Cytotoxic Activity of Aplykurodin A Isolated From Aplysia kurodai against AXIN1-Mutated Hepatocellular Carcinoma Cells by Promoting Oncogenic β-Catenin Degradation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
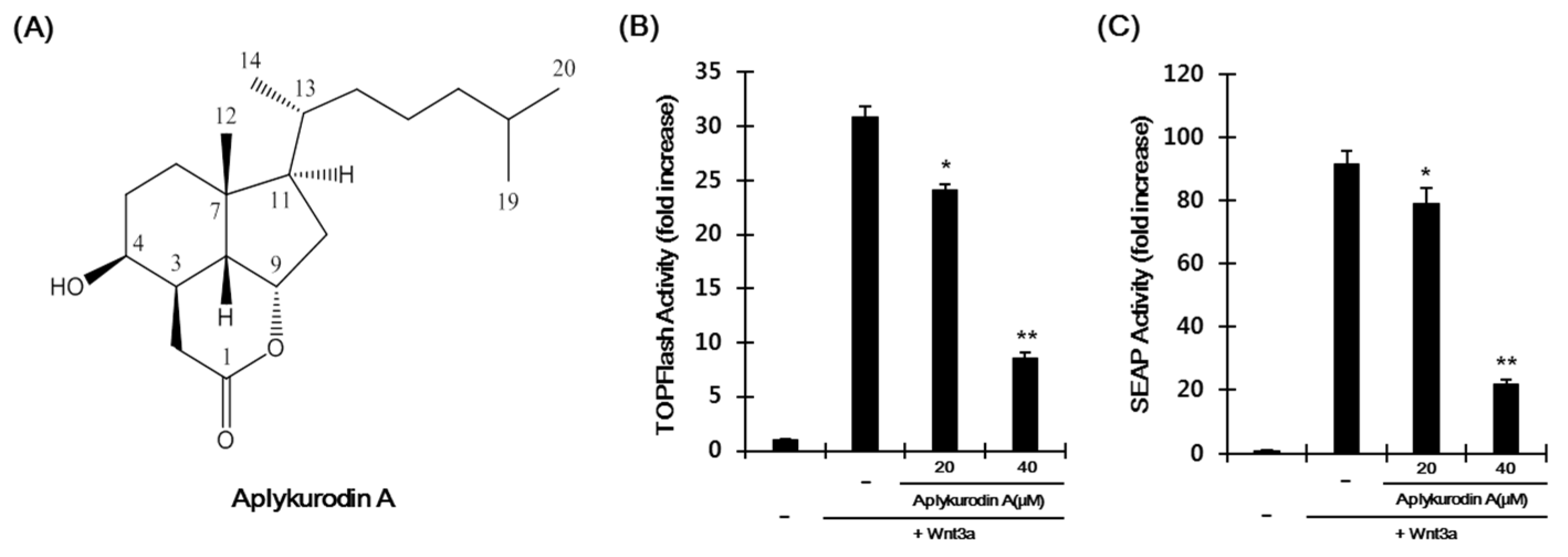
2.1. Aplykurodin A Suppresses the Wnt/β-Catenin Pathway
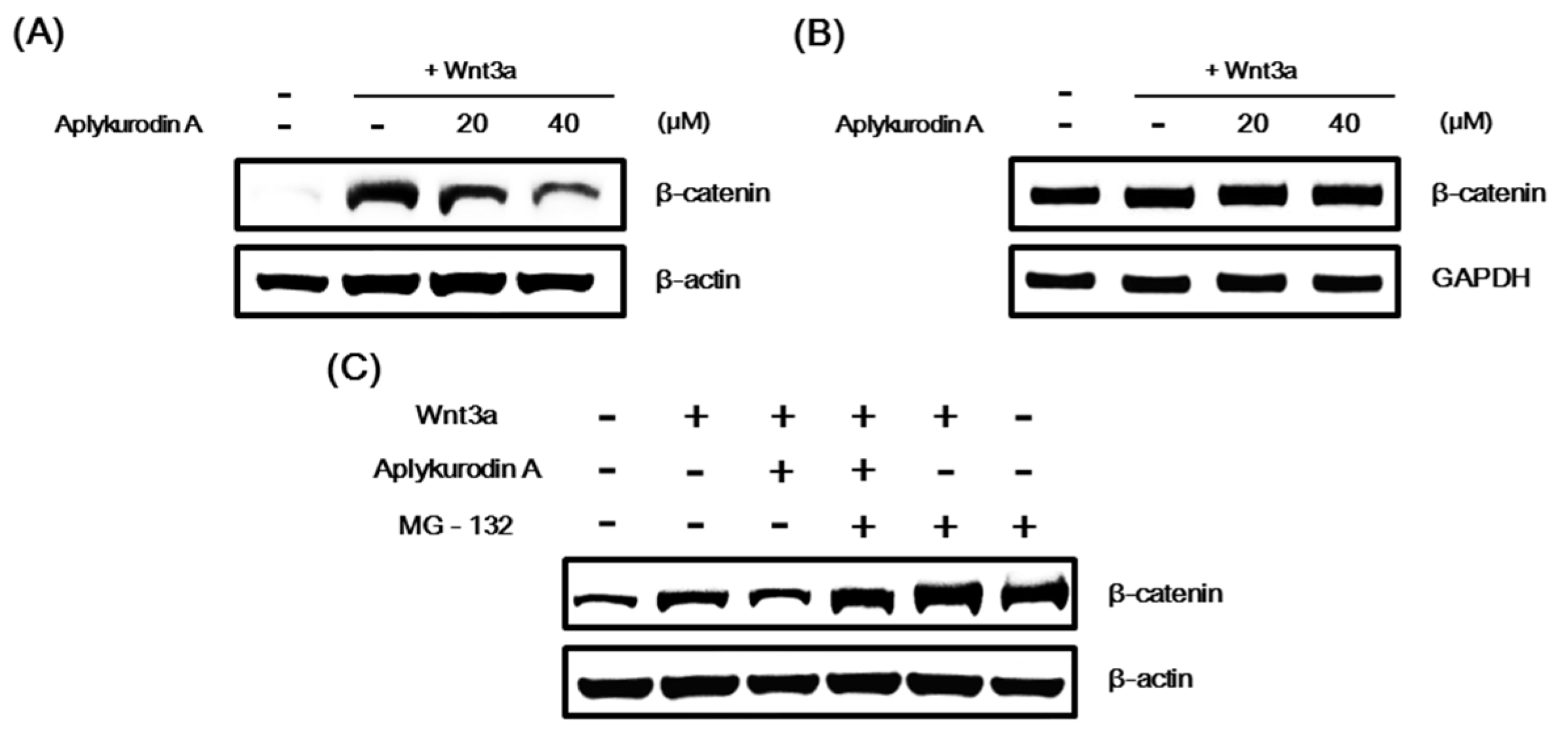
2.2. Aplykurodin A Promotes Proteasomal Degradation of β-Catenin
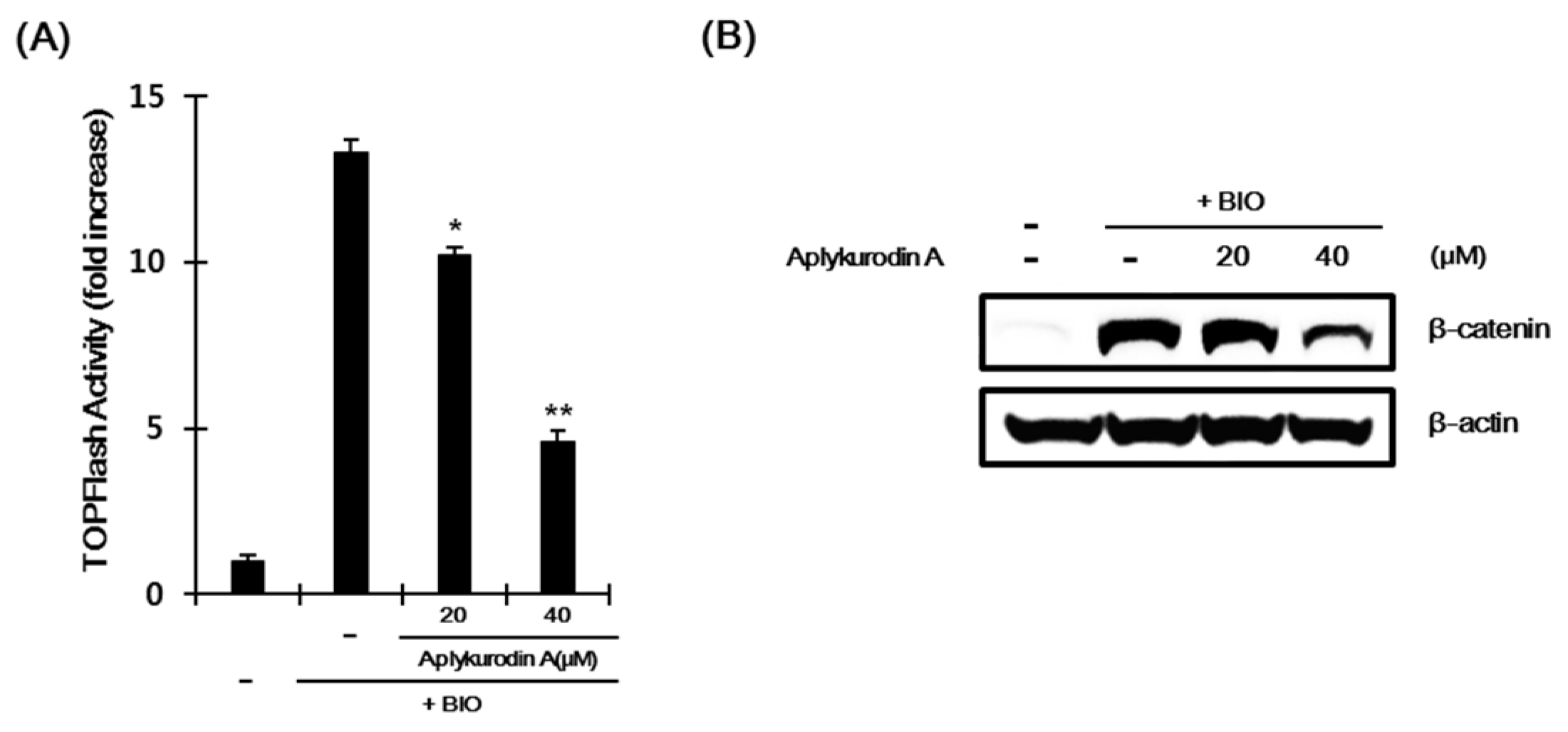
2.3. Aplykurodin A Promotes β-Catenin Degradation Through a Mechanism Independent of GSK-3β
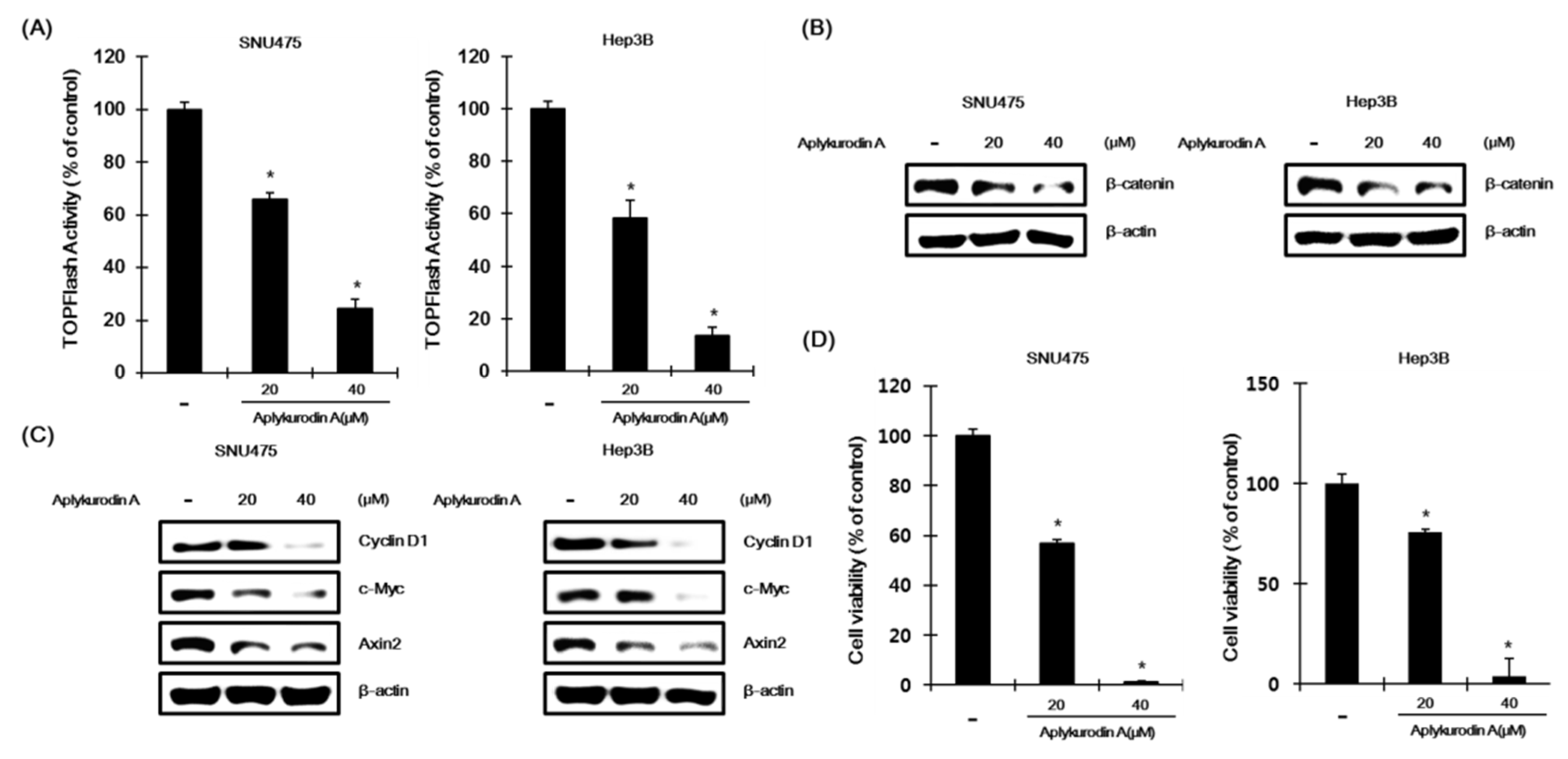
2.4. Aplykurodin A Has Anti-Porliferative Effects in AXIN1-Mutated HCC Cells
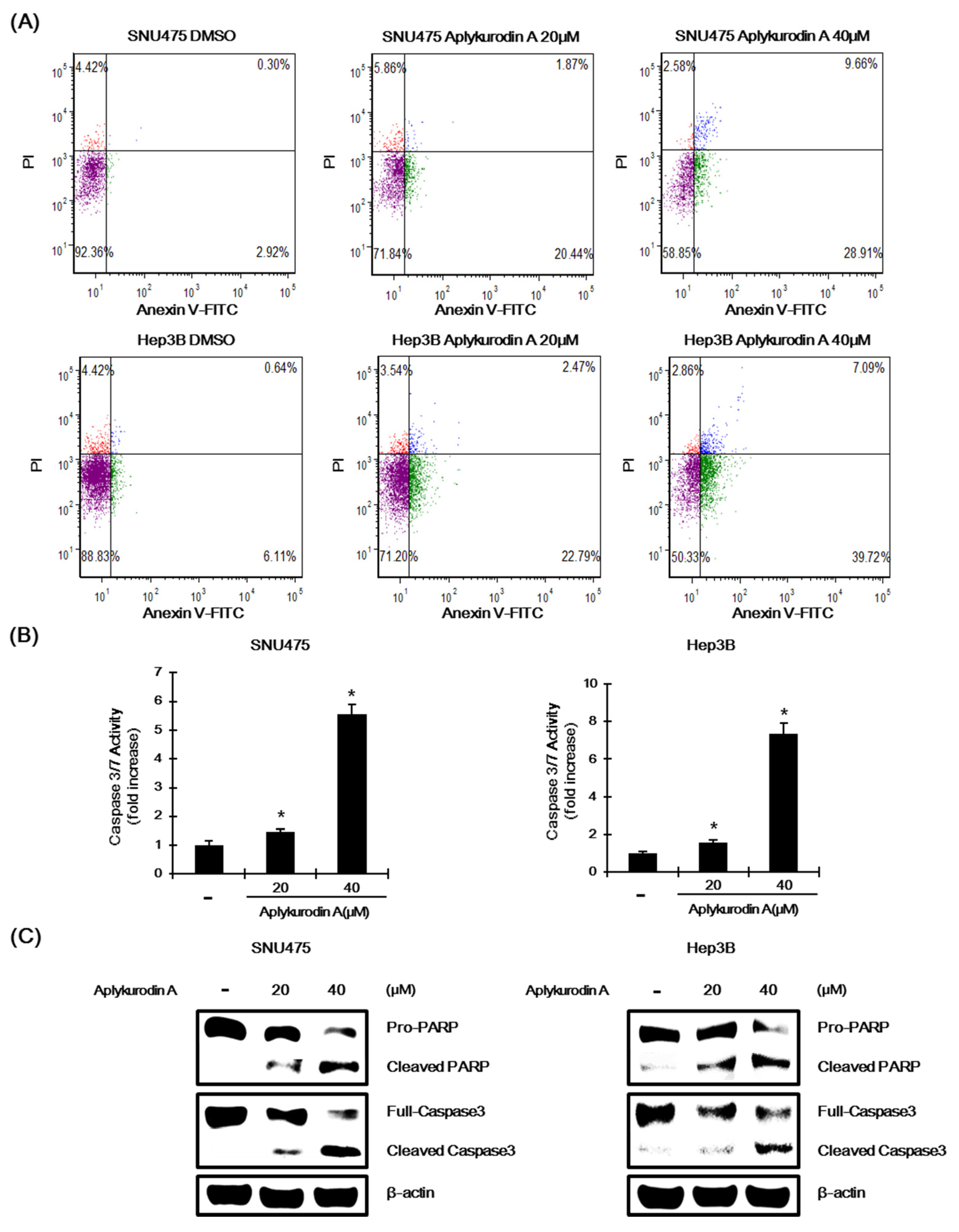
2.5. Aplykurodin A Induces Apoptosis in SNU475 and Hep3B Cells
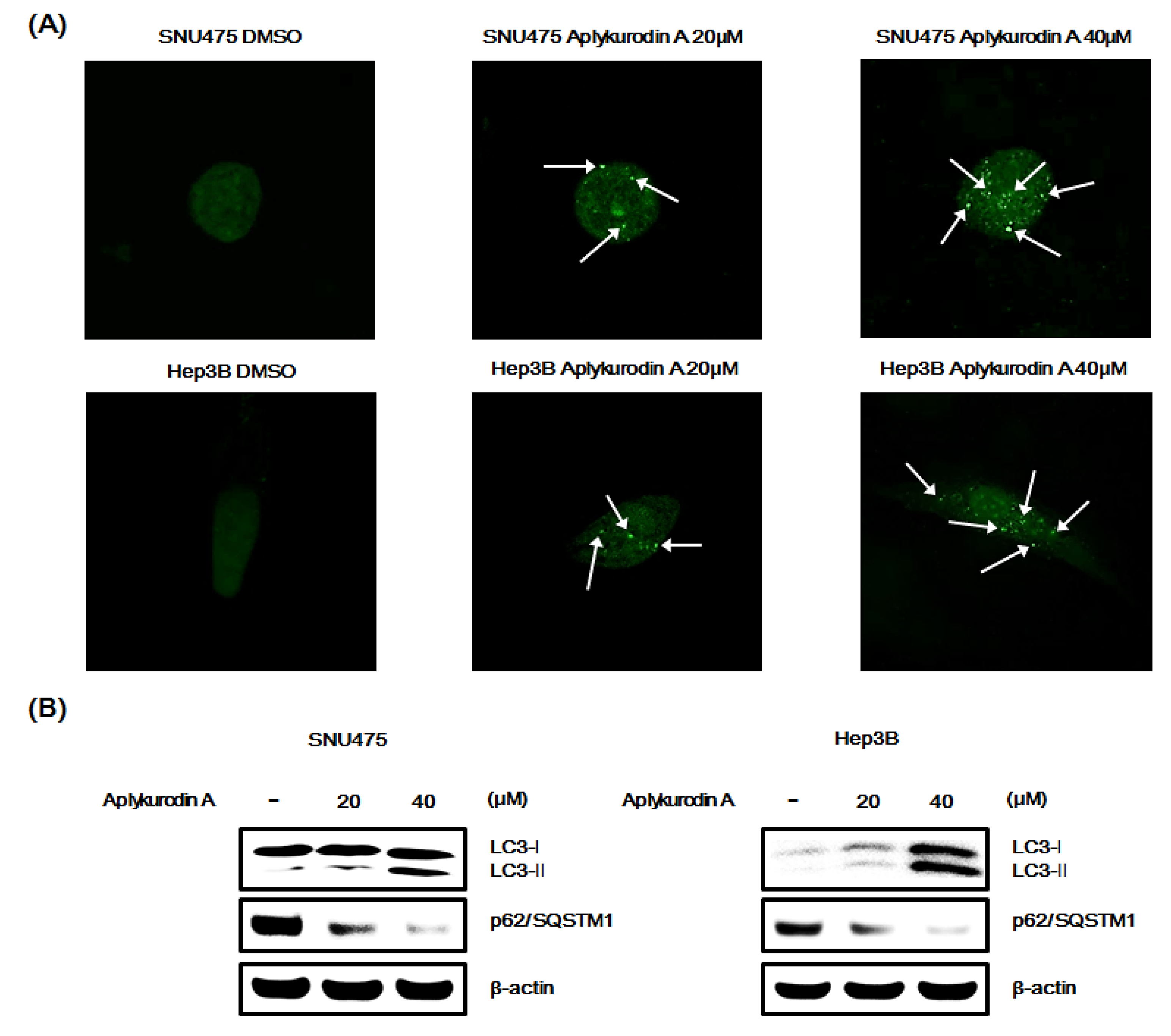
2.6. Aplykurodin A Induces Autophagy in SNU475 and Hep3B Cells
3. Discussion
4. Materials and Methods
4.1. Isolation of Aplykurodin A
4.2. Cell Culture, Reporter Assay, Transfection, and Chemicals Used
4.3. Western Blotting and Antibodies
4.4. RNA Extraction and Semi-Quantitative RT-PCR
4.5. Cell Viability Assay
4.6. Apoptosis Analysis and Caspase-3/7 Assay
4.7. Fluorescence Microscope Analysis of GFP-LC3
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA A Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhl, P.; Fricker, G.; Haberkorn, U.; Mier, W. Current Status in the Therapy of Liver Diseases. Int. J. Mol. Sci. 2014, 15, 7500–7512. [Google Scholar] [CrossRef] [Green Version]
- Padhya, K.; Marrero, J.; Singal, A. Recent advances in the treatment of hepatocellular carcinoma. Curr. Opin. Gastroenterol. 2013, 29, 285–292. [Google Scholar] [CrossRef]
- Llovet, J.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.; de Oliveira, A.; Santoro, A.; Raoul, J.; Forner, A.; et al. Sorafenib in Advanced Hepatocellular Carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Wodarz, A.; Nusse, R. Mechanisms of Wnt signaling in development. Annu. Rev. Cell Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef] [Green Version]
- Huelsken, J.; Birchmeier, W. New aspects of Wnt signaling pathways in higher vertebrates. Curr. Opin. Genet. Dev 2001, 11, 547–553. [Google Scholar] [CrossRef]
- Miller, J.R. The Wnts. Genome Biol. 2002, 3. reviews 3001.1–15. [Google Scholar]
- Hart, M.J.; de los Santos, R.; Albert, I.N.; Rubinfeld, B.; Polakis, P. Down-regulation of β-catenin by human Axin and its association with the APC tumor suppressor, β-catenin and GSK3β. Curr. Biol. 1998, 8, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, S.; Kishida, S.; Yamamoto, H.; Murai, H.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the Wnt signaling pathway, forms a complex with GSK-3beta and beta-catenin and promotes GSK-3beta-dependent phosphorylation of beta-catenin. EMBO J. 1998, 17, 1371–1384. [Google Scholar] [CrossRef]
- Aberle, H.; Bauer, A.; Stapper, T.J.; Kispert, A.; Kemler, R. β-Catenin is a target for the ubiquitin-proteasome pathway. EMBO J. 1997, 16, 3797–3804. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.; Concordet, J.P.; Lassot, I.; Albert, I.; del los Santos, R.; Durand, H.; Perret, C.; Rubinfeld, B.; Margottin, F.; Benarous, R.; et al. The F-box protein β-TrCP associates with phosphorylated β-catenin and regulates its activity in the cell. Curr. Biol. 1999, 9, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Lee, J.; Shon, J.; Phuc, N.M.; Jee, J.G.; Liu, K. The inhibitory potential of Broussochalcone A for the human cytochrome P450 2J2 isoform and its anti-cancer effects via FOXO3 activation. Phytomedicine 2018, 42, 199–206. [Google Scholar] [CrossRef]
- Srisuttee, R.; Koh, S.; Malilas, W.; Moon, J.; Cho, I.; Jhun, B.; Horio, Y.; Chung, Y. SIRT1 sensitizes hepatocellular carcinoma cells expressing hepatitis B virus X protein to oxidative stress-induced apoptosis. Biochem. Biophys. Res. Commun. 2012, 429, 45–50. [Google Scholar] [CrossRef]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- He, T.C.; Chan, T.A.; Vogelstein, B.; Kinzler, K.W. PPAR-d is an APC-regulated target of nonsteroidal anti-inflammatory drugs. Cell 1999, 99, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Tetsu, O.; McCormick, F. β-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Tsunoda, T.; Seiki, M.; Nakamura, Y.; Furukawa, Y. Identification of membrane-type matrix metalloproteinase-1 as a target of the β-catenin/Tcf4 complex in human colorectal cancers. Oncogene 2002, 21, 5861–5867. [Google Scholar] [CrossRef] [Green Version]
- Spinella, A.; Gavaqnin, M.; Crispino, A.; Cimino, G.; Martinez, E.; Ortea, J.; Sodano, G. 4-acetylaplykurodin B and aplydurodinone B, two ichthyotoxic degraded sterols from the Mediterranean mollusk Aplysia fasciata. J. Nat. Prod. 1992, 55, 989–993. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, H.; Su, A.; He, J.; Yu, H. Effect of aplysin on the proliferation and apoptosis in human gastric cancer cell SGC-7901. Chin. Pharmacol. Bull 2010, 26, 333–337. [Google Scholar]
- Yamada, K.; Ojika, M.; Ishigaki, T.; Yoshida, Y. Aplyronin A, a potent antitumor substance, and the congeners aplyronines B and C isolated from the sea hare Aplysia kurodai. J. Am. Chem. Soc. 1993, 115, 11020–11021. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Yamashita, Y.; Ohta, T. New cytotoxic and antibacterial compounds isolated from the sea hare, Aplysia kurodai. Mar. Drugs 2005, 3, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Higuchi, R.; Komori, T.; Fujioka, T.; Mihashi, K. Isolation and structures of aplykurodins A and B, two new isoprenoids from the marine mollusk Aplysia kurodai. Tetrahedron Lett. 1986, 27, 1153–1156. [Google Scholar] [CrossRef]
- Park, S.; Gwak, J.; Cho, M.; Song, T.; Won, J.; Kim, D.E.; Shin, J.G.; Oh, S. Hexachlorophene inhibits Wnt/β-catenin pathway by promoting Siah-mediated-β-catenin degradation. Mol. Pharmacol. 2006, 70, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.J.; Cho, M.; Song, J.Y.; Yun, Y.S.; Choi, I.W.; Kim, D.E.; Park, B.S.; Oh, S. Natural derivatives of curcumin attenuate the Wnt/β-catenin pathway through down-regulation of the transcriptional coactivator p300. Biochem. Biophys. Res. Commun. 2008, 377, 1304–1308. [Google Scholar] [CrossRef]
- Latres, E.; Chiaur, D.S.; Pagano, M. The human F box protein beta-Trcp associates with the Cul1/Skp1 complex and regulates the stability of β-catenin. Oncogene 1999, 18, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Meijer, L.; Skaltsounis, A.L.; Magiatis, P.; Polychronopoulos, P.; Knockaert, M.; Leost, M.; Ryan, X.P.; Vonica, C.A.; Brivanlou, A.; Dajani, R.; et al. GSK-3-selective inhibitors derived from Tyrian purple indirubins. Chem. Biol. 2003, 10, 1255–1266. [Google Scholar] [CrossRef] [Green Version]
- Ilyas, M.; Tomlinson, I.P.; Rowan, A.; Pignatelli, M.; Bodmer, W.F. β-Catenin mutations in cell lines established from human colorectal cancers. Proc. Natl. Acad. Sci. USA 1997, 94, 10330–10334. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Shi, C.; Jiang, L.; Tolstykh, T.; Cao, H.; Bangari, D.; Ryan, S.; Levit, M.; Jin, T.; Mamaat, K.; et al. Oncogenic dependency on β-catenin in liver cancer cell lines correlates with pathway activation. Oncotarget 2017, 8, 114526–114539. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Su, Y.; Lee, P.; Lei, H. Targeting NFKB by autophagy to polarize hepatoma-associated macrophage differentiation. Autophagy 2013, 9, 619–621. [Google Scholar] [CrossRef] [Green Version]
- Su, N.; Wang, P.; Li, Y. Role of Wnt/β-catenin pathway in inducing autophagy and apoptosis in multiple myeloma cells. Oncol. Lett. 2016, 12, 4623–4629. [Google Scholar] [CrossRef] [Green Version]
- Park, C.S.; Kim, S.I.; Lee, M.S.; Youn, C.Y.; Kim, D.J.; Jho, E.H.; Song, W.K. Modulation of beta-catenin phosphorylation/degradation by cyclin-dependent kinase 2. J. Biol. Chem. 2004, 279, 19592–19599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwak, J.; Cho, M.; Gong, S.; Won, J.; Kim, D.; Kim, E.; Lee, S.; Kim, M.; Kim, T.; Shin, J.; et al. Protein-kinase-C-mediated β-catenin phosphorylation negatively regulates the Wnt/β-catenin pathway. J. Cell Sci. 2006, 119, 4702–4709. [Google Scholar] [CrossRef] [Green Version]
- Gwak, J.; Jung, S.J.; Kang, D.I.; Kim, E.Y.; Kim, D.E.; Chung, Y.H.; Shin, J.G.; Oh, S. Stimulation of protein kinase C-alpha suppresses colon cancer cell proliferation by down-regulation of beta-catenin. J. Cell Mol. Med. 2009, 13, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, X. The signal pathways in azoxymethane-induced colon cancer and preventive implications. Cancer Biol. Ther. 2009, 8, 1313–1317. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase inhibition stabilizes axin and antagonizes Wnt signaling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.; Kim, J.; Song, G.; Zhou, W.; Goh, S.; Na, M.; Oh, S. Cytotoxic activity of aeroplysinin-1 against colon cancer cells by promoting β-catenin degradation. Food Chem. Toxicol. 2016, 93, 66–72. [Google Scholar] [CrossRef]
- Shin, S.; Son, Y.; Liu, K.; Kang, W.; Oh, S. Cytotoxic activity of broussochalcone a against colon and liver cancer cells by promoting destruction complex-independent β-catenin degradation. Food Chem. Toxicol. 2019, 131, 110550. [Google Scholar] [CrossRef]
- Norbury, C.; Zhivotovsky, B. DNA damage-induced apoptosis. Oncogene 2004, 23, 2797–2808. [Google Scholar] [CrossRef] [Green Version]
- Livesey, K.; Kang, R.; Vernon, P.; Buchser, W.; Loughran, P.; Watkins, S.; Zhang, L.; Manfredi, J.; Zeh, H.; Li, L.; et al. p53/HMGB1 Complexes Regulate Autophagy and Apoptosis. Cancer Res. 2012, 72, 1996–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gump, J.; Thorburn, A. Autophagy and apoptosis: What is the connection? Trends Cell Biol. 2011, 21, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Dignam, J.D.; Lebovitz, R.M.; Roeder, R.G. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 1983, 11, 1475–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Zhou, W.; Na, M.; Oh, S. Cytotoxic Activity of Aplykurodin A Isolated From Aplysia kurodai against AXIN1-Mutated Hepatocellular Carcinoma Cells by Promoting Oncogenic β-Catenin Degradation. Mar. Drugs 2020, 18, 210. https://doi.org/10.3390/md18040210
Lee J, Zhou W, Na M, Oh S. Cytotoxic Activity of Aplykurodin A Isolated From Aplysia kurodai against AXIN1-Mutated Hepatocellular Carcinoma Cells by Promoting Oncogenic β-Catenin Degradation. Marine Drugs. 2020; 18(4):210. https://doi.org/10.3390/md18040210
Chicago/Turabian StyleLee, Jaehoo, Wei Zhou, MinKyun Na, and Sangtaek Oh. 2020. "Cytotoxic Activity of Aplykurodin A Isolated From Aplysia kurodai against AXIN1-Mutated Hepatocellular Carcinoma Cells by Promoting Oncogenic β-Catenin Degradation" Marine Drugs 18, no. 4: 210. https://doi.org/10.3390/md18040210
APA StyleLee, J., Zhou, W., Na, M., & Oh, S. (2020). Cytotoxic Activity of Aplykurodin A Isolated From Aplysia kurodai against AXIN1-Mutated Hepatocellular Carcinoma Cells by Promoting Oncogenic β-Catenin Degradation. Marine Drugs, 18(4), 210. https://doi.org/10.3390/md18040210