. [Google Scholar] [CrossRef] [PubMed]
- Adluri, R.S.; Thirunavukkarasu, M.; Zhan, L.; Maulik, N.; Svennevig, K.; Bagchi, M.; Maulik, G. Cardioprotective Efficacy of a Novel Antioxidant Mix VitaePro Against Ex Vivo Myocardial Ischemia-Reperfusion Injury. Cell Biochem. Biophys. 2013, 67, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Kuo, C.-C.; Chou, J.; Delvolve, A.; Jackson, S.N.; Post, J.; Woods, A.S.; Hoffer, B.J.; Wang, Y.; Harvey, B.K. Astaxanthin reduces ischemic brain injury in adult rats. FASEB J. 2009, 23, 1958–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombes, J.S.; Sharman, J.E.; Fassett, R.G. Astaxanthin has no effect on arterial stiffness, oxidative stress, or inflammation in renal transplant recipients: A randomized controlled trial (the XANTHIN trial). Am. J. Clin. Nutr. 2016, 103, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, M.; Barzegari, A.; Letourneur, D.; Gueguen, V.; Pavon-Djavid, G. Oxidative stress regulation on endothelial cells by hydrophilic astaxanthin complex: Chemical, biological and molecular antioxidant activity evaluation. Oxid. Med. Cell. Longev. 2017, 2017, 8073798. [Google Scholar] [CrossRef] [PubMed]
- Abed, A.; Assoul, N.; Ba, M.; Derkaoui, S.M.; Portes, P.; Louedec, L.; Flaud, P.; Bataille, I.; Letourneur, D.; Meddahi-Pellé, A.; et al. Influence of polysaccharide composition on the biocompatibility of pullulan/dextran-based hydrogels. J. Biomed. Mater. Res. Part A 2011, 96, 535–542. [Google Scholar] [CrossRef]
- Derkaoui, S.M.; Avramoglou, T.; Barbaud, C.; Letourneur, D. Synthesis and Characterization of a New Polysaccharide-graft-polymethacrylate Copolymer for Three-Dimensional Hybrid Hydrogels. Biomacromolecules 2008, 9, 3033–3038. [Google Scholar] [CrossRef]
- Chaouat, M.; Le Visage, C.; Autissier, A.; Chaubet, F.; Letourneur, D. The evaluation of a small-diameter polysaccharide-based arterial graft in rats. Biomaterials 2006, 27, 5546–5553. [Google Scholar] [CrossRef]
- Purnama, A.; Aid-Launais, R.; Haddad, O.; Maire, M.; Mantovani, D.; Letourneur, D.; Hlawaty, H.; Le Visage, C. Fucoidan in a 3D scaffold interacts with vascular endothelial growth factor and promotes neovascularization in mice. Drug Deliv. Transl. Res. 2013, 5, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.T.; Wolfson, S.K.; Kuller, L.H. Lower extremity arterial disease and the aging process: A review. J. Clin. Epidemiol. 1992, 45, 529–542. [Google Scholar] [CrossRef]
- Biswas, C.; Shah, N.; Muthu, M.; La, P.; Fernando, A.P.; Sengupta, S.; Yang, G.; Dennery, P.A. Nuclear heme oxygenase-1 (HO-1) modulates subcellular distribution and activation of Nrf2, impacting metabolic and anti-oxidant defenses. J. Biol. Chem. 2014, 289, 26882–26894. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Fukuyama, N.; Aki, A.; Kanabuchi, K.; Kimura, K.; Taira, H.; Tanaka, E.; Wakana, N.; Mori, H.; Inoue, H. Search for appropriate experimental methods to create stable hind-limb ischemia in mouse. Tokai J. Exp. Clin. Med. 2006, 31, 118–122. [Google Scholar]
- Carmo-Araújo, E.M.; Dal-Pai-Silva, M.; Dal-Pai, V.; Cecchini, R.; Anjos Ferreira, A.L. Ischaemia and reperfusion effects on skeletal muscle tissue: Morphological and histochemical studies. Int. J. Exp. Pathol. 2007, 88, 147–154. [Google Scholar] [CrossRef]
- Grisotto, P.C.; dos Santos, A.C.; Coutinho-Netto, J.; Cherri, J.; Piccinato, C.E. Indicators of oxidative injury and alterations of the cell membrane in the skeletal muscle of rats submitted to ischemia and reperfusion. J. Surg. Res. 2000, 92, 1–6. [Google Scholar] [CrossRef]
- Harman, J.W. A histological study of skeletal muscle in acute ischemia. Am. J. Pathol. 1946, XXIII, 551–565. [Google Scholar]
- Sagara, G.; Ogikubo, O.; Takeyama, H.; Tada, T.; Suzuri, N.; Okamoto, H.; Otsuka, T. Comparison of effects of ischemia period in rat limb ischemia/reperfusion injury. Nagoya Med. J. 2009, 50, 83–91. [Google Scholar]
- Yoshihara, T.; Yamamoto, Y.; Shibaguchi, T.; Miyaji, N. Dietary astaxanthin supplementation attenuates disuse-induced muscle atrophy and myonuclear apoptosis in the rat soleus muscle. J. Physiol. Sci. 2016, 67, 181–190. [Google Scholar] [CrossRef]
- Santos, M.R.; Celotto, A.C.; Capellini, V.K.; Evora, P.R.; Piccinato, C.E.; Joviliano, E.E. The protective effect of cilostazol on isolated rabbit femoral arteries under conditions of ischemia and reperfusion: The role of the nitric oxide pathway. Clinics (Sao Paulo) 2012, 67, 171–178. [Google Scholar] [CrossRef]
- Huk, I.; Nanobashvili, J.; Neumayer, C.; Punz, A.; Mueller, M.; Afkhampour, K.; Mittlboeck, M.; Losert, U.; Polterauer, P.; Roth, E.; et al. L-arginine treatment alters the kinetics of nitric oxide and superoxide release and reduces ischemia/reperfusion injury in skeletal muscle. Circulation 1997, 96, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Koksal, C.; Bozkurt, A.K.; Cangel, U.; Ustundag, N.; Konukoǧlu, D.; Musellim, B.; Sayin, A.G. Attenuation of ischemia/reperfusion injury by N-acetylcysteine in a rat hind limb model. J. Surg. Res. 2003, 111, 236–239. [Google Scholar] [CrossRef]
- Koksal, C.; Bozkurt, A.K.; Sirin, G.; Konukoglu, D.; Ustundag, N. Aprotinin ameliorates ischemia/reperfusion injury in a rat hind limb model. Vasc. Pharmacol. 2004, 41, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.-Y.; Shan, W.-G.; Wang, S.-F.; Hu, M.-M.; Chen, Y. Effects of astaxanthin on blood coagulation, fibrinolysis and platelet aggregation in hyperlipidemic rats. Pharm. Biol. 2017, 55, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Gross, G.J.; Lockwood, S.F. Cardioprotection and myocardial salvage by a disodium disuccinate astaxanthin derivative (CardaxTM). Life Sci. 2004, 75, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhou, Y.; Li, X.F.; Wan, Q.J.; Yu, L.H. Preventive treatment of astaxanthin provides neuroprotection through suppression of reactive oxygen species and activation of antioxidant defense pathway after stroke in rats. Brain Res. Bull. 2017, 130, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Derkaoui, S.M.; Labbé, A.; Chevallier, P.; Holvoet, S.; Roques, C.; Avramoglou, T.; Mantovani, D.; Letourneur, D. A new dextran-graft-polybutylmethacrylate copolymer coated on 316L metallic stents enhances endothelial cell coverage. Acta Biomater. 2012, 8, 3509–3515. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.H.M.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage polarisation: An immunohistochemical approach for identifying M1 and M2 macrophages. PLoS ONE 2013, 8, e80908. [Google Scholar] [CrossRef]
- Klopfleisch, R. Macrophage reaction against biomaterials in the mouse model—Phenotypes, functions and markers. Acta Biomater. 2016, 43, 3–13. [Google Scholar] [CrossRef]
- Lau, S.K.; Chu, P.G.; Weiss, L.M. CD163A specific marker of macrophages in paraffin-embedded tissue samples. Am. J. Clin. Pathol. 2004, 122, 794–801. [Google Scholar] [CrossRef]
- Barbosa, N.; Sá, M.C.; Pinto, L.P.; Freitas, R.A. Immunohistochemical study of macrophages subpopulations associated with squamous cell carcinoma of the tongue, with and without metastasis. J. Bras. Patol. Med. Lab. 2015, 51, 415–421. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.M. Biological responses to materials. Annu. Rev. Mater. Res 2001, 31, 81–110. [Google Scholar] [CrossRef]
- Wokalek, H.; Ruh, H. Time course of wound healing. J. Biomater. Appl. 1991, 5, 337–362. [Google Scholar] [CrossRef] [PubMed]
- Racz, I.B.; Illyes, G.; Sarkadi, L.; Hamar, J. The functional and morphological damage of ischemic reperfused skeletal muscle. Eur. Surg. Res. 1997, 29, 254–263. [Google Scholar] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Badylak, S.F.; Valentin, J.E.; Ravindra, A.K.; McCabe, G.P.; Stewart-Akers, A.M. Macrophage phenotype as a determinant of biologic scaffold remodeling. Tissue Eng. Part A 2008, 14, 1835–1842. [Google Scholar] [CrossRef]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pan, H.; Wang, H.; Li, X.; Bu, X.; Wang, Q.; Gao, Y.; Wen, G.; Zhou, Y.; Cong, Z.; et al. Interplay between VEGF and Nrf2 regulates angiogenesis due to intracranial venous hypertension. Sci. Rep. 2016, 6, 37338. [Google Scholar] [CrossRef] [Green Version]
- Mann, G.E. Nrf2-mediated redox signalling in vascular health and disease. Free Radic. Biol. Med. 2014, 75, S1. [Google Scholar] [CrossRef]
- Shen, Y.; Liu, X.; Shi, J.; Wu, X. Involvement of Nrf2 in myocardial ischemia and reperfusion injury. Int. J. Biol. Macromol. 2019, 125, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, C.; Zachaki, S.; Alexoudi, A.; Chatzi, I.; Georgakakos, V.N.; Terzoudi, G.I.; Pantelias, G.E.; Karageorgiou, C.E.; Sambani, C. The C609T inborn polymorphism in NAD(P)H:quinone oxidoreductase 1 is associated with susceptibility to multiple sclerosis and affects the risk of development of the primary progressive form of the disease. Free Radic. Biol. Med. 2011, 51, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Latunde-Dada, G.O. Ferroptosis: Role of lipid peroxidation, iron and ferritinophagy. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 1893–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, J.; Kook, H.; Park, W.J. PICOT alleviates myocardial ischemia-reperfusion injury by reducing intracellular levels of reactive oxygen species. Biochem. Biophys. Res. Commun. 2017, 485, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Balla, G.; Jacob, H.S.; Eaton, J.W.; Belcher, J.D.; Vercellotti, G.M. Hemin: A possible physiological mediator of low density lipoprotein oxidation and endothelial injury. Arterioscler. Thromb. 1991, 11, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Nagy, E.; Eaton, J.W.; Jeney, V.; Soares, M.P.; Varga, Z.; Galajda, Z.; Szentmiklósi, J.; Méhes, G.; Csonka, T.; Smith, A.; et al. Red cells, hemoglobin, heme, iron and therogenesis. Aterioscler. Thromb. Vasc. Biol. 2010, 30, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Meguro, R.; Asano, Y.; Iwatsuki, H.; Shoumura, K. Perfusion-Perls and -Turnbull methods supplemented by DAB intensification for nonheme iron histochemistry: Demonstration of the superior sensitivity of the methods in the liver, spleen, and stomach of the rat. Histochem. Cell Biol. 2003, 120, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki, H.; Meguro, R.; Asano, Y.; Odagiri, S.; Li, C.; Shoumura, K. Chelatable Fe (II) is generated in the rat kidneys exposed to ischemia and reperfusion, and a divalent metal chelator, 2, 2′-dipyridyl, attenuates the acute ischemia/reperfusion-injury of the kidneys: A histochemical study by the perfusion-Perls and -Turn. Arch. Histol. Cytol. 2008, 71, 101–114. [Google Scholar] [CrossRef]
- Autissier, A.; Le Visage, C.; Pouzet, C.; Chaubet, F.; Letourneur, D. Fabrication of porous polysaccharide-based scaffolds using a combined freeze-drying/cross-linking process. Acta Biomater. 2010, 6, 3640–3648. [Google Scholar] [CrossRef]
- Sueishi, Y.; Ishikawa, M.; Yoshioka, D.; Endoh, N.; Oowada, S.; Shimmei, M.; Fujii, H.; Kotake, Y. Oxygen radical absorbance capacity (ORAC) of cyclodextrin-solubilized flavonoids, resveratrol and astaxanthin as measured with the ORAC-EPR method. J. Clin. Biochem. Nutr. 2012, 50, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Luyt, C.-E.; Meddahi-Pellé, A.; Ho-tin-noe, B.; Colliec-jouault, S.; Guezennec, J.; Louedec, L.; Prats, H.; Jacob, M.; Osborne-pellegrin, M.; Letourneur, D.; et al. Low-Molecular-Weight Fucoidan Promotes Therapeutic Revascularization in a Rat Model of Critical Hindlimb Ischemia. J. Pharmacol. Exp. Ther. 2003, 305, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzian, Z.; Gasser, T.C.; Blackwell, F.; Hyafil, F.; Louedec, L.; Deschildre, C.; Ghodbane, W.; Dorent, R.; Nicoletti, A.; Morvan, M.; et al. Peristrut microhemorrhages: A possible cause of in-stent neoatherosclerosis? Cardiovasc. Pathol. 2017, 26, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for valid histopathologic scoring in research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuluaga Tamayo, M.; Choudat, L.; Aid-Launais, R.; Thibaudeau, O.; Louedec, L.; Letourneur, D.; Gueguen, V.; Meddahi-Pellé, A.; Couvelard, A.; Pavon-Djavid, G. Astaxanthin Complexes to Attenuate Muscle Damage after In Vivo Femoral Ischemia-Reperfusion. Mar. Drugs 2019, 17, 354. https://doi.org/10.3390/md17060354
Zuluaga Tamayo M, Choudat L, Aid-Launais R, Thibaudeau O, Louedec L, Letourneur D, Gueguen V, Meddahi-Pellé A, Couvelard A, Pavon-Djavid G. Astaxanthin Complexes to Attenuate Muscle Damage after In Vivo Femoral Ischemia-Reperfusion. Marine Drugs. 2019; 17(6):354. https://doi.org/10.3390/md17060354
Chicago/Turabian StyleZuluaga Tamayo, Marisol, Laurence Choudat, Rachida Aid-Launais, Olivier Thibaudeau, Liliane Louedec, Didier Letourneur, Virginie Gueguen, Anne Meddahi-Pellé, Anne Couvelard, and Graciela Pavon-Djavid. 2019. "Astaxanthin Complexes to Attenuate Muscle Damage after In Vivo Femoral Ischemia-Reperfusion" Marine Drugs 17, no. 6: 354. https://doi.org/10.3390/md17060354
APA StyleZuluaga Tamayo, M., Choudat, L., Aid-Launais, R., Thibaudeau, O., Louedec, L., Letourneur, D., Gueguen, V., Meddahi-Pellé, A., Couvelard, A., & Pavon-Djavid, G. (2019). Astaxanthin Complexes to Attenuate Muscle Damage after In Vivo Femoral Ischemia-Reperfusion. Marine Drugs, 17(6), 354. https://doi.org/10.3390/md17060354