A Novel High-Throughput Screening Platform Identifies Itaconate Derivatives from Marine Penicillium antarcticum as Inhibitors of Mesenchymal Stem Cell Differentiation

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
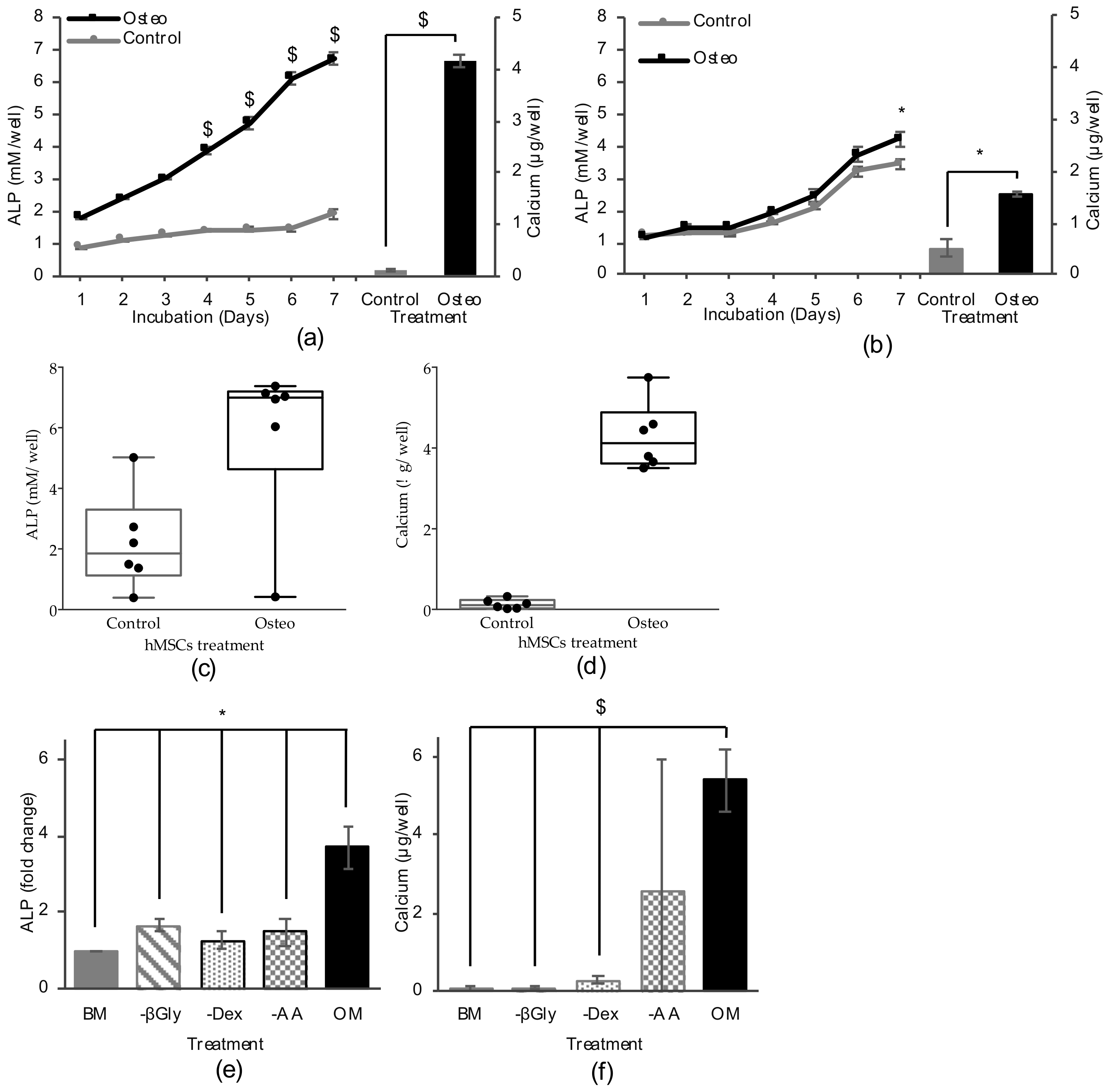
2.1. Osteogenic Differentiation Assay
2.1.1. Differentiation Method Development
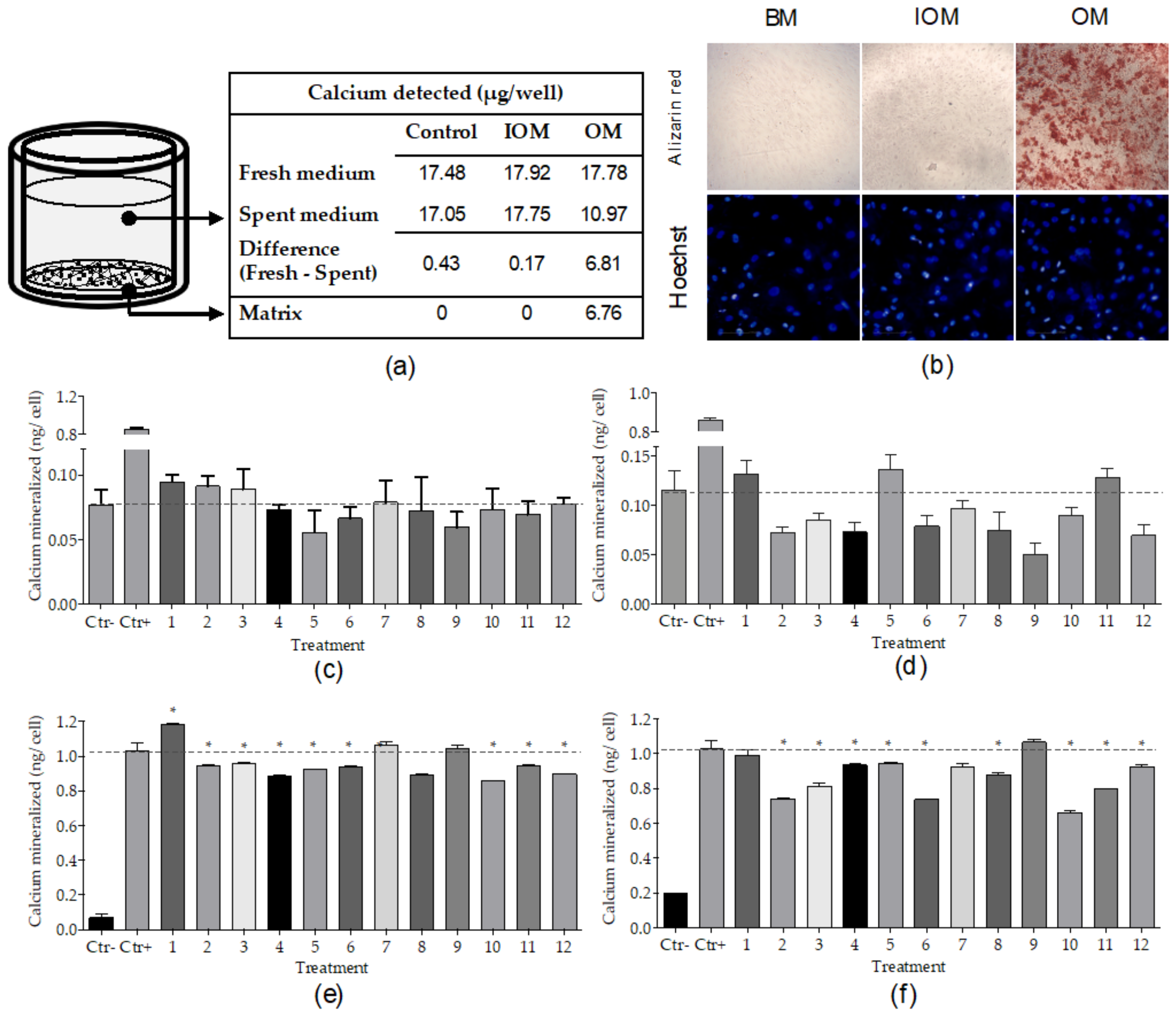
2.1.2. Screening Conditions: Differentiation Medium and Marker Detection
2.1.3. High Throughput Osteogenic Assay
2.2. Chondrogenic Assay
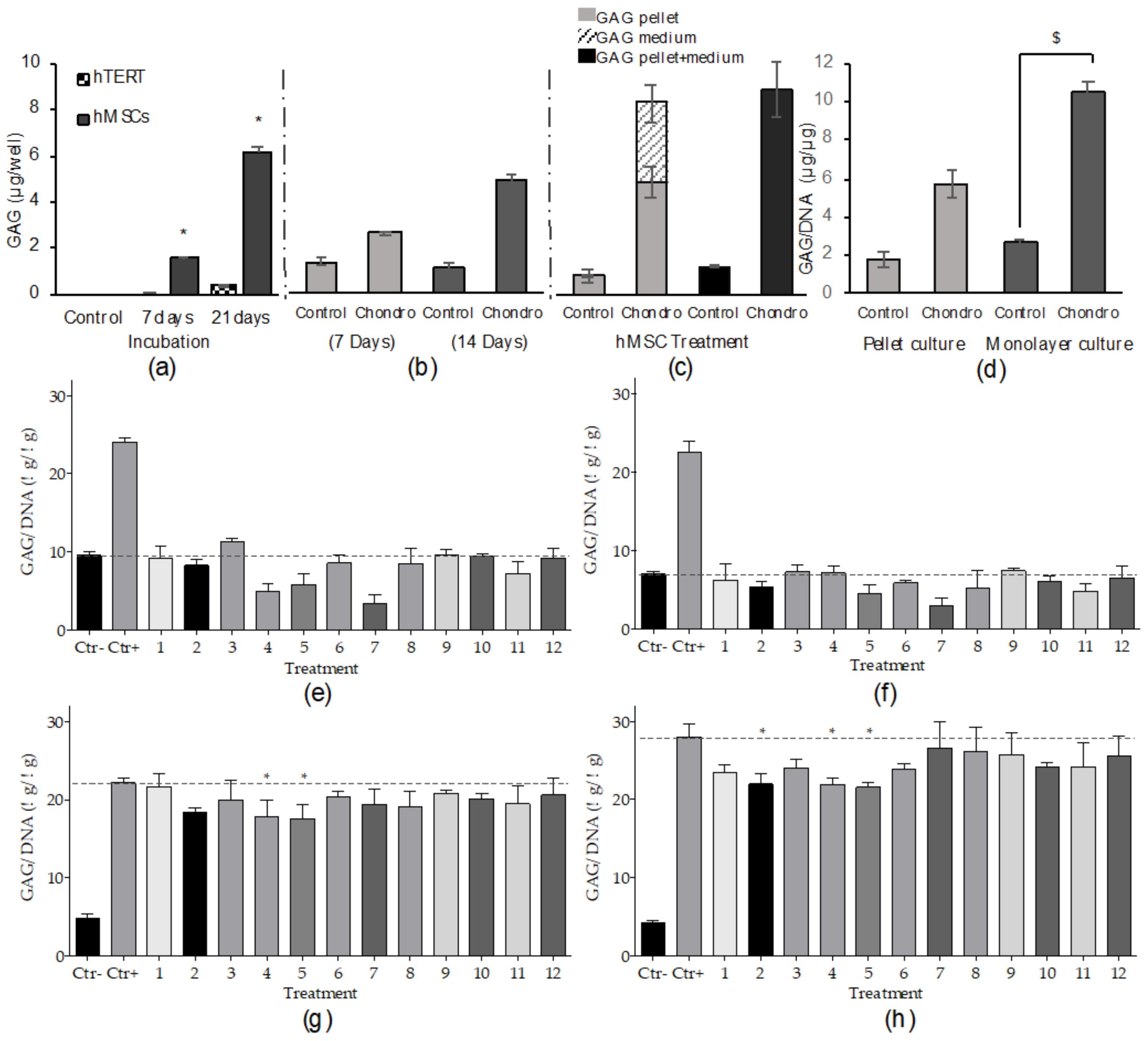
2.2.1. Differentiation Method Development
2.2.2. High-Throughput Chondrogenic Assay
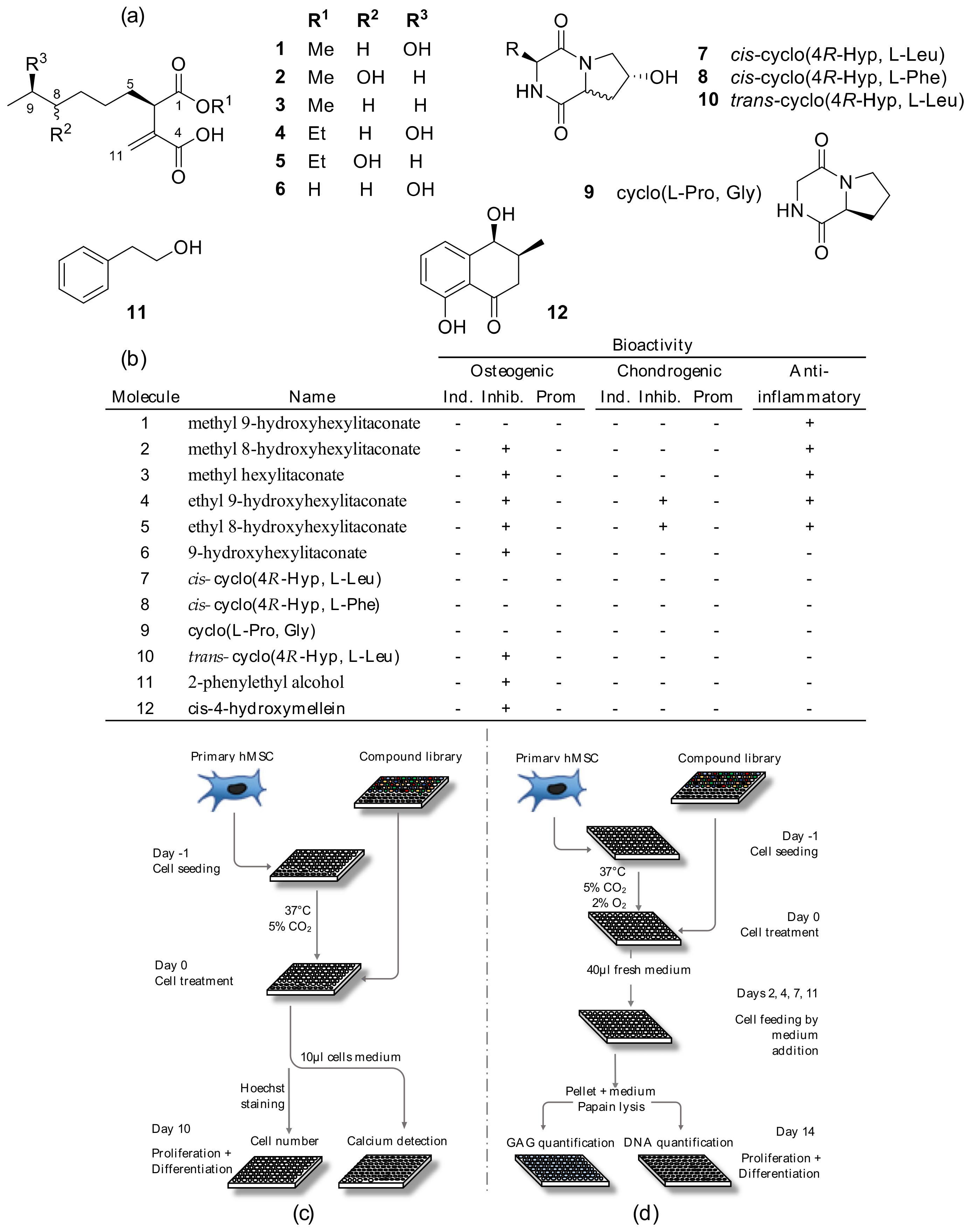
2.3. Fungal Metabolites Isolation and Structure Elucidation
2.4. Fungal Metabolites Bioactivity Screenings
3. Discussion
4. Materials and Methods
4.1. Primary and hTERT MSC Culture and Differentiation
4.2. Osteogenic Marker Detection for High Throughput Assay
4.3. Chondrogenic Marker Detection for High Throughput Assay
4.4. Automated High Throughput Osteogenic Assay
4.5. Automated High Throughput Chondrogenic Assay
4.6. Fungal Metabolites Isolation and Characterization
4.6.1. Biological Material
4.6.2. General Procedures
4.6.3. Isolation and Structure Elucidation
4.7. Fungal Metabolites Bioactivity Screenings
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leszczynska, A.; O’Doherty, A.; Farrell, E.; Pindjakova, J.; O’Brien, F.J.; O’Brien, T.; Barry, F.; Murphy, M. Differentiation of Vascular Stem Cells Contributes to Ectopic Calcification of Atherosclerotic Plaque. Stem Cells 2016, 34, 913–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckwalter, J.A.; Martin, J.A. Osteoarthritis. Adv. Drug Deliv. Rev. 2006, 58, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.S.; Le Merrer, M.; Glaser, D.L.; Pignolo, R.J.; Goldsby, R.E.; Kitterman, J.A.; Groppe, J.; Shore, E.M. Fibrodysplasia Ossificans Progressiva - FOP. Best Pract. Res. Clin. Rheumatol. 2008, 22, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.J.; Chung, H.W.; Choi, Y.L.; Moon, Y.W.; Seo, J.G.; Park, Y.S. Operative treatment of primary synovial osteochondromatosis of the hip. J. Bone Jt. Surg. 2006, 88, 2456–2464. [Google Scholar] [CrossRef]
- Go, A.S.; Chertow, G.M.; Fan, D.; McCulloch, C.E.; Hsu, C. Chronic Kidney Disease and the Risks of Death, Cardiovascular Events, and Hospitalization. N. Engl. J. Med. 2004, 351, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Besio, R.; Forlino, A. New frontiers for dominant osteogenesis imperfecta treatment: Gene/cellular therapy approaches. Adv. Regen. Biol. 2015, 2, 27964. [Google Scholar] [CrossRef]
- Kanis, J.A.; Melton, L.J.; Christiansen, C.; Johnston, C.C.; Khaltaev, N. The Diagnosis of Osteoporosis. J. Bone Miner. Res. 1994, 9, 1137–1141. [Google Scholar] [CrossRef]
- Besio, R.; Forlino, A. Treatment options for osteogenesis imperfecta. Expert Opin. Orphan Drugs 2015, 3165–3181. [Google Scholar] [CrossRef]
- Brewer, L.; Williams, D.; Moore, A. Current and future treatment options in osteoporosis. Eur. J. Clin. Pharmacol. 2011, 67, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.S.; Long, M.W.; Hankenson, K.D. Osteogenic Differentiation of Human Mesenchymal Stem Cells Is Regulated by Bone Morphogenetic Protein-6. J. Cell Biochem. 2006, 93, 538–554. [Google Scholar] [CrossRef]
- Govender, S.; Csimma, C.; Genant, H.K.; Valentin-Opran, A. Recombinant Human Bone Morphogenetic Protein-2 for Treatment of Open Tibial Fractures. J. Bone Jt. Surg. 2002, 84, 2123–2134. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Haynesworth, S.E.; Caplan, A.I.; Bruder, S.P. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J. Cell Biochem. 1997, 64, 295–312. [Google Scholar] [CrossRef]
- Choi, K.; Seo, Y.; Yoon, H.; Song, K.; Kwon, S.; Lee, H.; Park, J. Effect of Ascorbic Acid on Bone Marrow-Derived Mesenchymal Stem Cell Proliferation and Differentiation. J. Biosci. Bioeng. 2008, 105, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Fiorentini, E.; Granchi, D.; Leonardi, E.; Baldini, N.; Ciapetti, G. Effects of osteogenic differentiation inducers on in vitro expanded adult mesenchymal stromal cells. Int. J. Artif. Organs 2011, 34, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Song, I.; Caplan, A.I.; Dennis, J.E. In Vitro Dexamethasone Pretreatment Enhances Bone Formation of Human Mesenchymal Stem Cells In Vivo. J. Orthop. Res. 2009, 27, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Blaich, G.; Janssen, B.; Roth, G.; Salfeld, J. Overview: differentiating issues in the development of macromolecules compared with small molecules. In Handbook of Pharmaceutical Biotechnology; Cox Gad, S., Ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2006; pp. 89–123. [Google Scholar]
- Pyeritz, R.E. A Small Molecule for a Large Disease. N. Engl. J. Med. 2008, 358, 2829–2831. [Google Scholar] [CrossRef]
- Egusa, H.; Saeki, M.; Doi, M.; Fukuyasu, S.; Matsumoto, T.; Kamisaki, Y.; Yatani, H. A Small molecule Approach to Bone Regenerative Medicine in Dentistry. J. Oral. Biosci. 2010, 52, 107–118. [Google Scholar] [CrossRef]
- Lyssiotis, C.A.; Lairson, L.L.; Boitano, A.E.; Wurdak, H.; Zhu, S.; Schultz, P.G. Chemical control of stem cell fate and developmental potential. Angew Chemie Int. 2011, 50, 200–242. [Google Scholar] [CrossRef]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed Marine Natural Products in the Pharmaceutical and Cosmeceutical Industries: Tips for Success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Cho, S.G.; Kim, J.H.; Doan, T.K.P.; Hu, Q.S.; Ulhaq, R.; Song, E.K.; Yoon, T.R. Mevinolin enhances osteogenic genes (ALP, type I collagen and osteocalcin), CD44, CD47 and CD51 expression during osteogenic differentiation. Life Sci. 2009, 84, 290–295. [Google Scholar] [CrossRef]
- Carson, M.A.; Clarke, S.A. Bioactive Compounds from Marine Organisms: Potential for Bone Growth and Healing. Mar. Drugs 2018, 16, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, P.; Young, R.; O’Connell, E.; Baker, B.J.; Johnson, M.; Fearnhead, H.; Louise, A.; Murphy, M. Preliminary Evidence of Bioactivity from Deep Sea Coral and Sponge Extracts: Influencing Human Stem Cell Growth and Differentiation. In Proceedings of the XVI International Symposium on Marine Natural Products, XI European Conference on Marine Natural Products, Peniche, Portugal, 1–5 September 2019; Pedrosa, R.P., Gaudêncio, S., Vasconcelos, V., Eds.; Marine Drugs: Basel, Switzerland, 2020; Volume 18. [Google Scholar]
- Alves, H.; Dechering, K.; Van Blitterswijk, C.; De Boer, J. High-throughput assay for the identification of compounds regulating osteogenic differentiation of human mesenchymal stromal cells. PLoS ONE 2011, 6, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Brey, D.M.; Motlekar, N.A.; Diamond, S.L.; Mauck, R.L.; Garino, J.P.; Burdick, J.A. High-Throughput Screening of a Small Molecule Library for Promoters and Inhibitors of Mesenchymal Stem Cell Osteogenic Differentiation. Biotechnol. Bioeng. 2011, 108, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.M.; Heinegård, D.; McIntosh, A.; Sterchi, D.; Barry, F.P. Distribution of cartilage molecules in the developing mouse joint. Matrix Biol. 1999, 18, 487–497. [Google Scholar] [CrossRef]
- McRae, C.F.; Hocking, A.D.; Seppelt, R.D. Penicillium species from terrestrial habitats in the Windmill Islands, East Antarctica, including a new species, Penicillium antarcticum. Polar Biol. 1999, 21, 97–111. [Google Scholar] [CrossRef]
- Klemke, C.; Kehraus, S.; Wright, A.D.; Koenig, G.M. New secondary metabolites from the marine endophytic fungus Apiospora montagnei. J. Nat. Prod. 2004, 67, 1058–1063. [Google Scholar] [CrossRef]
- Nakahashi, A.; Miura, N.; Monde, K.; Tsukamoto, S. Stereochemical studies of hexylitaconic acid, an inhibitor of p53-HDM2 interaction. Bioorg. Med. Chem. Lett. 2009, 19, 3027–3303. [Google Scholar] [CrossRef] [Green Version]
- Antia, B.S.; Aree, T.; Kasettrathat, C.; Wiyakrutta, S.; Ekpa, O.D.; Ekpe, U.J.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Itaconic acid derivatives and diketopiperazine from the marine-derived fungus Aspergillus aculeatus CRI322-03. Phytochemistry 2011, 72, 816–820. [Google Scholar] [CrossRef]
- Furtado, N.A.J.C.; Pupo, M.T.; Carvalho, I.; Campo, V.L.; Duarte, M.C.T.; Bastos, J.K. Diketopiperazines produced by an Aspergillus fumigatus Brazilian strain. J. Brazil. Chem. Soc. 2005, 16, 1448–1453. [Google Scholar] [CrossRef]
- Shigemori, H.; Tenma, M.; Shimazaki, K.; Kobayashi, J. Three New Metabolites from the Marine Yeast Aureobasidium pullulans. J. Nat. Prod. 1998, 61, 696–698. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, J.; De Carvalho, V.R. Epi-isoshinanolone from Plumbago scandens. Phytochemistry 1986, 25, 764–765. [Google Scholar] [CrossRef]
- Huang, A.H.; Motlekar, N.A.; Stein, A.; Diamond, S.L.; Shore, E.M.; Mauck, R.L. High-throughput screening for modulators of mesenchymal stem cell chondrogenesis. Ann. Biomed. Eng. 2008, 36, 1909–1921. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.J.; Kim, J.; Kwon, S.K.; Oh, K.; Lee, J.; Lee, D.S.; Cho, J.; Park, S.B. A potent small-molecule inducer of chondrogenic differentiation of human bone marrow-derived mesenchymal stem cells. Chem. Sci. 2012, 3, 3071. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruder, S.P.; Jaiswal, N.; Haynesworth, S.E. Growth Kinetics, Self-Renewal, and the Osteogenic Potential of Purified Human Mesenchymal Stem Cells During Extensive Subcultivation and Following Cryopreservation. J. Cell Biochem. 1997, 64, 278–294. [Google Scholar]
- Johnstone, B.; Hering, T.M.; Caplan, A.I.; Goldberg, V.M.; Yoo, J.U. In Vitro Chondrogenesis of Bone Marrow-Derived Mesenchymal Progenitor Cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef]
- Bovio, E.; Garzoli, L.; Poli, A.; Luganini, A.; Villa, P.; Musumeci, R.; McCormack, G.P.; Cocuzza, C.E.; Gribaudo, G.; Mehiri, M.; et al. Marine Fungi from the Sponge Grantia compressa. Mar. Drugs 2019, 17, 220. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, S.K.; Prakash, V.; Ranjan, N. Marine fungi: A source of potential anticancer compounds. Front. Microbiol. 2018, 8, 1–24. [Google Scholar] [CrossRef]
- Nielsen, J.C.; Grijseels, S.; Prigent, S.; Ji, B.; Dainat, J.; Nielsen, K.F.; Frisvad, J.C.; Workman, M.; Nielsen, J. Global analysis of biosynthetic gene clusters reveals vast potential of secondary metabolite production in Penicillium species. Nat. Microbiol. 2017, 2, 17044. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Santos, L.; Silva, B.M.V.; Abreu, A.C.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Biodiversity of Penicillium species from marine environments in Portugal and description of Penicillium lusitanum sp. nov., a novel species isolated from sea water. Int. J. Syst. Evol. Microbiol. 2019, 39, 3014–3021. [Google Scholar]
- Oleinikova, G.K.; Kirichuk, N.N.; Afiyatullov, S.S. Nonpolar Compounds and Free Fatty Acids from Several Isolates of Marine Fungus Penicillium antarcticum. Chem. Nat. Compd. 2018, 54, 535–537. [Google Scholar] [CrossRef]
- Geiger, M.; Guitton, Y.; Vansteelandt, M.; Kerzaon, I.; Blanchet, E.; Robiou du Pont, T.; Frisvad, J.C.; Hess, P.; Pouchus, Y.F.; Grovel, O. Cytotoxicity and mycotoxin production of shellfish-derived Penicillium spp., a risk for shellfish consumers. Lett. Appl. Microbiol. 2013, 57, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vansteelandt, M.; Kerzaon, I.; Blanchet, E.; Fossi Tankoua, A.; Robiou du Pont, T.; Joubert, Y.; Monteau, F.; Le Bizec, B.; Frisvad, J.C.; Pouchus, Y.F.; et al. Patulin and secondary metabolite production by marine-derived Penicillium strains. Fungal Biol. 2012, 116, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.M.; Asotra, K.; Fitzpatrick, L.A.; Qiao, J.; Wilkin, D.J.; Detrano, R.C.; Dunstan, C.R.; Shah, P.K.; Rajavashisth, T.B. Calcification in atherosclerosis: Bone biology and chronic inflammation at the arterial crossroads. PNAS 2003, 100, 11201–11206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Sata, M.; Natori, T.; Kim-Kaneyama, J.; Nose, K.; Shibanuma, M.; Hirata, Y.; Nagai, R. Circulating progenitor cells contribute to neointimal formation in nonirradiated chimeric mice. FASEB J. 2008, 22, 428–436. [Google Scholar] [CrossRef]
- Somoza, R.A.; Welter, J.F.; Correa, D.; Caplan, A.I. Chondrogenic Differentiation of Mesenchymal Stem Cells: Challenges and Unfulfilled Expectations. Tissue Eng. Part B Rev. 2014, 20, 596–608. [Google Scholar] [CrossRef] [Green Version]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chung, T.; Oldenburg, K. Simple Statistical Parameter for Use in Evaluation and Validation of High Throughput Screening Assays. J. Biomol. Screen 1999, 4, 67–73. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchese, P.; Mahajan, N.; O’Connell, E.; Fearnhead, H.; Tuohy, M.; Krawczyk, J.; Thomas, O.P.; Barry, F.; Murphy, M.J. A Novel High-Throughput Screening Platform Identifies Itaconate Derivatives from Marine Penicillium antarcticum as Inhibitors of Mesenchymal Stem Cell Differentiation. Mar. Drugs 2020, 18, 192. https://doi.org/10.3390/md18040192
Marchese P, Mahajan N, O’Connell E, Fearnhead H, Tuohy M, Krawczyk J, Thomas OP, Barry F, Murphy MJ. A Novel High-Throughput Screening Platform Identifies Itaconate Derivatives from Marine Penicillium antarcticum as Inhibitors of Mesenchymal Stem Cell Differentiation. Marine Drugs. 2020; 18(4):192. https://doi.org/10.3390/md18040192
Chicago/Turabian StyleMarchese, Pietro, Nipun Mahajan, Enda O’Connell, Howard Fearnhead, Maria Tuohy, Janusz Krawczyk, Olivier P. Thomas, Frank Barry, and Mary J. Murphy. 2020. "A Novel High-Throughput Screening Platform Identifies Itaconate Derivatives from Marine Penicillium antarcticum as Inhibitors of Mesenchymal Stem Cell Differentiation" Marine Drugs 18, no. 4: 192. https://doi.org/10.3390/md18040192
APA StyleMarchese, P., Mahajan, N., O’Connell, E., Fearnhead, H., Tuohy, M., Krawczyk, J., Thomas, O. P., Barry, F., & Murphy, M. J. (2020). A Novel High-Throughput Screening Platform Identifies Itaconate Derivatives from Marine Penicillium antarcticum as Inhibitors of Mesenchymal Stem Cell Differentiation. Marine Drugs, 18(4), 192. https://doi.org/10.3390/md18040192