Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum

 ,
,  ,
,  , , ,
, , ,  ,
, 
 and
and 
Abstract
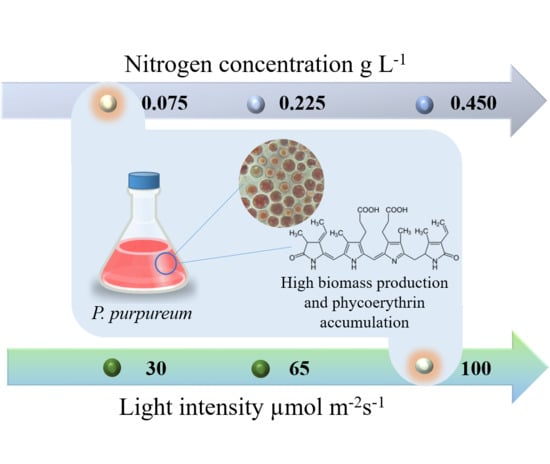
:
1. Introduction
2. Results
2.1. Experimental Results and Mathematical Model Fitness
2.2. Influence of Light Intensity and NaNO3 Concentration on the Production of P. purpureum Biomass
2.3. Influence of Light Intensity and NaNO3 Concentration on the Production of PE by P. purpureum
3. Discussion
3.1. Mathematical Model Parameter Analysis
3.2. Effect of Light Intensity on Biomass and PE Production in P. purpureum
3.3. Effect of Nitrogen Concentration on Biomass and Phycoerythrin from P. purpureum
4. Materials and Methods
4.1. Strain, Medium and Culture Conditions
4.2. Mathematical Model and Kinetic Parameters Calculation to Determine Growth Parameters
- , . Initial biomass () in the sample (mg L−1), biomass counts for each time.
- : Time (h).
- : Constant determined from the experimental data. Represents the maximum population of biomass counts when time increases indefinitely.
- : Constant determined from the experimental data.
- : Constant determined from the experimental data.
4.3. PE Extraction Methodology
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hirose, Y.; Misawa, N.; Yonekawa, C.; Nagao, N.; Watanabe, M.; Ikeuchi, M.; Eki, T. Characterization of the genuine type 2 chromatic acclimation in the two Geminocystis cyanobacteria. DNA Res. 2017, 24, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa-Hernández, J.E.; Romero-Castillo, K.D.; Parra-Arroyo, L.; Aguilar-Aguila-Isaías, M.A.; García-Reyes, I.E.; Ahmed, I.; Parra-Saldivar, R.; Bilal, M.; Iqbal, H. Mexican Microalgae Biodiversity and State-Of-The-Art Extraction Strategies to Meet Sustainable Circular Economy Challenges: High-Value Compounds and Their Applied Perspectives. Mar. Drugs 2019, 17, 174. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Nishizawa, M.; Shimizu, Y.; Saeki, H. Anti-inflammatory effects of dulse (Palmaria palmata) resulting from the simultaneous water-extraction of phycobiliproteins and chlorophyll a. Food Res. Int. 2017, 100, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ying, J.; Chang, Q.; Zhu, W.; Yang, G.; Xu, T.; Yi, H.; Pan, R.; Zhang, E.; Zeng, X.; et al. Effects of phycoerythrin from Gracilaria lemaneiformis in proliferation and apoptosis of SW480 cells. Oncol. Rep. 2016, 36, 3536–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.N.; Sonani, R.R.; Jakharia, K.; Bhastana, B.; Patel, H.M.; Chaubey, M.G.; Singh, N.K.; Madamwar, D. Antioxidant activity and associated structural attributes of Halomicronema phycoerythrin. Int. J. Biol. Macromol. 2018, 111, 359–369. [Google Scholar] [CrossRef]
- Al-Essa, M.K.; Melzer, S.; Tarnok, A.; Hadidi, K.A.; El-Khateeb, M. Fast RBC loading by fluorescent antibodies and nuclei staining dye and their potential bioanalytical applications. Z. Für Nat. C 2018, 73, 95–105. [Google Scholar] [CrossRef]
- Wu, J.; Chen, H.; Zhao, J.; Jiang, P. Fusion proteins of streptavidin and allophycocyanin alpha subunit for immunofluorescence assay. Biochem. Eng. J. 2017, 125, 97–103. [Google Scholar] [CrossRef]
- Chen, X.; Wu, M.; Yang, Q.; Wang, S. Preparation, characterization of food grade phycobiliproteins from Porphyra haitanensis and the application in liposome-meat system. LWT 2017, 77, 464–474. [Google Scholar] [CrossRef]
- Intrarapuk, A.; Awakairt, S.; Thammapalerd, N.; Mahannop, P.; Pattanawong, U.; Suppasiri, T. Comparison between R-phycocyanin-labeled and R-phycoerythrin-labeled monoclonal antibody (Mab) probes for the detection of Entamoeba histolytica trophozoites. Southeast Asian J. Trop. Med. Public Health 2001, 32, 165–171. [Google Scholar]
- Chandra, R.; Parra, R.; MN Iqbal, H. Phycobiliproteins: A novel green tool from marine origin blue-green algae and red algae. Protein Pept. Lett. 2017, 24, 118–125. [Google Scholar] [CrossRef]
- Koch, M.; Kniggendorf, A.K.; Meinhardt-Wollweber, M.; Roth, B. In vivo determination of carotenoid resonance excitation profiles of Chlorella vulgaris, Haematococcus pluvialis, and Porphyridium purpureum. J. Raman Spectrosc. 2018, 49, 404–411. [Google Scholar] [CrossRef]
- Rodrigues, R.D.P.; de Castro, F.C.; de Santiago-Aguiar, R.S.; Rocha, M.V.P. Ultrasound-assisted extraction of phycobiliproteins from Spirulina (Arthrospira) platensis using protic ionic liquids as solvent. Algal Res. 2018, 31, 454–462. [Google Scholar] [CrossRef]
- Kersen, P.; Paalme, T.; Pajusalu, L.; Martin, G. Biotechnological applications of the red alga Furcellaria lumbricalis and its cultivation potential in the Baltic Sea. Bot. Mar. 2017, 60, 207–218. [Google Scholar]
- Fuentes-Grünewald, C.; Bayliss, C.; Zanain, M.; Pooley, C.; Scolamacchia, M.; Silkina, A. Evaluation of batch and semi-continuous culture of Porphyridium purpureum in a photobioreactor in high latitudes using Fourier Transform Infrared spectroscopy for monitoring biomass composition and metabolites production. Bioresour. Technol. 2015, 189, 357–363. [Google Scholar] [CrossRef]
- Chang, J.; Le, K.; Song, X.; Jiao, K.; Zeng, X.; Ling, X.; Shi, T.; Tang, X.; Sun, Y.; Lin, L. Scale-up cultivation enhanced arachidonic acid accumulation by red microalgae Porphyridium purpureum. Bioprocess Biosyst. Eng. 2017, 40, 1763–1773. [Google Scholar]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef]
- Contreras-Angulo, J.R.; Mata, T.M.; Cuellar-Bermudez, S.P.; Caetano, N.S.; Chandra, R.; Garcia-Perez, J.S.; Muylaert, K.; Parra-Saldivar, R. Symbiotic Co-Culture of Scenedesmus sp. and Azospirillum brasilense on N-Deficient Media with Biomass Production for Biofuels. Sustainability 2019, 11, 707. [Google Scholar] [CrossRef]
- Fuentes-Grünewald, C.; Garcés, E.; Alacid, E.; Sampedro, N.; Rossi, S.; Camp, J. Improvement of lipid production in the marine strains Alexandrium minutum and Heterosigma akashiwo by utilizing abiotic parameters. J. Ind. Microbiol. Biotechnol. 2012, 39, 207–216. [Google Scholar]
- Heo, J.; Shin, D.S.; Cho, K.; Cho, D.H.; Lee, Y.J.; Kim, H.S. Indigenous microalga Parachlorella sp. JD-076 as a potential source for lutein production: Optimization of lutein productivity via regulation of light intensity and carbon source. Algal Res. 2018, 33, 1–7. [Google Scholar] [CrossRef]
- Sciandra, A.; Lazzara, L.; Claustre, H.; Babin, M. Responses of growth rate, pigment composition and optical properties of Cryptomonas sp. to light and nitrogen stresses. Mar. Ecol. Prog. Ser. 2000, 201, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Gigova, L.G.; Ivanova, N.J. Microalgae respond differently to nitrogen availability during culturing. J. Biosci. 2015, 40, 365–374. [Google Scholar] [CrossRef]
- Singh, P.; Guldhe, A.; Kumari, S.; Rawat, I.; Bux, F. Investigation of combined effect of nitrogen, phosphorus and iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surface methodology. Biochem. Eng. J. 2015, 94, 22–29. [Google Scholar] [CrossRef]
- Guihéneuf, F.; Stengel, D.B. Towards the biorefinery concept: Interaction of light, temperature and nitrogen for optimizing the co-production of high-value compounds in Porphyridium purpureum. Algal Res. 2015, 10, 152–163. [Google Scholar] [CrossRef]
- Kathiresan, S.; Sarada, R.; Bhattacharya, S.; Ravishankar, G.A. Culture media optimization for growth and phycoerythrin production from Porphyridium purpureum. Biotechnol. Bioeng. 2007, 96, 456–463. [Google Scholar] [CrossRef]
- Del Pilar Sánchez-Saavedra, M.; Castro-Ochoa, F.Y.; Nava-Ruiz, V.M.; Ruiz-Güereca, D.A.; Villagómez-Aranda, A.L.; Siqueiros-Vargas, F.; Molina-Cárdenas, C.A. Effects of nitrogen source and irradiance on Porphyridium cruentum. J. Appl. Phycol. 2018, 30, 783–792. [Google Scholar] [CrossRef]
- Baer, S.; Heining, M.; Schwerna, P.; Buchholz, R.; Hübner, H. Optimization of spectral light quality for growth and product formation in different microalgae using a continuous photobioreactor. Algal Res. 2016, 14, 109–115. [Google Scholar] [CrossRef]
- Coward, T.; Fuentes-Grünewald, C.; Silkina, A.; Oatley-Radcliffe, D.L.; Llewellyn, G.; Lovitt, R.W. Utilising light-emitting diodes of specific narrow wavelengths for the optimization and co-production of multiple high-value compounds in Porphyridium purpureum. Bioresour. Technol. 2016, 221, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Munier, M.; Jubeau, S.; Wijaya, A.; Morancais, M.; Dumay, J.; Marchal, L.; Jaouen, P.; Fleurence, J. Physicochemical factors affecting the stability of two pigments: R-phycoerythrin of Grateloupia turuturu and B-phycoerythrin of Porphyridium cruentum. Food Chem. 2014, 150, 400–407. [Google Scholar] [CrossRef]
- Darvehei, P.; Bahri, P.A.; Moheimani, N.R. Model development for the growth of microalgae: A review. Renew. Sustain. Energy Rev. 2018, 97, 233–258. [Google Scholar] [CrossRef]
- Fleck, P.; Posten, C. Modelling of growth and product formation of Porphyridium purpureum under defined conditions. IFAC Proc. Vol. 2004, 37, 281–286. [Google Scholar] [CrossRef]
- Fleck-Schneider, P.; Lehr, F.; Posten, C. Modelling of growth and product formation of Porphyridium purpureum. J. Biotechnol. 2007, 132, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.K.; Das, D. Improvement of carbon dioxide biofixation in a photobioreactor using Anabaena sp. PCC 7120. Process Biochem. 2013, 48, 1126–1132. [Google Scholar] [CrossRef]
- Zhen, G.; Lu, X.; Kobayashi, T.; Kumar, G.; Xu, K. Anaerobic co-digestion on improving methane production from mixed microalgae (Scenedesmus sp., Chlorella sp.) and food waste: Kinetic modeling and synergistic impact evaluation. Chem. Eng. J. 2016, 299, 332–341. [Google Scholar] [CrossRef]
- Dammak, M.; Hadrich, B.; Barkallah, M.; Hentati, F.; Hlima, H.B.; Pichon, C.; Denis, M.; Fendri, I.; Michaud, P.; Abdelkafi, S. Modelling Tetraselmis sp. growth-kinetics and optimizing bioactive-compound production through environmental conditions. Bioresour. Technol. 2018, 249, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Whiting, R.C.; Damert, W.C. When is simple good enough: A comparison of the Gompertz, Baranyi, and three-phase linear models for fitting bacterial growth curves. Food Microbiol. 1997, 14, 313–326. [Google Scholar] [CrossRef]
- Assunção, M.F.; Varejão, J.M.; Santos, L.M. Nutritional characterization of the microalga Ruttnera lamellosa compared to Porphyridium purpureum. Algal Res. 2017, 26, 8–14. [Google Scholar] [CrossRef]
- Kavitha, M.D.; Kathiresan, S.; Bhattacharya, S.; Sarada, R. Culture media optimization of Porphyridium purpureum: Production potential of biomass, total lipids, arachidonic and eicosapentaenoic acid. J. Food Sci. Technol. 2016, 53, 2270–2278. [Google Scholar] [CrossRef]
- Schulze, C.; Wetzel, M.; Reinhardt, J.; Schmidt, M.; Felten, L.; Mundt, S. Screening of microalgae for primary metabolites including β-glucans and the influence of nitrate starvation and irradiance on β-glucan production. J. Appl. Phycol. 2016, 28, 2719–2725. [Google Scholar] [CrossRef]
- Sandefur, H.N.; Asgharpour, M.; Mariott, J.; Gottberg, E.; Vaden, J.; Matlock, M.; Hestekin, J. Recovery of nutrients from swine wastewater using ultrafiltration: Applications for microalgae cultivation in photobioreactors. Ecol. Eng. 2016, 94, 75–81. [Google Scholar] [CrossRef]
- Gargouch, N.; Karkouch, I.; Elleuch, J.; Elkahoui, S.; Michaud, P.; Abdelkafi, S.; Laroche, C.; Fendri, I. Enhanced B-phycoerythrin production by the red microalga Porphyridium marinum: A powerful agent in industrial applications. Int. J. Biol. Macromol. 2018, 120, 2106–2114. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Wu, H.; Jiang, P.; Chen, Z.; Xiang, W. Growth and Biochemical Composition of Porphyridium purpureum SCS-02 under Different Nitrogen Concentrations. Mar. Drugs 2019, 17, 124. [Google Scholar] [CrossRef] [PubMed]
- Bazdar, E.; Roshandel, R.; Yaghmaei, S.; Mardanpour, M.M. The effect of different light intensities and light/dark regimes on the performance of photosynthetic microalgae microbial fuel cell. Bioresour. Technol. 2018, 261, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Qiang, H.; Richmond, A. Productivity and photosynthetic efficiency of Spirulina platensis as affected by light intensity, algal density and rate of mixing in a flat plate photobioreactor. J. Appl. Phycol. 1996, 8, 139–145. [Google Scholar] [CrossRef]
- Köst, H.P.; Senser, M.; Wanner, G. Effect of nitrate and sulphate starvation on Porphyridium cruentum cells. Z. Pflanzenphysiol. 1984, 113, 231–249. [Google Scholar] [CrossRef]
- Adda, M.; Merchuk, J.C.; Arad, S.M. Effect of nitrate on growth and production of cell-wall polysaccharide by the unicellular red alga Porphyridium. Biomass 1986, 10, 131–140. [Google Scholar] [CrossRef]
- Razaghi, A.; Godhe, A.; Albers, E. Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Open Life Sci. 2014, 9, 156–162. [Google Scholar] [CrossRef]
- Oh, S.H.; Han, J.G.; Kim, Y.; Ha, J.H.; Kim, S.S.; Jeong, M.H.; Jeong, H.S.; Kim, N.Y.; Cho, J.S.; Yoon, W.B.; et al. Lipid production in Porphyridium cruentum grown under different culture conditions. J. Biosci. Bioeng. 2009, 108, 429–434. [Google Scholar] [CrossRef]
- George, B.; Pancha, I.; Desai, C.; Chokshi, K.; Paliwal, C.; Ghosh, T.; Mishra, S. Effects of different media composition, light intensity and photoperiod on morphology and physiology of freshwater microalgae Ankistrodesmus falcatus–A potential strain for bio-fuel production. Bioresour. Technol. 2014, 171, 367–374. [Google Scholar] [CrossRef]
- Ji, F.; Hao, R.; Liu, Y.; Li, G.; Zhou, Y.; Dong, R. Isolation of a novel microalgae strain Desmodesmus sp. and optimization of environmental factors for its biomass production. Bioresour. Technol. 2013, 148, 249–254. [Google Scholar] [CrossRef]
- Wiltbank, L.B.; Kehoe, D.M. Two cyanobacterial photoreceptors regulate photosynthetic light harvesting by sensing teal, green, yellow, and red light. MBio 2016, 7, e02130-15. [Google Scholar] [CrossRef]
- Ramlov, F.; Souza, J.; Faria, A.V.; Maraschin, M.; Horta, P.A.; Yokoya, N.S. Growth and accumulation of carotenoids and nitrogen compounds in Gracilaria domingensis (Kütz.) Sonder ex Dickie (Gracilariales, Rhodophyta) cultured under different irradiance and nutrient levels. Rev. Bras. Farmacogn. 2011, 21, 255–261. [Google Scholar] [CrossRef]
- Beuckels, A.; Smolders, E.; Muylaert, K. Nitrogen availability influences phosphorus removal in microalgae-based wastewater treatment. Water Res. 2015, 77, 98–106. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Light Intensity (µmol m−2 s−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 | 65 | 100 | ||||||||
| NaNO3 (g L−1) | NaNO3 (g L−1) | NaNO3 (g L−1) | ||||||||
| 0.075 | 0.225 | 0.45 | 0.075 | 0.225 | 0.45 | 0.075 | 0.225 | 0.45 | ||
| Gompertz model constants | a | 0.74 | 0.88 | 2.05 | 1.33 | 1.74 | 1.04 | 1.59 | 1.03 | 2.27 |
| b | 9.3 | 14.26 | 1.51 | 1.76 | 1.16 | 3.11 | 1.42 | 1.28 | 1.27 | |
| c | 0.07 | 0.09 | 0.007 | 0.013 | 0.006 | 0.017 | 0.009 | 0.005 | 0.008 | |
| µmax (d−1) | 0.05 | 0.08 | 0.015 | 0.018 | 0.011 | 0.018 | 0.015 | 0.005 | 0.018 | |
| λ (h) | 122.3 | 139.1 | 70.64 | 56.1 | 25.72 | 124.7 | 45.13 | 56 | 34.31 | |
| G (h) | 13.74 | 8.22 | 46.96 | 38.23 | 62.27 | 39.37 | 46.64 | 45.78 | 39.2 | |
| R2 (%) | 93.83 | 94.71 | 95.42 | 88.6 | 91.49 | 86.2 | 94.82 | 91.47 | 95.59 | |
| Productivity (mg L−1 d−1) | 173.2 | 151.8 | 138.6 | 349.8 | 208.2 | 160.6 | 386 | 258 | 259.9 | |
| DW (g L−1) | N (g L−1) | Nitrogen Source | Y (N/DW) | Time (d) | Light Intensity (μmol m−2 s−1) | Reference |
|---|---|---|---|---|---|---|
| 5.54 | 1.78 | KNO3 | 0.045 | 16 | 350 | [41] |
| 3.4 | 1 | NaNO3 | 0.049 | 10 | 100 | [23] |
| 9.12 | 0.97 | NaNO3 | 0.018 | 14 | 18.85 | [24] |
| 3.0–5.403 | 0.075–0.45 | NaNO3 | 0.0023–0.025 | 14 | 100 | This report |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosa-Hernández, J.E.; Rodas-Zuluaga, L.I.; Castillo-Zacarías, C.; Rostro-Alanís, M.; de la Cruz, R.; Carrillo-Nieves, D.; Salinas-Salazar, C.; Fuentes Grunewald, C.; Llewellyn, C.A.; Olguín, E.J.; et al. Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum. Mar. Drugs 2019, 17, 460. https://doi.org/10.3390/md17080460
Sosa-Hernández JE, Rodas-Zuluaga LI, Castillo-Zacarías C, Rostro-Alanís M, de la Cruz R, Carrillo-Nieves D, Salinas-Salazar C, Fuentes Grunewald C, Llewellyn CA, Olguín EJ, et al. Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum. Marine Drugs. 2019; 17(8):460. https://doi.org/10.3390/md17080460
Chicago/Turabian StyleSosa-Hernández, Juan Eduardo, Laura Isabel Rodas-Zuluaga, Carlos Castillo-Zacarías, Magdalena Rostro-Alanís, Reynaldo de la Cruz, Danay Carrillo-Nieves, Carmen Salinas-Salazar, Claudio Fuentes Grunewald, Carole A. Llewellyn, Eugenia J. Olguín, and et al. 2019. "Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum" Marine Drugs 17, no. 8: 460. https://doi.org/10.3390/md17080460
APA StyleSosa-Hernández, J. E., Rodas-Zuluaga, L. I., Castillo-Zacarías, C., Rostro-Alanís, M., de la Cruz, R., Carrillo-Nieves, D., Salinas-Salazar, C., Fuentes Grunewald, C., Llewellyn, C. A., Olguín, E. J., Lovitt, R. W., Iqbal, H. M. N., & Parra-Saldívar, R. (2019). Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum. Marine Drugs, 17(8), 460. https://doi.org/10.3390/md17080460